
Chemical Characterization, α-Glucosidase, α-Amylase and Lipase Inhibitory Properties of the Australian Honey Bee Propolis



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Sample Collection
2.3. Extraction
2.4. Preparative-Scale Isolation and Purification
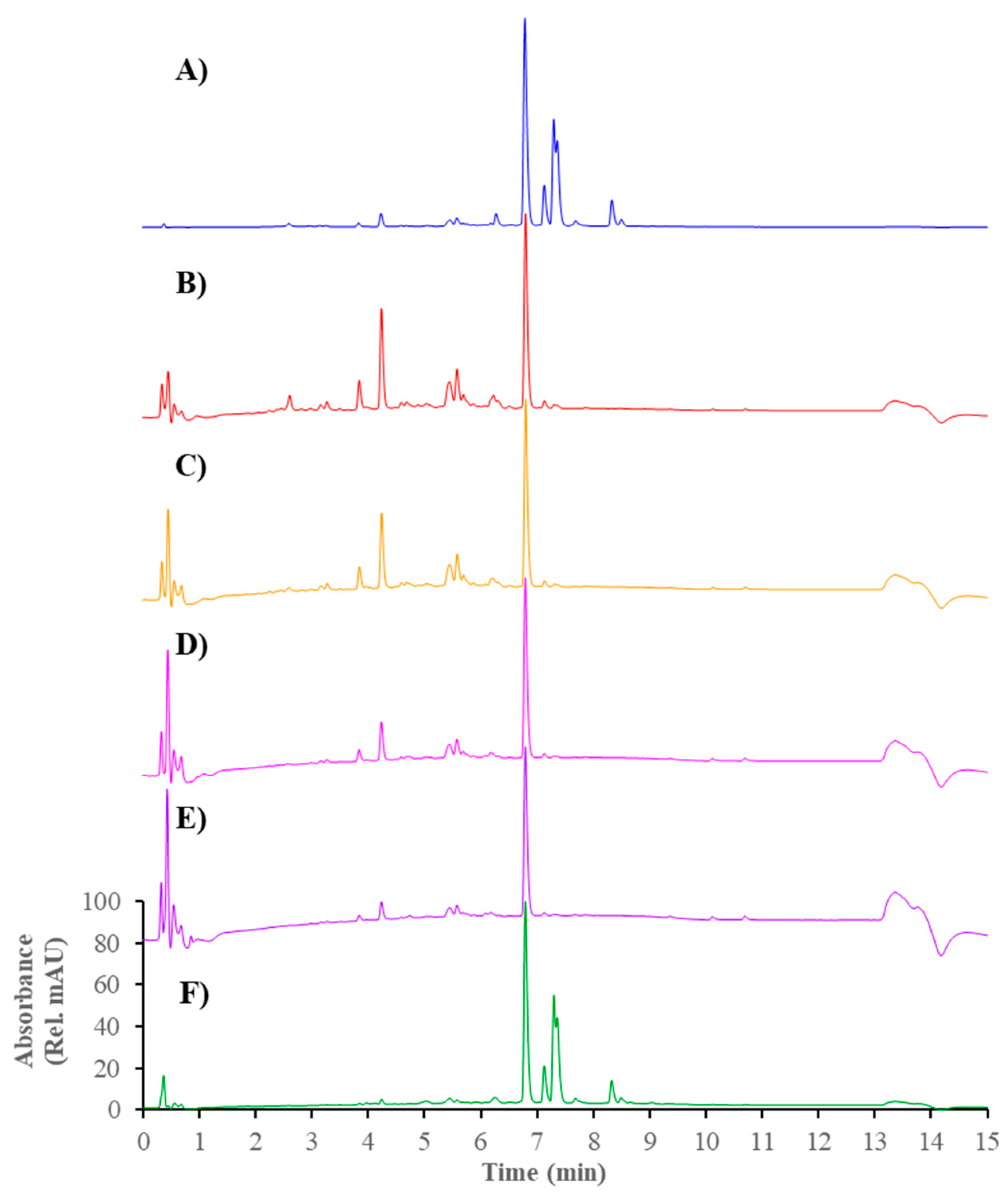
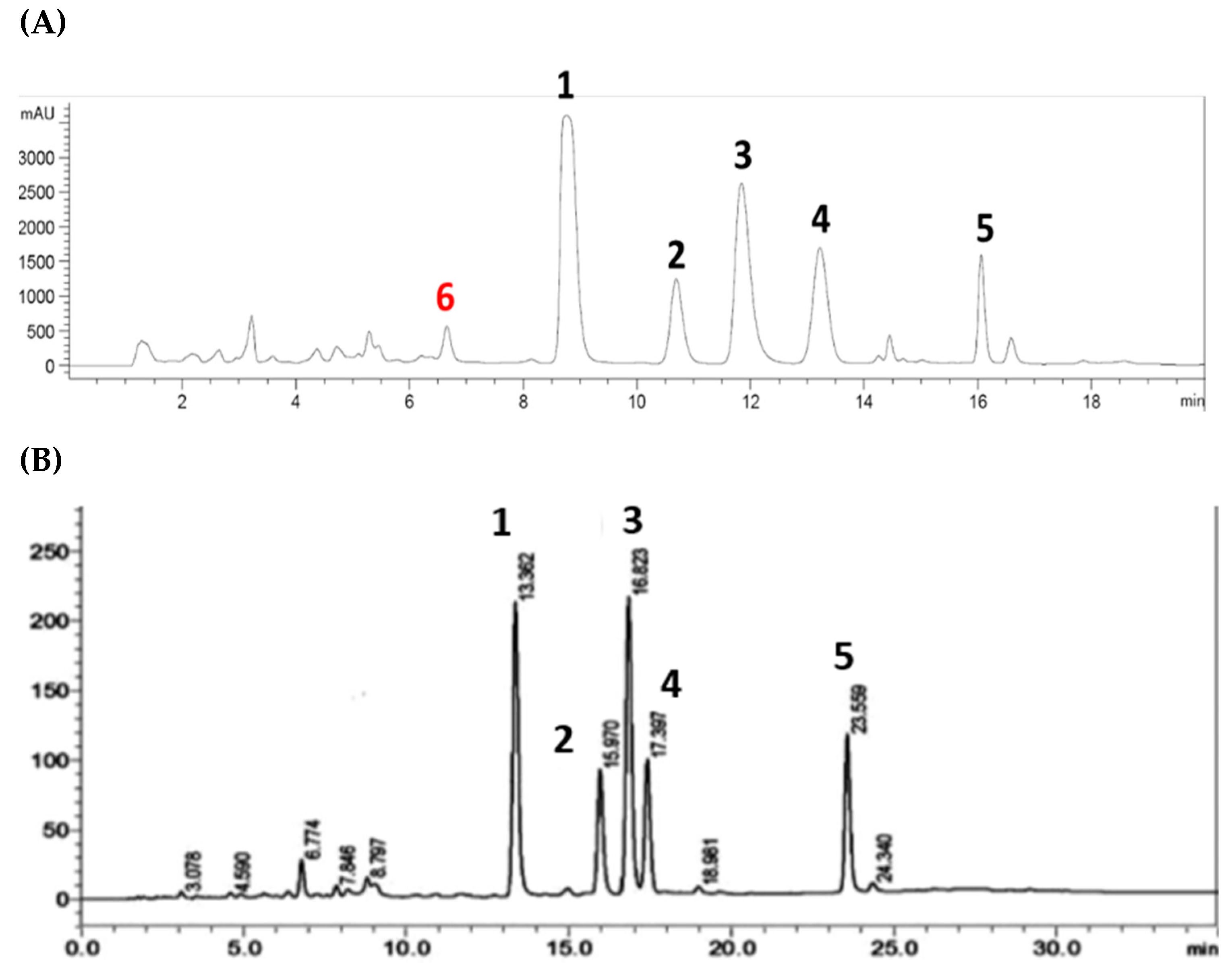
2.5. Analytical HPLC-DAD Analysis
2.6. LC–MS Analysis
2.7. NMR Experiments
2.8. α-Glucosidase Inhibition Assay
2.9. α-Amylase Inhibition Assay
2.10. Lipase Inhibition Assay
2.11. α-Glucosidase Binding Assay
2.12. Molecular Docking Study
3. Results and Discussion
3.1. α-Glucosidase Binding Assay
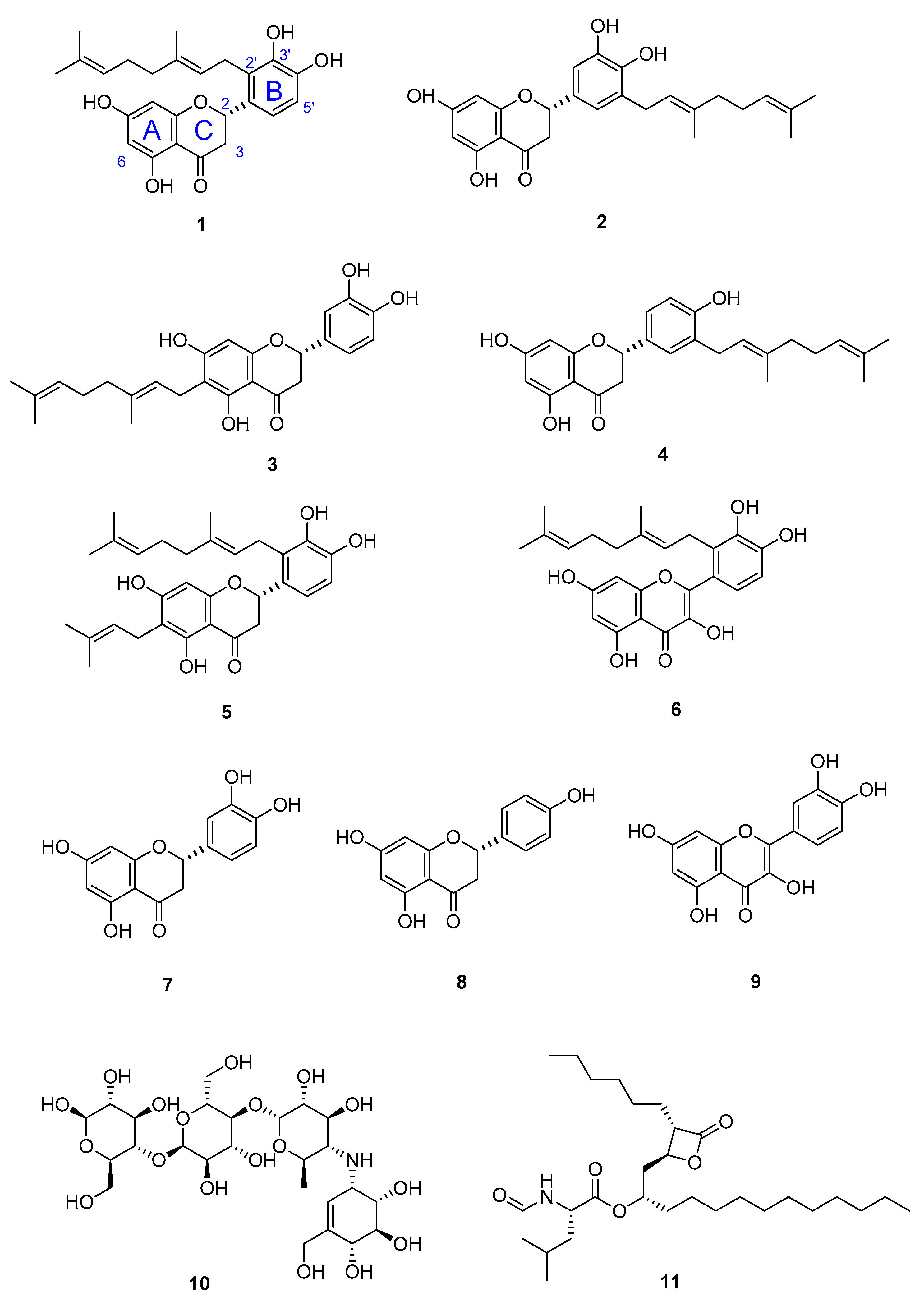
3.2. NMR Identification of α-Glucosidase Inhibitors
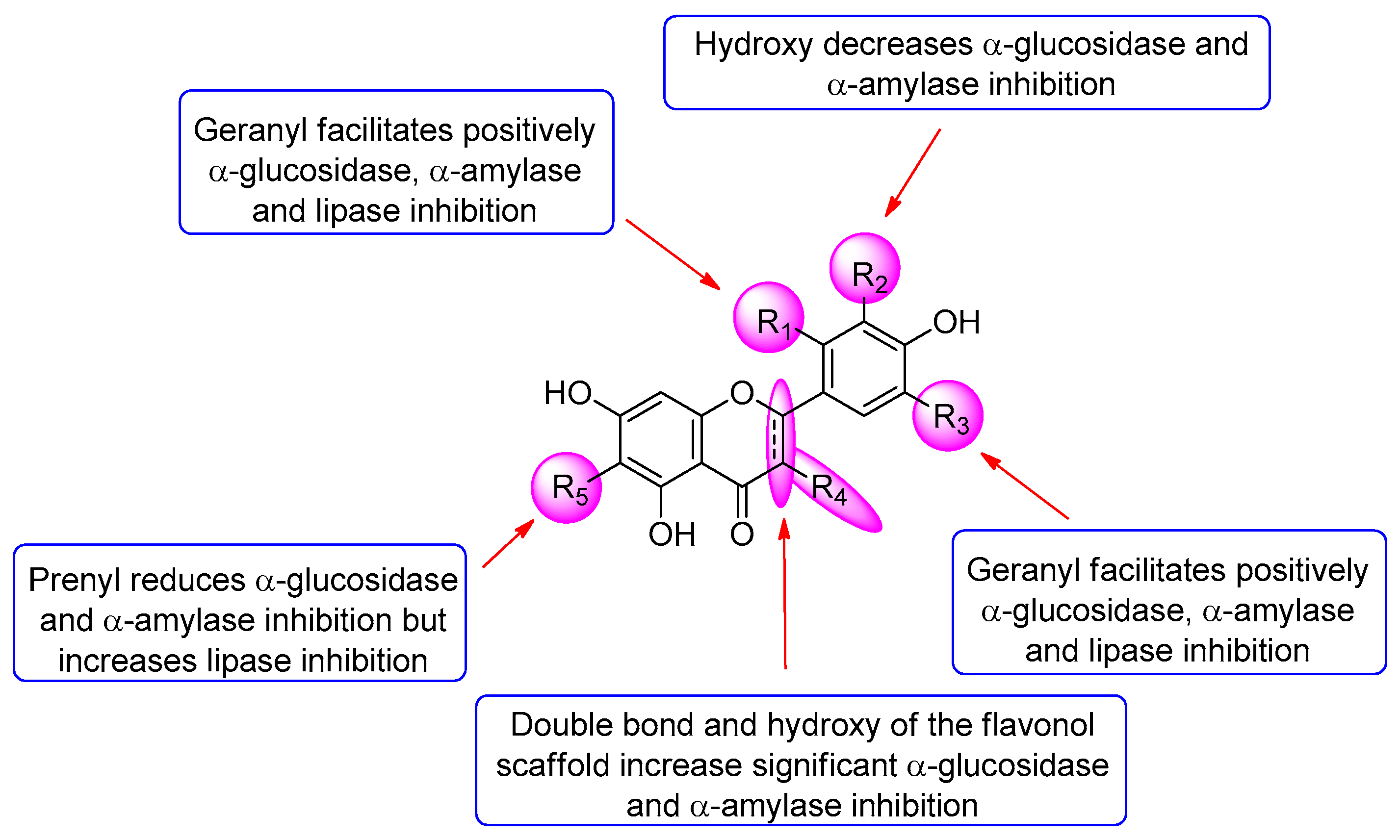
3.3. Inhibitory Activity Assessment of the Isolated Compounds
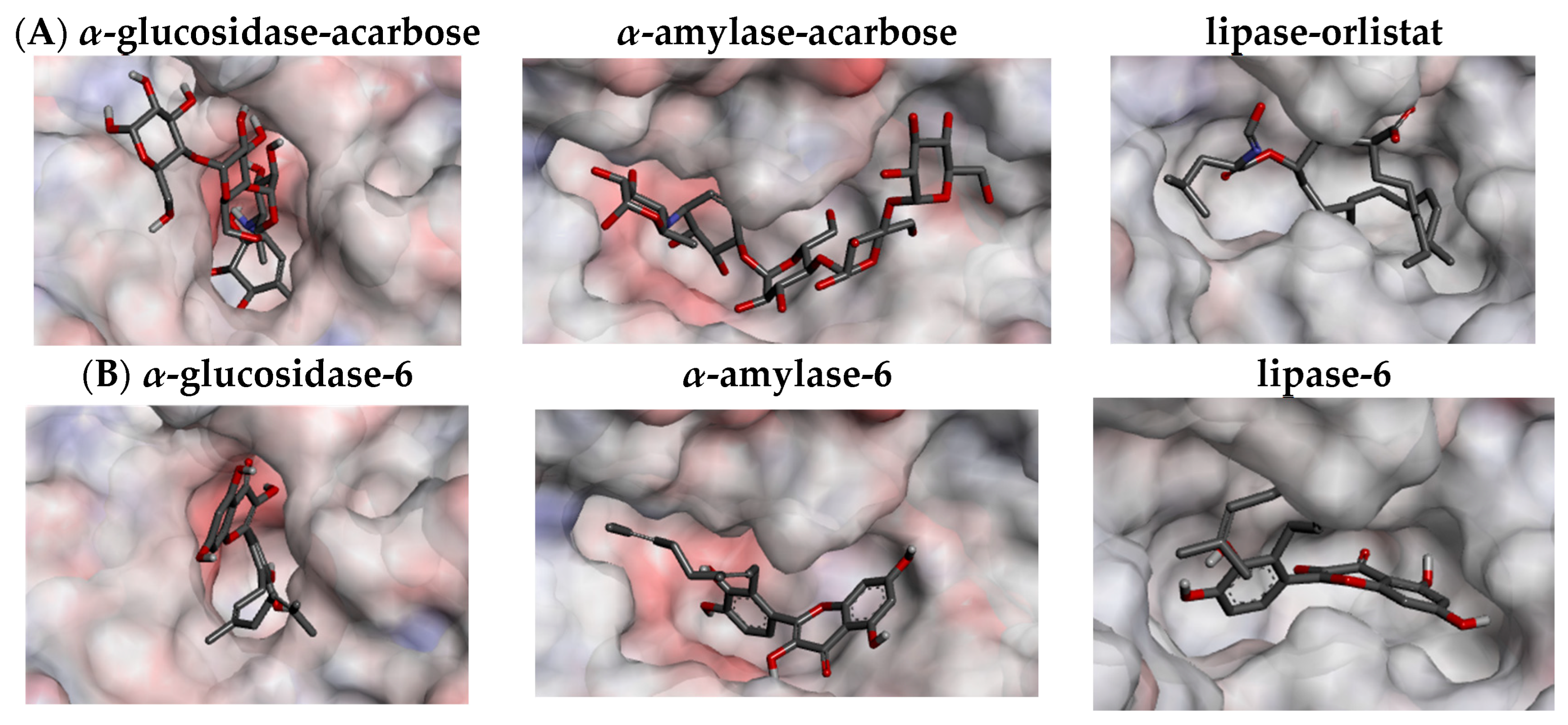
3.4. Ligand–Protein Interactions
3.5. Propolis Identification
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anderson, J.W.; Randles, K.M.; Kendall, C.W.C.; Jenkins, D.J.A. Carbohydrate and fiber recommendations for individuals with diabetes: A quantitative assessment and meta-analysis of the evidence. J. Am. Coll. Nutr. 2004, 23, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.; Min Cho, J.; Kwon, Y.-I.; Kim, S.-C.; Yeob Shin, D.; Ho Lee, J. Chitosan oligosaccharide (GO2KA1) improves postprandial glycemic response in subjects with impaired glucose tolerance and impaired fasting glucose and in healthy subjects: A crossover, randomized controlled trial. Nutr. Diabetes 2019, 9, 31. [Google Scholar] [CrossRef] [PubMed]
- Petersen, M.J.; de Cássia Lemos Lima, R.; Kjaerulff, L.; Staerk, D. Immobilized α-amylase magnetic beads for ligand fishing: Proof of concept and identification of α-amylase inhibitors in Ginkgo Biloba. Phytochem. 2019, 164, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Sainsbury, E.; Shi, Y.; Flack, J.; Colagiuri, S. Burden of Diabetes in Australia: It’s Time for More Action; The University of Sydney: Sydney, Australia, 2018; pp. 1–15. [Google Scholar]
- Lee, C.M.Y.; Colagiuri, R.; Magliano, D.J.; Cameron, A.J.; Shaw, J.; Zimmet, P.; Colagiuri, S. The cost of diabetes in adults in Australia. Diabetes Res. Clin. Pract. 2013, 99, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, A.K.; Rao, J.M. Diabetes mellitus and multiple therapeutic approaches of phytochemicals: Present status and future prospects. Curr. Sci. 2002, 83, 30–38. [Google Scholar]
- Hawash, M.; Jaradat, N.; Shekfeh, S.; Abualhasan, M.; Eid, A.M.; Issa, L. Molecular docking, chemo-informatic properties, alpha-amylase, and lipase inhibition studies of benzodioxol derivatives. BMC Chem. 2021, 15, 40. [Google Scholar] [CrossRef]
- Tomasik, P.; Horton, D. Chapter 2—Enzymatic conversions of starch. In Advances in Carbohydrate Chemistry and Biochemistry; Horton, D., Ed.; Academic Press: Cambridge, MA, USA, 2012; Volume 68, pp. 59–436. [Google Scholar]
- Hanefeld, M. Cardiovascular benefits and safety profile of acarbose therapy in prediabetes and established type 2 diabetes. Cardiovasc. Diabetol. 2007, 6, 20. [Google Scholar] [CrossRef] [Green Version]
- Poovitha, S.; Parani, M. In vitro and in vivo α-amylase and α-glucosidase inhibiting activities of the protein extracts from two varieties of bitter gourd (Momordica charantia L.). BMC Complement. Altern. Med. 2016, 16, 185. [Google Scholar] [CrossRef] [Green Version]
- Laar, F.A.V.D. Alpha-glucosidase inhibitors in the early treatment of type 2 diabetes. Vasc. Health Risk Manag. 2008, 4, 1189–1195. [Google Scholar] [CrossRef] [Green Version]
- Pujirahayu, N.; Bhattacharjya, D.K.; Suzuki, T.; Katayama, T. α-Glucosidase inhibitory activity of cycloartane-type triterpenes isolated from Indonesian stingless bee propolis and their structure–activity relationship. Pharmaceuticals 2019, 12, 102. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Chen, M.X.; Kongstad, K.T.; Jäger, A.K.; Staerk, D. Potential of Polygonum cuspidatum root as an antidiabetic food: Dual high-resolution α-glucosidase and PTP1B inhibition profiling combined with HPLC-HRMS and NMR for identification of antidiabetic constituents. J. Agric. Food Chem. 2017, 65, 4421–4427. [Google Scholar] [CrossRef] [PubMed]
- Mosleh, R.; Hawash, M.; Jarrar, Y. The relationships among the organizational factors of a tertiary healthcare center for type 2 diabetic patients in Palestine. Endocr. Metab. Immune Disord. Drug Targets 2021, 21, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Golay, A.; Ybarra, J. Link between obesity and type 2 diabetes. Best Pract. Res. Clin. Endocrinol. Metab. 2005, 19, 649–663. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, H. Effects of propolis extract and propolis-derived compounds on obesity and diabetes: Knowledge from cellular and animal Models. Molecules 2019, 24, 4394. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.Y.; Liu, M.; Portincasa, P.; Wang, D.Q.H. New insights into the molecular mechanism of intestinal fatty acid absorption. Eur. J. Clin. Investig. 2013, 43, 1203–1223. [Google Scholar] [CrossRef] [Green Version]
- Ballinger, A. Orlistat in the treatment of obesity. Expert Opin. Pharmacother. 2000, 1, 841–847. [Google Scholar] [CrossRef]
- Xiong, Y.; Ng, K.; Zhang, P.; Warner, R.D.; Shen, S.; Tang, H.-Y.; Liang, Z.; Fang, Z. In vitro α-glucosidase and α-amylase inhibitory activities of free and bound phenolic extracts from the bran and kernel fractions of five Sorghum grain genotypes. Foods 2020, 9, 1301. [Google Scholar] [CrossRef]
- Fu, C.; Jiang, Y.; Guo, J.; Su, Z. Natural products with anti-obesity effects and different mechanisms of action. J. Agric. Food Chem. 2016, 64, 9571–9585. [Google Scholar] [CrossRef]
- Lankatillake, C.; Luo, S.; Flavel, M.; Lenon, G.B.; Gill, H.; Huynh, T.; Dias, D.A. Screening natural product extracts for potential enzyme inhibitors: Protocols, and the standardisation of the usage of blanks in α-amylase, α-glucosidase and lipase assays. Plant Methods 2021, 17, 3. [Google Scholar] [CrossRef]
- Mohamed, G.A.; Ibrahim, S.R.M.; Elkhayat, E.S.; El Dine, R.S. Natural anti-obesity agents. Bull. Fac. Pharm. Cairo Univ. 2014, 52, 269–284. [Google Scholar] [CrossRef] [Green Version]
- Perera, P.K.; Li, Y. Functional herbal food ingredients used in type 2 diabetes mellitus. Pharm. Rev. 2012, 6, 37–45. [Google Scholar] [CrossRef] [Green Version]
- Kazeem, M.I.; Davies, T.C. Anti-diabetic functional foods as sources of insulin secreting, insulin sensitizing and insulin mimetic agents. J. Funct. Foods 2016, 20, 122–138. [Google Scholar] [CrossRef]
- Kawser Hossain, M.; Abdal Dayem, A.; Han, J.; Yin, Y.; Kim, K.; Kumar Saha, S.; Yang, G.-M.; Choi, H.Y.; Cho, S.-G. Molecular mechanisms of the anti-obesity and anti-diabetic properties of flavonoids. Int. J. Mol. Sci. 2016, 17, 569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boccellino, M.; D’Angelo, S. Anti-obesity effects of polyphenol intake: Current status and future possibilities. Int. J. Mol. Sci. 2020, 21, 5642. [Google Scholar] [CrossRef]
- Marcucci, M.C. Propolis: Chemical composition, biological properties and therapeutic activity. Apidologie 1995, 26, 83–99. [Google Scholar] [CrossRef]
- Tran, T.D.; Ogbourne, S.M.; Brooks, P.R.; Sánchez-Cruz, N.; Medina-Franco, J.L.; Quinn, R.J. Lessons from exploring chemical space and chemical diversity of propolis components. Int. J. Mol. Sci. 2020, 21, 4988. [Google Scholar] [CrossRef]
- Braakhuis, A. Evidence on the health benefits of supplemental propolis. Nutrients 2019, 11, 2705. [Google Scholar] [CrossRef] [Green Version]
- Tsuda, T.; Kumazawa, S. Propolis: Chemical constituents, plant origin, and possible role in the prevention and treatment of obesity and diabetes. J. Agric. Food Chem. 2021, 69, 15484–15494. [Google Scholar] [CrossRef]
- Fu, Y.; Luo, J.; Qin, J.; Yang, M. Screening techniques for the identification of bioactive compounds in natural products. J. Pharm. Biomed. Anal. 2019, 168, 189–200. [Google Scholar] [CrossRef]
- Wang, L.; Tan, N.; Wang, H.; Hu, J.; Diwu, W.; Wang, X. A systematic analysis of natural α-glucosidase inhibitors from flavonoids of Radix scutellariae using ultrafiltration UPLC-TripleTOF-MS/MS and network pharmacology. BMC Complement. Med. Ther. 2020, 20, 72. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Guo, M. Rapid screening for α-glucosidase inhibitors from Gymnema sylvestre by affinity ultrafiltration–HPLC-MS. Front. Pharmacol. 2017, 8, 228. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Xu, L.; Ye, Y.H.; Zhu, M.F.; Li, J.; Tu, Z.C.; Yang, S.H.; Liao, H. Phytochemical profiles and screening of α-glucosidase inhibitors of four Acer species leaves with ultra-filtration combined with UPLC-QTOF-MS/MS. Ind. Crops Prod. 2019, 129, 156–168. [Google Scholar] [CrossRef]
- Peng, M.J.; Shi, S.Y.; Chen, L.; Zhang, S.H.; Cai, P.; Chen, X.Q. Online coupling solid-phase ligand-fishing with high-performance liquid chromatography–diode array detector–tandem mass spectrometry for rapid screening and identification of xanthine oxidase inhibitors in natural products. Anal. Bioanal. Chem. 2016, 408, 6693–6701. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Zhang, Y.; Guo, Y.; Shi, S. Identification of major α-glucosidase inhibitors in Radix Astragali and its human microsomal metabolites using ultrafiltration HPLC–DAD–MSn. J. Pharm. Biomed. Anal. 2015, 104, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Wubshet, S.G.; Brighente, I.M.C.; Moaddel, R.; Staerk, D. Magnetic ligand fishing as a targeting tool for HPLC-HRMS-SPE-NMR: α-glucosidase inhibitory ligands and alkylresorcinol glycosides from Eugenia catharinae. J. Nat. Prod. 2015, 78, 2657–2665. [Google Scholar] [CrossRef] [Green Version]
- Lv, D.; Xu, J.; Qi, M.; Wang, D.; Xu, W.; Qiu, L.; Li, Y.; Cao, Y. A strategy of screening and binding analysis of bioactive components from traditional Chinese medicine based on surface plasmon resonance biosensor. J. Pharm. Anal. 2021, 12, 500–508. [Google Scholar] [CrossRef]
- Kumazawa, S.; Ueda, R.; Hamasaka, T.; Fukumoto, S.; Fujimoto, T.; Nakayama, T. Antioxidant prenylated flavonoids from propolis collected in Okinawa, Japan. J. Agric. Food Chem. 2007, 55, 7722–7725. [Google Scholar] [CrossRef]
- Kumazawa, S.; Goto, H.; Hamasaka, T.; Fukumoto, S.; Fujimoto, T.; Nakayama, T. A new prenylated flavonoid from propolis collected in Okinawa, Japan. Biosci. Biotechnol. Biochem. 2004, 68, 260–262. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.N.; Wu, C.L.; Lin, J.K. Propolin C from propolis induces apoptosis through activating caspases, Bid and cytochrome c release in human melanoma cells. Biochem. Pharmacol. 2004, 67, 53–66. [Google Scholar] [CrossRef]
- Inui, S.; Hosoya, T.; Shimamura, Y.; Masuda, S.; Ogawa, T.; Kobayashi, H.; Shirafuji, K.; Moli, R.T.; Kozone, I.; Shin-ya, K.; et al. Solophenols B–D and solomonin: New prenylated polyphenols isolated from propolis collected from the Solomon Islands and their antibacterial activity. J. Agric. Food Chem. 2012, 60, 11765–11770. [Google Scholar] [CrossRef]
- Venturelli, S.; Burkard, M.; Biendl, M.; Lauer, U.M.; Frank, J.; Busch, C. Prenylated chalcones and flavonoids for the prevention and treatment of cancer. Nutrition 2016, 32, 1171–1178. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, K.; Tsurumaru, Y.; Yamamoto, H.; Yazaki, K. Molecular characterization of a membrane-bound prenyltransferase specific for isoflavone from Sophora flavescens. J. Biol. Chem. 2011, 286, 24125–24134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quiroga, E.N.; Sampietro, D.A.; Sgariglia, M.A.; Soberón, J.R.; Vattuone, M.A. Antimycotic activity of 5′-prenylisoflavanones of the plant Geoffroea decorticans, against Aspergillus species. Int. J. Food Microbiol. 2009, 132, 42–46. [Google Scholar] [CrossRef]
- Edziri, H.; Mastouri, M.; Mahjoub, M.A.; Mighri, Z.; Mahjoub, A.; Verschaeve, L. Antibacterial, antifungal and cytotoxic activities of two flavonoids from Retama raetam Flowers. Molecules 2012, 17, 7284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sohn, H.Y.; Son, K.H.; Kwon, C.S.; Kwon, G.S.; Kang, S.S. Antimicrobial and cytotoxic activity of 18 prenylated flavonoids isolated from medicinal plants: Morus alba L., Morus mongolica Schneider, Broussnetia papyrifera (L.) Vent, Sophora flavescens Ait and Echinosophora koreensis Nakai. Phytomedicine 2004, 11, 666–672. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; He, Z.-D.; Jiang, R.-W.; Ye, W.-C.; Xu, H.-X.; But, P.P.-H. Antiviral flavonoids from the root bark of Morus alba L. Phytochemistry 2003, 62, 1235–1238. [Google Scholar] [CrossRef]
- Liu, M.; Hansen, P.E.; Wang, G.; Qiu, L.; Dong, J.; Yin, H.; Qian, Z.; Yang, M.; Miao, J. Pharmacological profile of xanthohumol, a prenylated flavonoid from Hops (Humulus lupulus). Molecules 2015, 20, 754. [Google Scholar] [CrossRef]
- Jin, J.H.; Kim, J.S.; Kang, S.S.; Son, K.H.; Chang, H.W.; Kim, H.P. Anti-inflammatory and anti-arthritic activity of total flavonoids of the roots of Sophora flavescens. J. Ethnopharmacol. 2010, 127, 589–595. [Google Scholar] [CrossRef]
- Molčanová, L.; Janošíková, D.; Dall´Acqua, S.; Šmejkal, K. C-prenylated flavonoids with potential cytotoxic activity against solid tumor cell lines. Phytochem. Rev. 2019, 18, 1051–1100. [Google Scholar] [CrossRef]
- De Naeyer, A.; Vanden Berghe, W.; Pocock, V.; Milligan, S.; Haegeman, G.; De Keukeleire, D. Estrogenic and anticarcinogenic properties of kurarinone, a lavandulyl flavanone from the roots of Sophora flavescens. J. Nat. Prod. 2004, 67, 1829–1832. [Google Scholar] [CrossRef]
- Dong, X.; Fan, Y.; Yu, L.; Hu, Y. Synthesis of four natural prenylflavonoids and their estrogen-like activities. Arch. Pharm. Chem. Life Sci. 2007, 340, 372–376. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.-G.; Matsuzaki, K.; Takamatsu, S.; Kitanaka, S. Inhibitory effects of constituents from Morus alba var. multicaulis on differentiation of 3T3-L1 cells and nitric oxide production in RAW264.7 cells. Molecules 2011, 16, 6010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Ryu, Y.B.; Kang, N.S.; Lee, B.W.; Heo, J.S.; Jeong, I.-Y.; Park, K.H. Glycosidase inhibitory flavonoids from Sophora flavescens. Biol. Pharm. Bull. 2006, 29, 302–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osorio, M.; Carvajal, M.; Vergara, A.; Butassi, E.; Zacchino, S.; Mascayano, C.; Montoya, M.; Mejías, S.; Martín, M.C.; Vásquez-Martínez, Y. Prenylated flavonoids with potential antimicrobial activity: Synthesis, biological activity, and in slico study. Int. J. Mol. Sci. 2021, 22, 5472. [Google Scholar] [CrossRef]
- Yazaki, K.; Sasaki, K.; Tsurumaru, Y. Prenylation of aromatic compounds, a key diversification of plant secondary metabolites. Phytochemistry 2009, 70, 1739–1745. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.W.; Ye, S.; Ting, C.; Yu, Y.H. Antibacterial activity of propolins from Taiwanese green propolis. J. Food Drug Anal. 2018, 26, 761–768. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Kim, Y.G.; Khadke, S.K.; Yamano, A.; Woo, J.T.; Lee, J. Antimicrobial and antibiofilm activities of prenylated flavanones from Macaranga tanarius. Phytomedicine 2019, 63, 153033. [Google Scholar] [CrossRef]
- Shahinozzaman, M.; Taira, N.; Ishii, T.; Halim, M.A.; Hossain, M.A.; Tawata, S. Anti-inflammatory, Anti-diabetic, and Anti-Alzheimer’s effects of prenylated flavonoids from Okinawa propolis: An investigation by experimental and computational studies. Molecules 2018, 23, 2479. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.N.; Weng, M.S.; Wu, C.L.; Lin, J.K. Comparison of radical scavenging activity, cytotoxic effects and apoptosis induction in human melanoma cells by Taiwanese propolis from different sources. Evid. Based Complement. Alternat. Med. 2004, 1, 375152. [Google Scholar] [CrossRef] [Green Version]
- Weng, M.S.; Liao, C.H.; Chen, C.N.; Wu, C.L.; Lin, J.K. Propolin H from Taiwanese propolis induces G1 arrest in human lung carcinoma cells. J. Agric. Food Chem. 2007, 55, 5289–5298. [Google Scholar] [CrossRef]
- Hsieh, C.-Y.; Li, L.-H.; Rao, Y.K.; Ju, T.-C.; Nai, Y.-S.; Chen, Y.-W.; Hua, K.-F. Mechanistic insight into the attenuation of gouty inflammation by Taiwanese green propolis via inhibition of the NLRP3 inflammasome. J. Cell. Physiol. 2019, 234, 4081–4094. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.J.; Huang, C.H.; Wu, C.L.; Lin, J.K.; Chen, Y.W.; Lin, C.L.; Chuang, S.E.; Huang, C.Y.; Chen, C.N. Propolin G, a prenylflavanone, isolated from Taiwanese propolis, induces caspase-dependent apoptosis in brain cancer cells. J. Agric. Food Chem. 2007, 55, 7366–7376. [Google Scholar] [CrossRef] [PubMed]
- Su, K.Y.; Hsieh, C.Y.; Chen, Y.W.; Chuang, C.T.; Chen, C.T.; Chen, Y.L.S. Taiwanese green propolis and propolin G protect the liver from the pathogenesis of fibrosis via eliminating TGF-β-induced smad2/3 phosphorylation. J. Agric. Food Chem. 2014, 62, 3192–3201. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.D.; Bui, T.Q.; Le, T.A.; Nguyen, M.T.; Hai, N.T.T.; Pham, N.H.; Phan, M.N.; Healy, P.C.; Pham, N.B.; Quinn, R.J.; et al. Styracifoline from the Vietnamese Plant Desmodium styracifolium: A Potential Inhibitor of Diabetes-Related and Thrombosis-Based Proteins. ACS Omega 2021, 6, 23211–23221. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.H.; Chien, Y.W.; Chang, M.L.; Hou, C.C.; Chan, C.H.; Tang, H.W.; Huang, H.Y. Taiwanese green propolis ethanol extract delays the progression of type 2 diabetes mellitus in rats treated with streptozotocin/high-fat diet. Nutrients 2018, 10, 503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumazawa, S.; Nakamura, J.; Murase, M.; Miyagawa, M.; Ahn, M.-R.; Fukumoto, S. Plant origin of Okinawan propolis: Honeybee behavior observation and phytochemical analysis. Naturwissenschaften 2008, 95, 781. [Google Scholar] [CrossRef]
- NatureWise Biotech & Medicals Corporation. Available online: https://www.naturewiseppls.com/en/about-propolis/ (accessed on 1 June 2022).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Retention Time (min) | (+) m/z [M+H]+ | (−) m/z [M-H]− | Predicted MW |
|---|---|---|---|
| 6.267 | 439 | 437 | 438 |
| 6.793 | 425 | 423 | 424 |
| 7.133 | 425 | 423 | 424 |
| 7.300 | 425 | 423 | 424 |
| 7.360 | 409 | 407 | 408 |
| 7.687 | 409 | 407 | 408 |
| 8.327 | 493 | 491 | 492 |
| 8.493 | 493 | 491 | 492 |
| Compound | IC50 (µM) | ||
|---|---|---|---|
| α-Glucosidase | α-Amylase | Lipase | |
| 1 | 252.4 | 204.6 | 51.8 |
| 2 | 298.4 | 134.9 | 53.0 |
| 3 | 421.3 | 203.9 | 30.3 |
| 4 | 178.5 | 52.5 | 33.1 |
| 5 | 776.6 | 246.0 | 43.4 |
| 6 | 57.8 | 142.1 | 32.3 |
| 7 | 13% at 250 µM | 41% at 250 µM | 12% at 83 µM |
| 8 | 647.4 c | 121.5 d | 24% at 83 µM |
| 9 | 19.2 | 17.4 | 37% at 83 µM |
| Acarbose a | 396.7 | 624.7 | - b |
| Orlistat a | - b | - b | 5.4 |
| Compounds | Biological Properties |
|---|---|
| Propolin D (1) | Antioxidant [39], antimicrobial [58,59], anti-inflammatory [60], anti-Alzheimer’s [60], anticancer [61], α-glucosidase inhibition [60], α-amylase inhibition *, lipase inhibition * |
| Propolin F (2) | Antioxidant [39], antimicrobial [58,59], anti-inflammatory [60], anti-Alzheimer’s [60], anticancer [61], α-glucosidase inhibition [60], α-amylase inhibition *, lipase inhibition * |
| Propolin C (3) | Antioxidant [39], antimicrobial [58,59], anti-inflammatory [60], anti-Alzheimer’s [60], anticancer [41,61], α-glucosidase inhibition [60], α-amylase inhibition *, lipase inhibition * |
| Propolin H (4) | Antioxidant [39], antimicrobial [59], anti-inflammatory [60], anti-Alzheimer’s [60], anticancer [62], α-glucosidase inhibition [60], α-amylase inhibition *, lipase inhibition * |
| Propolin G (5) | Antioxidant [39], antimicrobial [58,59], anti-inflammatory [60,63], anti-Alzheimer’s [60], anticancer [64], hepatoprotective [65], α-glucosidase inhibition [60], α-amylase inhibition *, lipase inhibition * |
| Solophenol D (6) | Antibacterial [42], α-glucosidase inhibition *, α-amylase inhibition *, lipase inhibition * |
| Compound | Binding Affinity (kcal/mol) | Hydrogen Bond (Distance, Å) | Hydrophobic Bond (Distance, Å) | Electrostatic Bond (Distance, Å) | Total Interactions |
|---|---|---|---|---|---|
| α-glucosidase (2ZQ0) | |||||
| 1 | −9.4 | Met334 (1.93); Glu508 (1.90); Glu532 (2.77) | π-π: Phe536 (4.89) π-alkyl: Tyr533 (5.02); Phe536 (4.46); Phe536 (4.83); Val471 (4.80) | Glu532 (3.91) | 9 |
| 2 | −9.3 | Asn216 (2.55); His437 (3.09); Ser217 (1.55); Glu391 (2.81) | π-alkyl: Phe536 (5.02); Phe536 (4.66); Phe536 (4.83); Val471 (5.18); Val471 (5.48) | Glu439 (3.99) | 10 |
| 3 | −9.8 | Glu508 (2.59); Glu391 (1.89); Trp341 (2.41) | π-π: Phe536 (4.24) π-alkyl: Phe536 (4.44); Val471 (5.24) | 6 | |
| 4 | −9.3 | Ser217 (2.31); Glu532 (2.16) | π-σ: Val471 (3.74); Phe536 (3.82) π-π: Trp341 (5.41); Phe401 (5.21) π-alkyl: Phe536 (4.03) | 7 | |
| 5 | −9.6 | Glu508 (2.09); Glu508 (2.79); Glu194 (3.08); Met334 (2.07) | π-π: Phe536 (4.85) π-alkyl: Tyr533 (5.02); Phe536 (4.47); Phe536 (4.19); Val471 (4.94) | Glu532 (3.80) | 10 |
| 6 | −9.2 | Ser217 (1.88); Ser217 (2.60); Ser217 (2.07); Ser217 (2.62); Glu194 (3.06) | π-σ: Val471 (3.91) π-alkyl: Trp341 (5.45); Trp341 (4.93); Trp400 (5.09); Phe401 (4.84); Phe401 (4.19); Phe536 (4.64) Alkyl: Ile335 (4.61) | Glu532 (3.84) | 14 |
| 7 | −8.9 | Lys467 (2.64); Glu391 (2.61); Ser217 (1.11) | π-alkyl: Val471 (5.49) | Glu439 (4.47) Glu532 (4.17) | 6 |
| 8 | −8.4 | Ser217 (2.28); Glu194 (2.65) | π-π: Trp341 (4.76) π-alkyl: Val471 (4.35) | 4 | |
| 9 | −8.4 | Ser217 (2.43); Trp331 (2.12); Trp331 (2.30); Glu391 (2.12); Asn216 (2.87); His437 (3.42) | π-π: Trp341 (5.47); Trp400 (5.53) π-alkyl: Val471 (5.17); Val471 (5.32) | 10 | |
| Acarbose (10) b | −9.7 | Asn216 (2.86); Trp331 (2.01); Trp341 (2.67); His507 (2.89); Glu532 (3.06); Glu532 (2.40); Glu508 (2.11); Glu526 (2.37); Glu391 (2.33); His437 (2.34); Pro215 (2.24); Pro215 (2.35); Pro215 (3.71); Phe536 (3.42); Phe536 (2.66); Ser217 (2.00) | π-alkyl: Phe536 (5.31) | 17 | |
| α-Amylase (2QV4) | |||||
| 1 | −10.4 | Gln63 (2.69); Asp197 (3.02); His299 (2.45) | π-π: Trp59 (3.84); Trp59 (4.21); Tyr62 (4.30) π-alkyl: His201 (4.93) Alkyl: Leu162 (4.11); Leu162 (4.39); Lys200 (4.49); Ile235 (4.64) | Asp300 (3.79) | 12 |
| 2 | −9.5 | Gln63 (2.47); Gln63 (1.33); Asp197 (2.36); His299 (2.34); Glu233 (2.68) | π-π: Trp59 (4.16); Trp59 (4.14); Tyr62 (4.67) Alkyl: Leu162 (5.41); Ala198 (4.89) | Asp197 (4.94) | 11 |
| 3 | −10.0 | Gln63 (2.56); Asp300 (2.05); Asp197 (2.95) | π-π: Trp59 (4.22); Trp59 (4.22); Tyr62 (4.22) π-alkyl: Trp59 (4.49) Alkyl: Ile51 (3.77); Val107 (3.97) | 9 | |
| 4 | −9.5 | Gln63 (2.55); His299 (2.84); Asp300 (2.21) | π-π: Trp59 (3.86); Trp59 (4.09); Tyr62 (4.45) Alkyl: Leu162 (5.48); Ala198 (4.90) | 8 | |
| 5 | −9.4 | His305 (2.62); His201 (2.41); His305 (3.24) | π-σ: Tyr62 (3.74); Trp59 (3.82) π-alkyl: Trp59 (3.95); Trp59 (4.71); Trp59 (4.34); Tyr62 (5.32); Leu162 (4.89); Ile235 (5.33) Alkyl: Ala198 (5.23) | 12 | |
| 6 | −10.0 | Gln63 (1.99); Asp300 (2.95); Asp197 (2.38); Arg195(2.13) | π-π: Trp59 (4.98); Trp59 (4.65); Trp59 (4.60); Tyr62 (4.48) π-alkyl: His201 (4.97) Alkyl: Leu162 (4.25); Lys200 (4.40); Ile235 (4.37) | Asp300 (3.71) | 13 |
| 7 | −9.0 | Gln63 (2.47); Gln63 (2.27); His299 (2.75); Asp300 (2.48); His305 (3.65) | π-π: Trp59 (3.86); Trp59 (4.12); Tyr62 (4.27) | 8 | |
| 8 | −8.9 | Gln63 (2.39); Asp197 (2.02); His305 (3.62); Arg195(2.23) | π-π: Trp59 (3.86); Trp59 (4.10); Tyr62 (4.11) | 7 | |
| 9 | −9.0 | Gln63 (2.20); Gln63 (2.61); Asp300 (3.06); Tyr62 (2.74); His305 (3.58) | π-π: Trp59 (5.09); Trp59 (3.87); Trp59 (5.40); Trp59 (4.03); Tyr62 (4.54) | 10 | |
| Acarbose (10) b | −10.1 | Gln63 (1.97); Asn105 (2.50); Asn105 (2.63); Ala106 (1.94); Thr163 (2.85); Arg195(2.23); His305 (2.76); Glu233 (3.29); Asp300 (2.91); Gly164 (3.39); Thr163 (3.56); Glu233 (3.61); His305 (3.83); Tyr62 (3.50) | 14 | ||
| Lipase (6KSM) | |||||
| 1 | −9.4 | Phe17 (2.32); Phe17 (2.31); Phe17 (2.92); Ala175 (2.28) | π-σ: Val309 (3.87) π-alkyl: Phe17 (5.41); Phe59 (4.93); Phe178 (5.24); Ala174 (4.81); Ala175 (5.41) Alkyl: Leu18 (5.33); Leu18 (4.12); Leu18 (4.49); Met188 (4.34) | 14 | |
| 2 | −10.1 | His115 (2.64); Tyr32 (2.28); Tyr32 (2.87); Ser172 (3.12) | π-π: His349 (4.64) π-alkyl: Tyr32 (4.56); Phe285(4.91); Ala174 (5.04); Ala175 (4.90); Leu242 (4.22); Val309 (4.81) Alkyl: Pro30 (4.73); Pro30 (4.25); Val350 (4.63); Val355 (3.86); Val355 (4.62); Leu287 (4.30); Ile353 (5.45) | 18 | |
| 3 | −9.2 | Ala175 (2.90); Leu242 (2.74); Tyr32 (2.60); Ser172 (2.96) | π-π: His349 (5.91) π-alkyl: Phe285(4.75); Leu18 (5.37); Pro168 (4.98); Leu242 (4.39); Val309 (4.60); Val310 (5.48) Alkyl: Pro30 (5.32); Val350 (5.01); Val355 (4.94); Leu287 (5.29) | 15 | |
| 4 | −9.5 | π-σ: Val309 (3.92) π-alkyl: Phe17 (5.36); Phe17 (4.65); Phe178 (5.41); Leu242 (5.20); Met288 (5.27); Val350 (5.03) Alkyl: Ala174 (4.69); Ala175 (3.98); Ala175 (3.79); Ala239 (4.33); Leu242 (4.90); Leu242 (4.73); Val309 (5.29) | 14 | ||
| 5 | −10.0 | Phe17 (2.29); Phe17 (2.84); Phe17 (2.90); Ala175 (2.35) | π-alkyl: Phe17 (5.32); Tyr29 (4.46); Phe59 (5.28); Phe178 (5.35); Ala174 (5.06); Ala175 (5.30); Leu242 (4.56); Val309 (4.47) Alkyl: Leu18 (5.21); Leu18 (4.35); Met188 (4.25); Pro30 (4.76) | 16 | |
| 6 | −9.1 | π-σ: Val309 (3.72); Val309 (3.94) π-π: Phe17 (5.27); His349 (4.35); His349 (5.68) π-alkyl: Ala174 (4.75); Ala175 (5.23); Val309 (5.07) Alkyl: Met288 (4.61); Val350 (5.34); Val350 (5.24) | 11 | ||
| 7 | −8.6 | Gly16 (3.07); Leu242 (2.87) | π-π: His349 (4.45) π-alkyl: Ala174 (5.03); Ala175 (4.88); Leu242 (4.21); Val309 (4.82) | 7 | |
| 8 | −8.1 | π-π: Phe17 (5.59) π-alkyl: Ala174 (4.90); Ala175 (4.91); Leu242 (4.55); Val309 (4.83) | 5 | ||
| 9 | −8.6 | Phe17 (2.92); Ala175 (2.74); Tyr32 (2.53) | π-σ: Val309 (3.86) π-π: His349 (4.36); His349 (5.69) π-alkyl: Ala174 (4.73); Ala175 (5.24); Val309 (5.04) | 9 | |
| Orlistat (11) b | −7.0 | His115 (3.71); His349 (3.41) | π-alkyl: Phe17 (5.40); Phe17 (5.48); Tyr32 (4.86); Phe178 (5.00); His349 (4.72) Alkyl: Leu18 (5.46); Leu18 (4.42); Pro168 (4.22); Ala174 (4.27); Ala175 (3.94); Val309 (4.73); Val309 (5.34); Val310 (5.12); Val355 (5.35); Met188 (5.10); Leu242 (4.08); Leu242 (4.64) | 19 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uddin, S.; Brooks, P.R.; Tran, T.D. Chemical Characterization, α-Glucosidase, α-Amylase and Lipase Inhibitory Properties of the Australian Honey Bee Propolis. Foods 2022, 11, 1964. https://doi.org/10.3390/foods11131964
Uddin S, Brooks PR, Tran TD. Chemical Characterization, α-Glucosidase, α-Amylase and Lipase Inhibitory Properties of the Australian Honey Bee Propolis. Foods. 2022; 11(13):1964. https://doi.org/10.3390/foods11131964
Chicago/Turabian StyleUddin, Sabah, Peter R. Brooks, and Trong D. Tran. 2022. "Chemical Characterization, α-Glucosidase, α-Amylase and Lipase Inhibitory Properties of the Australian Honey Bee Propolis" Foods 11, no. 13: 1964. https://doi.org/10.3390/foods11131964
APA StyleUddin, S., Brooks, P. R., & Tran, T. D. (2022). Chemical Characterization, α-Glucosidase, α-Amylase and Lipase Inhibitory Properties of the Australian Honey Bee Propolis. Foods, 11(13), 1964. https://doi.org/10.3390/foods11131964