
Advances in Evaluation of Antioxidant and Toxicological Properties of Stryphnodendron rotundifolium Mart. in Drosophila melanogaster Model

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Botanical Material and Extraction Procedure
2.2. Total Flavonoid Content
2.3. Stock of Flies
Whole Fly Homogenate Preparation
2.4. Analysis of In Vitro Antioxidant Activity
2.4.1. ABTS•+ Free Radical Capture
2.4.2. Iron-Reducing Antioxidant Power (FRAP)
2.4.3. DPPH● Assay
2.4.4. Relative Antioxidant Capacity (RAC)
2.4.5. Inhibition of Lipid Peroxidation
2.5. In Vivo Toxicity Assays
2.5.1. Aging and Survival Assays
2.5.2. Startle-Induced Negative Geotaxis
2.5.3. Video-Assisted Open Field Motion Tracking
2.6. Statistical Analysis
3. Results and Discussion
3.1. Total Flavonoid Content and In Vitro Antioxidant Activity ABTS•+, FRAP, DPPH●, RAC
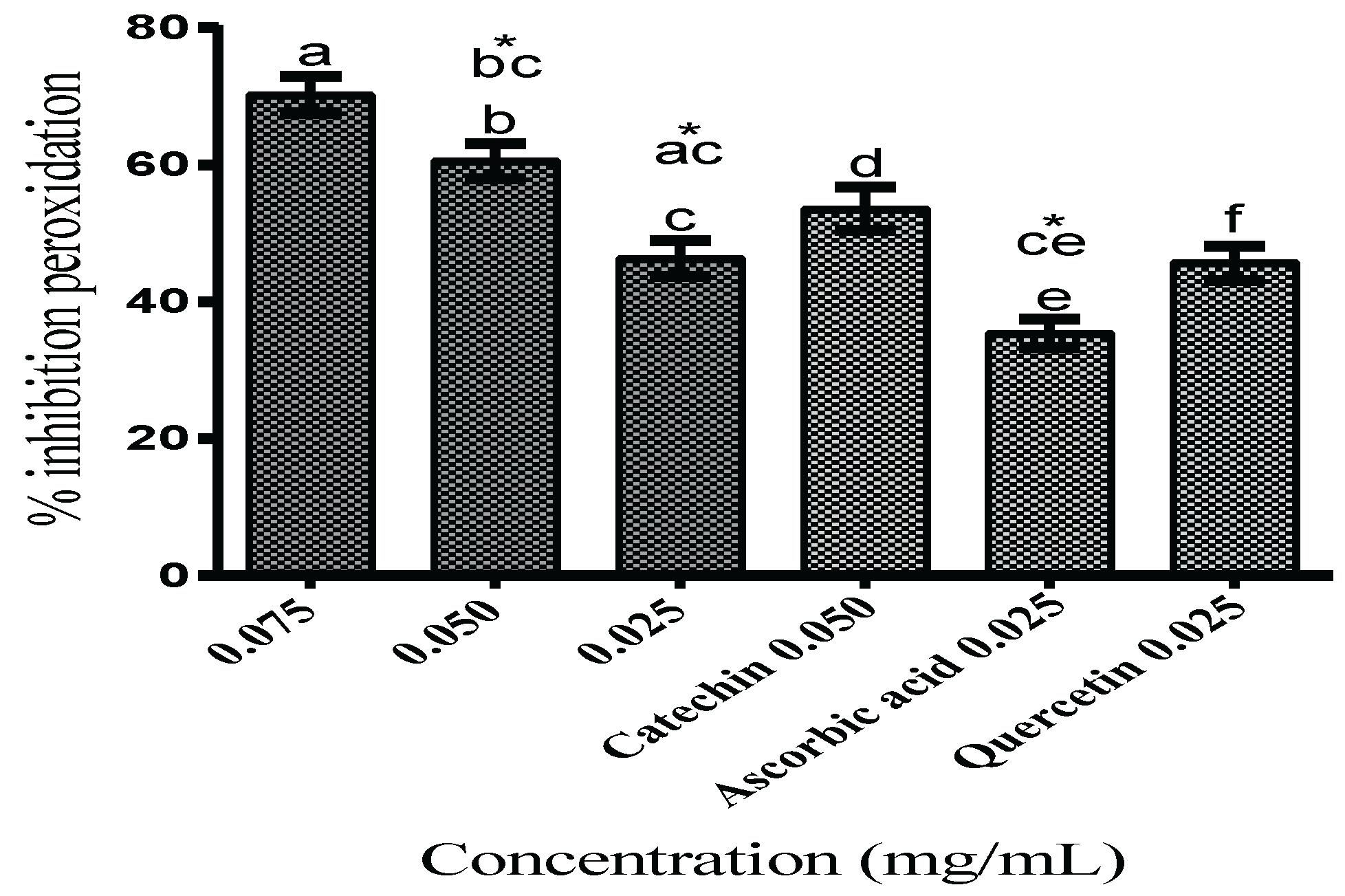
Lipid Peroxidation Inhibition
3.2. Toxicity and Locomotor Functions
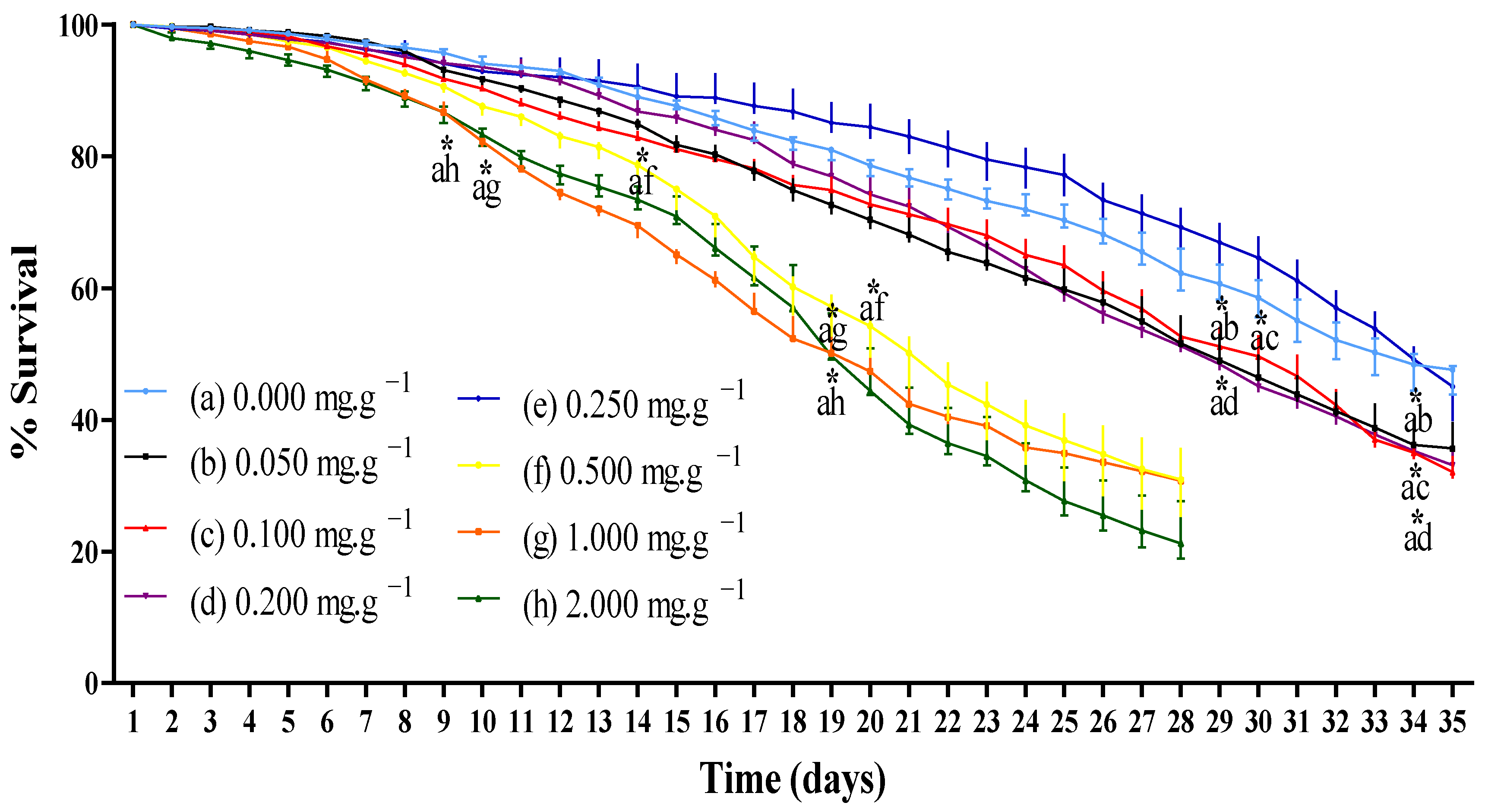
3.2.1. Survival Rate
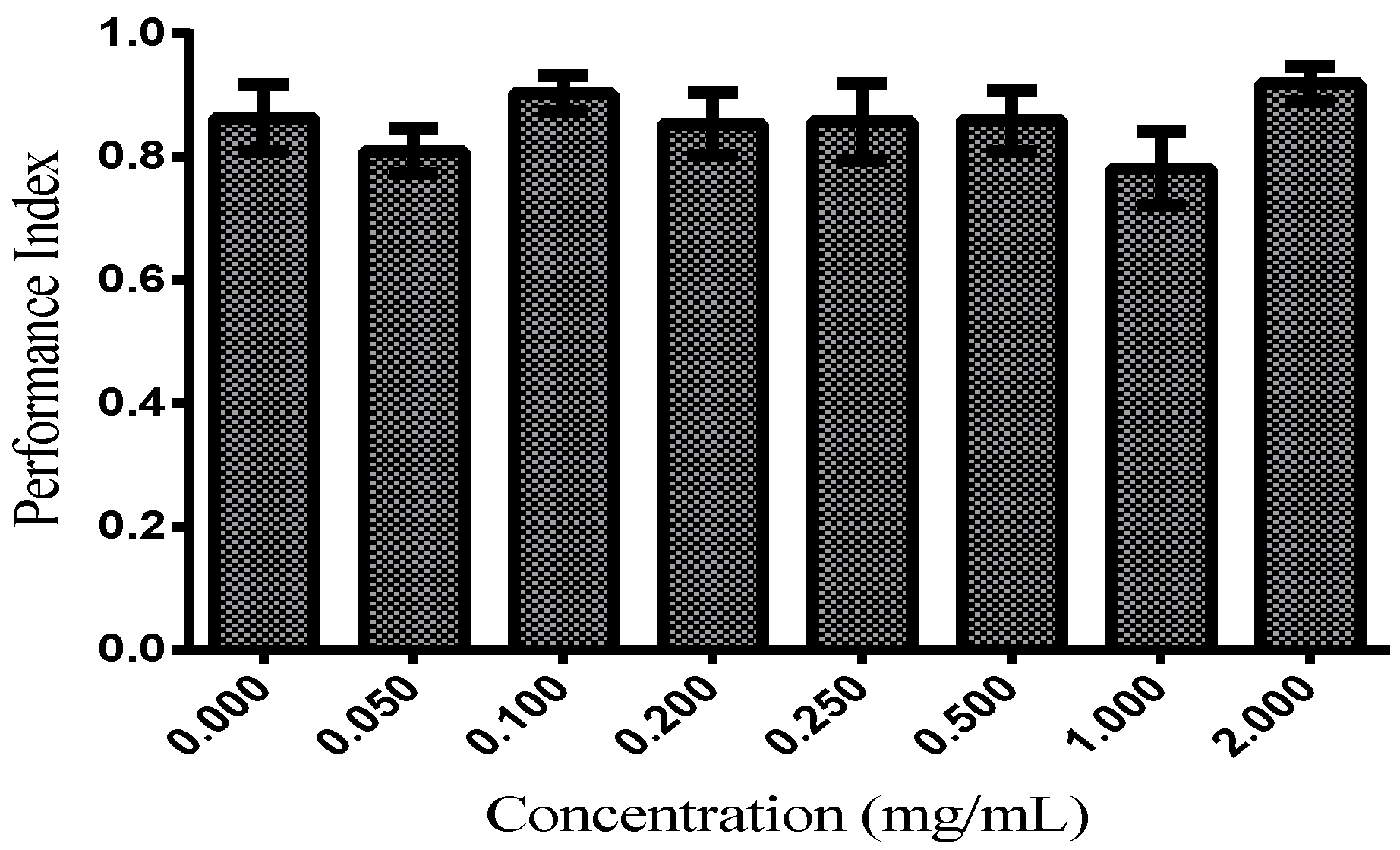
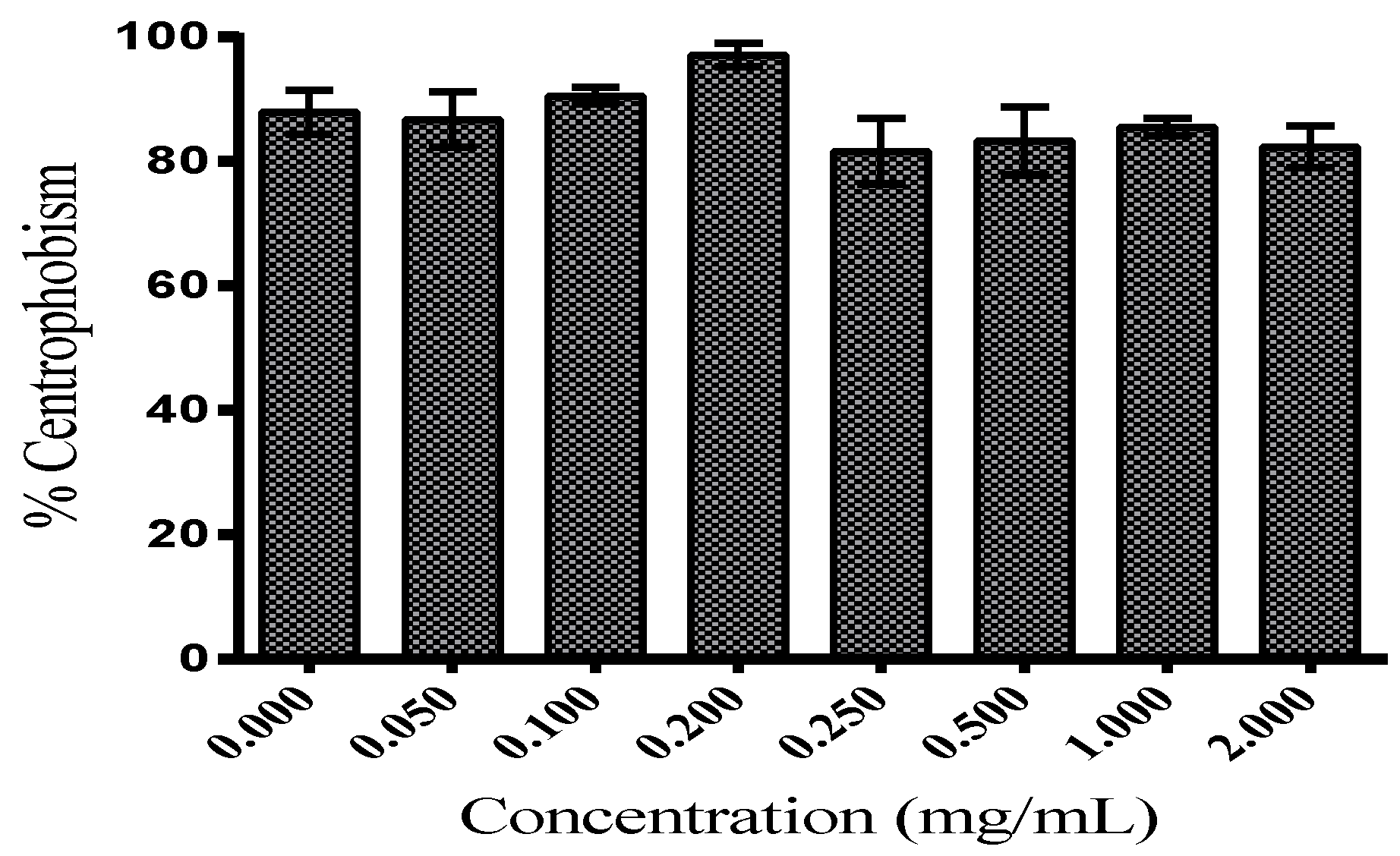
3.2.2. Locomotor Functions
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oliveira, D.R.; Washington, S.F.J.; Bitu, V.C.N.; Pinheiro, P.G.; Menezes, C.D.A.; Brito, J.F.E.; Albuquerque, U.P.; Kerntopf, M.R.; Coutinho, H.D.M.; Fachinetto, R.; et al. Ethnopharmacological study of Stryphnodendron rotundifolium in two communities in the semi-arid region of northeastern Brazil. Rev. Bras. Farmacogn. 2014, 24, 124–132. [Google Scholar] [CrossRef] [Green Version]
- de Carvalho Nilo Bitu, V.; de Carvalho Nilo Bitu, V.; Matias, E.F.F.; de Lima, W.P.; Portelo, A.d.C.; Coutinho, H.D.M.; de Menezes, I.R.A. Ethnopharmacological study of plants sold for therapeutic purposes in public markets in northeast Brazil. J. Ethnopharmacol. 2015, 172, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Macêdo, D.G.; de Menezes, I.R.A.; Lacerda, S.R.; de Silva, M.A.P.; Ribeiro, D.A.; Macêdo, M.S.; Oliveira, L.G.S.; Saraiva, M.E.; Alencar, S.R.; Oliveira, S.F.; et al. Versatility and consensus of the use of medicinal plants in an area of cerrado in the Chapada do Araripe, Barbalha—CE-Brazil. J. Med. Plants Res. 2016, 10, 505–514. [Google Scholar] [CrossRef] [Green Version]
- Macedo, J.G.F.; de Menezes, I.R.A.; Ribeiro, D.A.; de Oliveira Santos, M.; de Macêdo, D.G.; Macedo, M.J.F.; Almeida, B.V.; de Oliveira, L.G.S.; Leite, C.P.; de Almeida Souza, M. Analysis of the variability of therapeutic indications of medicinal species in the northeast of Brazil: Comparative study. Evid. Based Complement. Altern. Med. 2018, 2018, 6769193. [Google Scholar] [CrossRef] [Green Version]
- Macêdo, M.J.F.; Ribeiro, D.A.; Santos, M.O.; Macêdo, D.G.; Macedo, J.G.F.; Almeida, B.V.; Saraiva, M.E.; Lacerda, M.N.S.; Souza, M.M.A. Fabaceae medicinal flora with therapeutic potential in Savanna areas in the Chapada do Araripe, Northeastern Brazil. Rev. Bras. Farmacogn. 2018, 28, 738–750. [Google Scholar] [CrossRef]
- Durão, H.L.G.; Costa, K.G.; Medeiros, M. Etnobotânica de plantas medicinais na comunidade quilombola de Porto Alegre, Cametá, Pará, Brasil. Boletim do Museu Paraense Emílio Goeldi. Ciências Nat. 2021, 16, 245–258. [Google Scholar] [CrossRef]
- Awika, J.M.; Rooney, L.W.; Wu, X.; Ronald, L.; Cisneros-Zevallos, L. Screening methods to measure antioxidant activity of sorghum (Sorghum bicolor) and sorghum products. J. Agric. Food Chem. 2003, 51, 6657–6662. [Google Scholar] [CrossRef]
- Costa, J.G.M.; Leite, G.O.; Dubois, A.F.; Seeger, R.P.; Boligon, A.A.; Athayde, M.L.; Campos, A.R.; Rocha, J.B.T. Antioxidant effect of Stryphnodendron rotundifolium Martius extracts from Cariri-Ceará state (Brazil): Potential involvement in its therapeutic use. Molecules 2012, 17, 934. [Google Scholar] [CrossRef]
- Salazar, G.J.T.; Dias, F.J.; Ribeiro, P.R.V.; de Brito, E.S.; Canuto, K.M.; Coutinho, H.D.M.; Ribeiro-Filho, J.; Gallo, M.; Montesano, D.; Naviglio, D.; et al. Antioxidant Activity of Stryphnodendron rotundifolium Mart. Stem Bark Fraction in an Iron Overload Model. Foods 2021, 10, 2683. [Google Scholar] [CrossRef]
- Aquino, M.D.A.; Teles, A.A.B.; Holanda, V.N.; Menezes, I.R.A.; Vandesmet, V.C.S. Evaluation of the Hepatic Activity of the Hydroalcolic Extract of Bark Stryphnodendron rotundifolium Mart. Rev. Interfaces 2017, 5, 48–54. [Google Scholar] [CrossRef]
- Barbosa, C.R. Uso do Barbatimão (Stryphnodendron rotundifolium) como Aditivo na Alimentação de Cordeiros em Confinamento. Master’s Thesis in Animal Science, Federal University of Grande Dourados, Faculty of Agrarian Sciences, Dourados, Brazil, 2021; p. 111. Available online: https://repositorio.ufgd.edu.br/jspui/bitstream/prefix/4587/1/CristianeRebou%c3%a7asBarbosa.pdf (accessed on 25 April 2022).
- Coutinho, H.D.M.; Martins, G.M.A.B.; Morais-Braga, M.F.B.; Menezes, I.R.A.; Machado, A.J.T.; Freitas, M.A.; Santos, A.T.L.; Costa, M.S.; Campina, F.F.; Silva, A.R.P.; et al. Stryphnodendron rotundifolium Mart. As an Adjuvant for the Plant Germination and Development under Toxic Concentrations of HgCl2 and AlCl3. Water Air Soil Pollut. 2017, 228, 424. [Google Scholar] [CrossRef]
- Funari, C.S.; Ferro, V.O. Análise de própolis. Food Sci. Technol. 2006, 26, 171–178. Available online: https://www.scielo.br/j/cta/a/YRSX8qYyTD3dcgjnDyq6VwK/?format=pdf&lang=pt (accessed on 21 October 2021). [CrossRef] [Green Version]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying in the improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Benzie, I.; Strain, J. Ferric reducing/antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1999; Volume 299, pp. 15–27. [Google Scholar] [CrossRef]
- Antolovich, M.; Prenzler, P.; Patsalides, E.; Mc Donald, S.; Robards, K. Methods for testing antioxidant activity. Analyst 2002, 127, 183–198. [Google Scholar] [CrossRef]
- Choi, J.; Hee, J.L.; Sam, S.K. Alatemin, Cassiaside and Rubrofusarin gentiobioside, Radical Scavenging Principles from the Seeds of Cassia tora on 1,1-Diphenyl-2-picrylhydrazyl (DPPH) Radical. Arch. Pharmacal Res. 1994, 17, 462–466. [Google Scholar] [CrossRef]
- Prieto, P.; Pineda, M.; Aguilar, M. Spectrophotometric quantitation of antioxidant capacity through the formation of a phosphomolybdenum complex. Specific application to the determination of vitamin E. Anal. Biochem. 1999, 269, 337–341. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Duavy, S.M.; Ecker, A.; Salazar, G.T.; Loreto, J.; Costa, J.G.M.; Barbosa, N.V. Pequi enriched diets protect Drosophila melanogaster against paraquat-induced locomotor deficits and oxidative stress. J. Toxicol. Environ. Health 2019, 82, 664–677. [Google Scholar] [CrossRef]
- Feany, M.B.; Bender, W.W. A Drosophila model of Parkinson’s disease. Nature 2000, 404, 394–398. [Google Scholar] [CrossRef]
- White, K.E.; Humphrey, D.M.; Hirth, F. The dopaminergic system in the aging brain of Drosophila. Front. Neurosci. 2010, 4, 205. [Google Scholar] [CrossRef] [Green Version]
- Abolaji, A.O.; Kamdem, J.P.; Lugokenski, T.H.; Nascimento, T.K.; Waczuk, E.P.; Farombi, E.O.; Loreto, É.L.S.; Rocha, J.B.T. Involvement of oxidative stress in 4-vinylcyclohexene-inducid toxicity in Drosophila melanogaster. Free. Radic. Biol. Med. 2014, 71, 99–108. [Google Scholar] [CrossRef]
- Coulom, H.; Birman, S. Chronic exposure to rotenone models sporadic Parkinson’s disease in Drosophila melanogaster. J. Neurosci. 2004, 24, 10993–10998. [Google Scholar] [CrossRef] [Green Version]
- Heijnen, C.G.; Haenen, G.R.; Oostveen, R.M.; Stalpers, E.M.; Bast, A. Protection of flavonoids against lipid peroxidation: The structure activity relationship revisited. Free Rad. Res. 2002, 36, 575–581. [Google Scholar] [CrossRef] [Green Version]
- Rice-Evans, C.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Antioxidant properties of phenolic compounds. Trends Plant Sci. 1997, 2, 152–159. [Google Scholar] [CrossRef]
- Lopes, G.K.; Schulman, H.M.; Hermes-Lima, M. Polyphenol tannic acid inhibits hidroxil radical formation from Fenton reaction by complexing ferrous ions. Biochim. Biophys. Acta Gen. Subj. 1999, 1472, 142–152. [Google Scholar] [CrossRef]
- Pannala, A.S.; Chan, T.S.; O’Brien, P.J.; Rice-Evans, C.A. Flavonoid B-Ring chemistry and antioxidant activity: Fast Reaction Kinetics. Biochem. Biophys. Res. Commun. 2001, 282, 1161–1168. [Google Scholar] [CrossRef]
- Macáková, K.; Kolečkář, V.; Cahlíková, L.; Chlebek, J.; Hošt’álková, A.; Kuča, K.; Jun, D.; Opletal, L. Chapter 6: Tannins and their Influence on Health. In Recent Advances in Medicinal Chemistry, 1st ed.; Choudhary, M.I., Perry, G., Rahman, A., Eds.; Elsevier: San Antonio, TX, USA; Bentham Science Publishers: Soest, The Netherlands, 2014; Volume 1, pp. 159–208. [Google Scholar] [CrossRef]
- Baldivia, D.D.S.; Leite, D.F.; Castro, D.T.H.; Campos, J.F.; Santos, U.P.D.; Paredes-Gamero, E.J.; Carollo, C.A.; Silva, D.B.; Picoli, S.K.; Santos, E.L. Evaluation of In vitro Antioxidant and Anticancer Properties of the Aqueous Extract from the Stem Bark of Stryphnodendron adstringens. Int. J. Mol. Sci. 2018, 19, 2432. [Google Scholar] [CrossRef] [Green Version]
- Zocoler, D.A.M.; Sanches, C.A.C.; Albrecht, I.; Mello, P.J.C. Antioxidant capacity of extracts and isolated compounds from Stryphnodendron obovatum Benth. Braz. J. Pharm. Sci. 2009, 45, 443–452. [Google Scholar] [CrossRef]
- Lu, Y.; Yeap, F.L. Antioxidant and radical scavenging activities of polyphenols from apple pomace. Food Chem. 2000, 68, 81–85. [Google Scholar] [CrossRef]
- Halliwell, B.; Aeschbach, R.; Loliger, J.; Arouma, O.I. The characterization of antioxidants. Food Chem. Toxicol. 1995, 33, 601–617. [Google Scholar] [CrossRef]
- Olivieri, N.F. The beta-thalassemias. N. Engl. J. Med. 1999, 341, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Steensma, D.P.; Tefferi, A. The myelodysplastic syndrome(s): A perspective and review highlighting current controversies. Leuk. Res. 2003, 27, 95–120. [Google Scholar] [CrossRef]
- Costa, A.F.; Faulhaber, G.A.M.; Krug, B.C.; Gonçalves, C.B.T.; Amaral, K.M.; Schneiders, R.E.; Xavier, L.C.; Mosca, M. Protocolo Clínico e Diretrizes Terapêuticas: Sobrecarga de Ferro. Portaria SAS/MS No. 1324. Brazil documentos. 2013. Available online: https://bvsms.saude.gov.br/bvs/saudelegis/sas/2013/prt1324_25_11_2013.html (accessed on 22 April 2022).
- Nichols, C.D. Drosophila melanogaster neurobiology, neuropharmacology, and how the fly can inform central nervous system drug discovery. Pharmacol. Ther. 2006, 112, 677–700. [Google Scholar] [CrossRef]
- Fernández, J.R.; Grant, M.D.; Tulli, N.M.; Karkowski, L.M.; McClearn, G.E. Differences in locomotor activity across the lifespan of Drosophila melanogaster. Exp. Gerontol. 1999, 34, 621–631. [Google Scholar] [CrossRef]
- Gargano, J.W.; Martin, I.; Bhandari, P.; Grotewiel, M.S. Rapid iterative negative geotaxis (RING): A new method for assessing age-related locomotor decline in Drosophila. Exp. Gerontol. 2005, 40, 386–395. [Google Scholar] [CrossRef]
- Besson, M.; Martin, J.-R. Centrophobism/thigmotaxis, a new role for the mushroom bodies in Drosophila. J. Neurobiol. 2004, 62, 386–396. [Google Scholar] [CrossRef]
- Lebreton, S.; Martin, J.R. Mutations affecting the cAMP transduction pathway disrupt the centrophobism behavior. J. Neurogenet. 2009, 23, 225–234. [Google Scholar] [CrossRef]
- Zhao, X.; Sun, H.; Hou, A.; Zhao, Q.; Wei, T.; Xin, W. Antioxidant properties of two gallotannins isolated from the leaves of Pistacia weinmannifolia. Biochim. Biophys. Acta 2005, 1725, 103–110. [Google Scholar] [CrossRef]
- Ou, K.; Gu, L. Absorption and metabolism of proanthocyanidins. J. Funct. Foods 2014, 7, 43–53. [Google Scholar] [CrossRef]
- Kruszewski, M. Labile iron pool: The main determinant of cellular response to oxidative stress. Mutat. Res. Mol. Mech. Mutagen. 2003, 531, 81–92. [Google Scholar] [CrossRef]
- Liu, Z.D.; Hider, R.C. Design of iron chelators with therapeutic application. Coord. Chem. Rev. 2002, 232, 151–171. [Google Scholar] [CrossRef]
- Birch, N.; Wang, X.; Chong, H.S. Iron chelators as therapeutic iron depletion agents. Expert Opin. Ther. Patents 2006, 16, 1533–1556. [Google Scholar] [CrossRef]
- Choudhry, V.P.; Naithani, R. Current status of iron overload and chelation with deferasirox. Indian J. Pediatr. 2007, 8, 759–764. [Google Scholar] [CrossRef]
- Vassilis, L.; Helen, B.; Efstathios, G.; Antonis, K. Deferasirox administration for the treatment of non-transfusional iron overload in patients with thalassaemia intermedia. Br. J. Haematol. 2010, 151, 504–508. [Google Scholar] [CrossRef]
- Santini, V.; Alessandrino, P.E.; Angelucci, E.; Barosi, G.; Billio, A.; Di Maio, M.; Finelli, C.; Locatelli, F.; Marchetti, M.; Morra, E.; et al. Clinical Management of Myelodysplastic Syndromes: Update of SIE, SIES, GITMO Practice Guidelines. Leuk. Res. 2010, 34, 1576–1588. [Google Scholar] [CrossRef]
- Musto, P.; Maurillo, L.; Simeon, V.; Poloni, A.; Finelli, C.; Balleari, E.; Ricco, A.; Rivellini, F.; Cortelezzi, A.; Tarantini, G.; et al. Iron-Chelating Therapy with Deferasirox in Transfusion-Dependent, Higher Risk Myelodysplastic Syndromes: A Retrospective, Multicenter Study. Br. J. Haematol. 2017, 177, 741–750. [Google Scholar] [CrossRef]
- Palumbo, G.A.; Galimberti, S.; Barcellini, W.; Cilloni, D.; Di Renzo, N.; Elli, E.M.; Finelli, C.; Maurillo, L.; Ricco, A.; Musto, P.; et al. From Biology to Clinical Practice: Iron Chelation Therapy with Deferasirox. Front. Oncol. 2021, 11, 752192. [Google Scholar] [CrossRef]
- Di Veroli, A.; Campagna, A.; De Muro, M.; Trawinska, M.M.; Leonetti, C.S.; Petriccione, L.; Romano, A.; D’Addosio, A.; Cenfra, A.; Montanaro, M.; et al. Deferasirox in the treatment of iron overload during myeloproliferative neoplasms in fibrotic phase: Does ferritin decrement matter? Leuk. Res. 2018, 76, 65–69. [Google Scholar] [CrossRef]
- Current and Resolved Drug Shortages and Discontinuations Reported to FDA, and Drugs@FDA: FDA-Approved Drugs. Available online: www.accessdata.fda.gov (accessed on 25 May 2021).
- de Toledo Espindola, P.P.; dos Santos da Rocha, P.; Carollo, C.A.; Schmitz, W.O.; Pereira, Z.V.; do Carmo Vieira, M.; Santos, E.L.; de Picoli Souza, K. Antioxidant and antihyperlipidemic effects of Campomanesia adamantium O. Berg root. Oxidative Med. Cell. Longev. 2016, 2016, 7910340. [Google Scholar] [CrossRef] [Green Version]
- Lopes, R.H.; Macorini, L.F.; Antunes, K.Á.; Espindola, P.T.; Alfredo, T.M.; da Rocha, P.S.; Pereira, Z.V.; Santos, E.L.; Souza, K.P. Antioxidant and hypolipidemic activity of the hydroethanolic extract of Curatella americana l. leaves. Oxidative Med. Cell. Longev. 2016, 2016, 9681425. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ABTS•+ | FRAP | DPPH● | ABTS•+ | FRAP | DPPH● | |
|---|---|---|---|---|---|---|
| (IC50 ± SEM) μg/mL | (Max. Act. ± SEM)% | |||||
| TFSR | 0.74 ± 0.12 | 3.27 ± 0.13 | 8.65 ± 1.10 | 95.30 ± 0.60 | 74.7 ± 0.2 (5.0 μg/mL) | 82.4 ± 3.2 |
| Quercetin | 0.47 ± 0.03 | 0.90 ± 0.04 | - | 99.73 ± 0.03 | 75.6 ± 0.1 (1.5 μg/mL) | - |
| Rutin | 1.68 ± 0.27 | - | - | 80.41 ± 0.05 | - | - |
| Gallic acid | 1.02 ± 0.14 | - | - | 93.0 ± 0.1 (1.75 μg/mL) | - | |
| Catechin | 1.62 ± 0.10 | - | - | 87.5 ± 0.1 (2.75 μg/mL) | - | |
| Ascorbic acid | 1.95 ± 0.07 | 82.2 ± 3.2 | ||||
| TFSRAA | TFSRRutin | TFSRCatechin | |
|---|---|---|---|
| IC50 (mg/mL) | 0.0111 ± 0.0014 | 0.01201 ± 0.0013 | 0.01296 ± 0.0014 |
| Maximum activity (%) | 80.62 ± 2.63 | 76.40 ± 2.42 | 71.62 ± 2.81 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salazar, G.J.T.; Ecker, A.; Adefegha, S.A.; da Costa, J.G.M. Advances in Evaluation of Antioxidant and Toxicological Properties of Stryphnodendron rotundifolium Mart. in Drosophila melanogaster Model. Foods 2022, 11, 2236. https://doi.org/10.3390/foods11152236
Salazar GJT, Ecker A, Adefegha SA, da Costa JGM. Advances in Evaluation of Antioxidant and Toxicological Properties of Stryphnodendron rotundifolium Mart. in Drosophila melanogaster Model. Foods. 2022; 11(15):2236. https://doi.org/10.3390/foods11152236
Chicago/Turabian StyleSalazar, Gerson Javier Torres, Assis Ecker, Stephen Adeniyi Adefegha, and José Galberto Martins da Costa. 2022. "Advances in Evaluation of Antioxidant and Toxicological Properties of Stryphnodendron rotundifolium Mart. in Drosophila melanogaster Model" Foods 11, no. 15: 2236. https://doi.org/10.3390/foods11152236
APA StyleSalazar, G. J. T., Ecker, A., Adefegha, S. A., & da Costa, J. G. M. (2022). Advances in Evaluation of Antioxidant and Toxicological Properties of Stryphnodendron rotundifolium Mart. in Drosophila melanogaster Model. Foods, 11(15), 2236. https://doi.org/10.3390/foods11152236