

Inhibition of Chitosan with Different Molecular Weights on Barley-Borne Fusarium graminearum during Barley Malting Process for Improving Malt Quality


Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms and Materials
2.2. Determination of Minimum Inhibition Concentration (MIC)
2.3. Barley Grain Surface Disinfection
2.4. Mold Spore Suspension Preparation and Grain Inoculation
2.5. Barley Malting Process
2.6. Influence of Chitosan on Fusarium graminearum Infection during Malting
2.7. Analysis of the Infection Rate of Fusarium graminearum
2.8. Quantification of Fusarium graminearum Growth by PCR and Photometric Assay
2.9. Analysis of Deoxynivalenol (DON) Content
2.10. Analysis of Barley Germination Performances and Malt Quality Parameters
2.11. Statistical Analysis
3. Results and Discussion
3.1. Inhibitory Effects of Chitosan against Fusarium graminearum
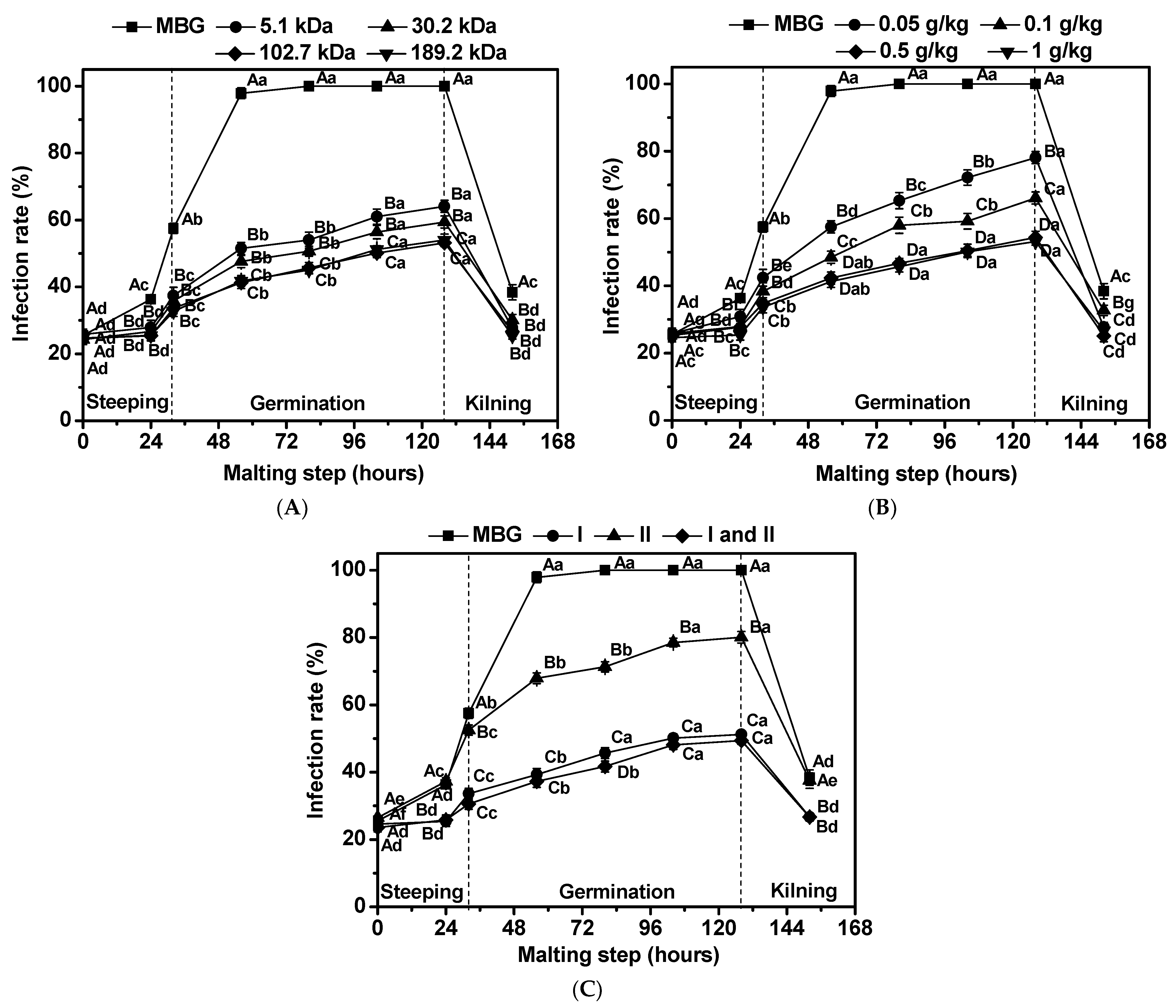
3.2. Effects of Chitosan on Infection Rate of Fusarium graminearum in Barley
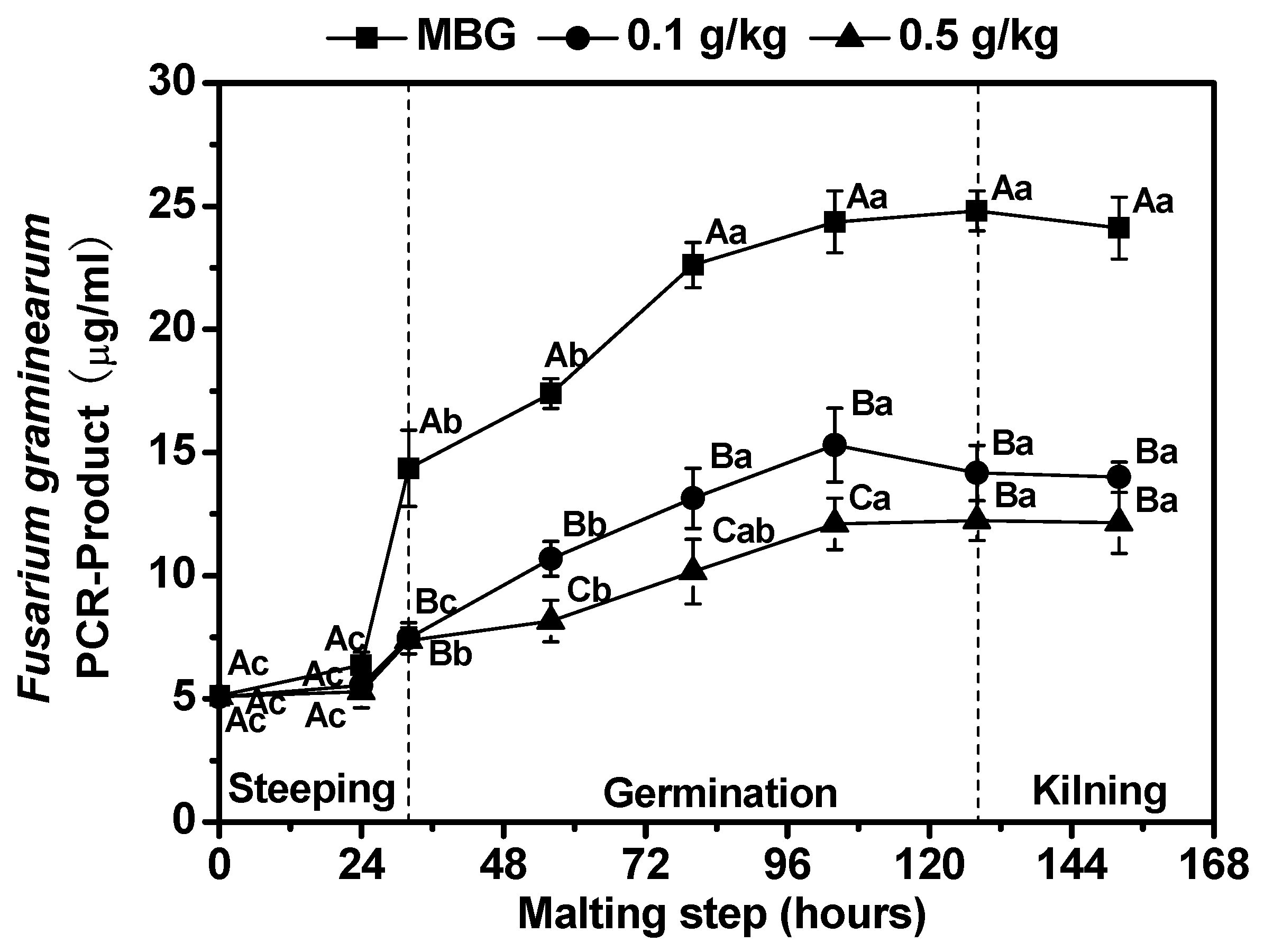
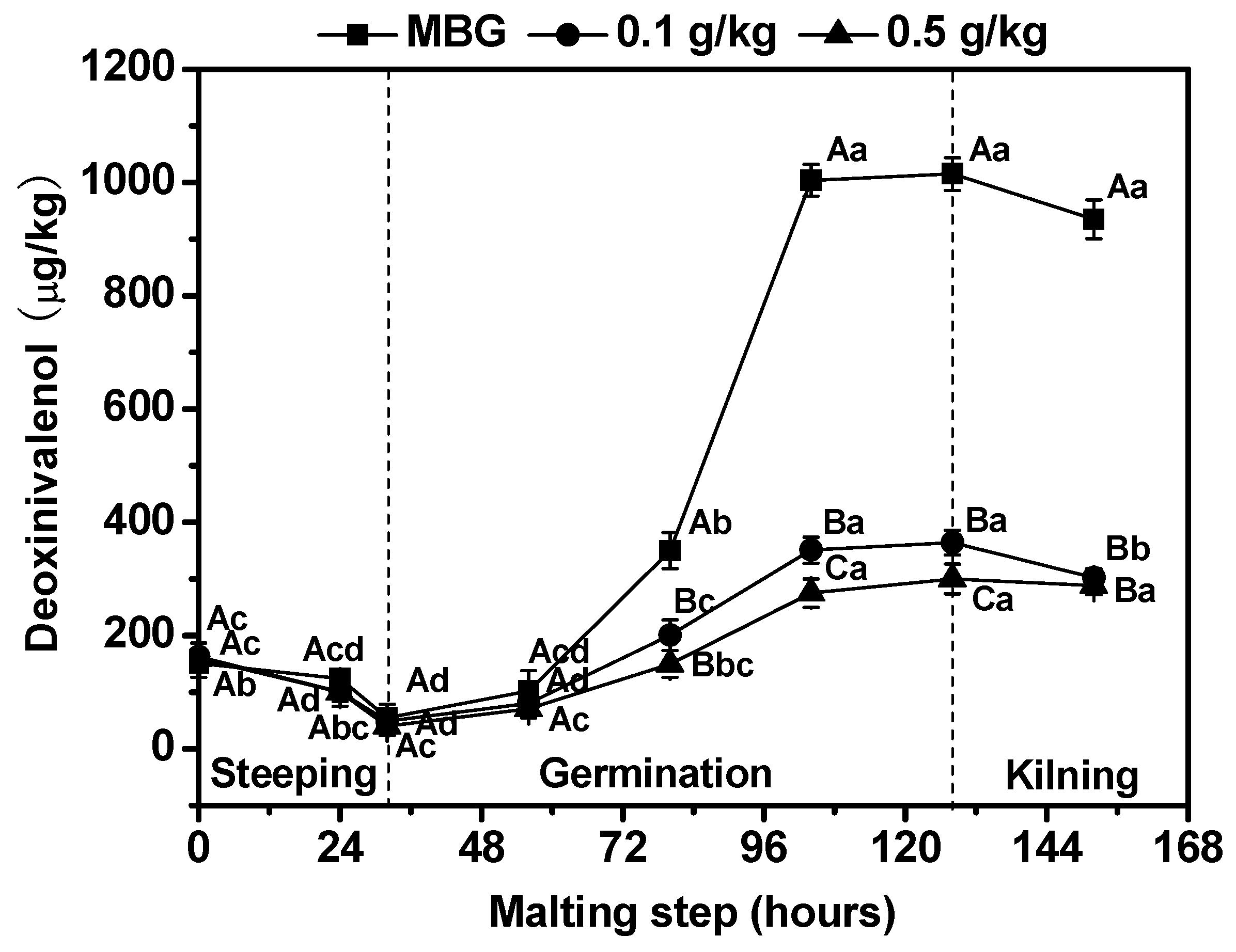
3.3. Effects of Chitosan on Fusarium graminearum Biomass and Deoxynivalenol (DON) Content in Barley
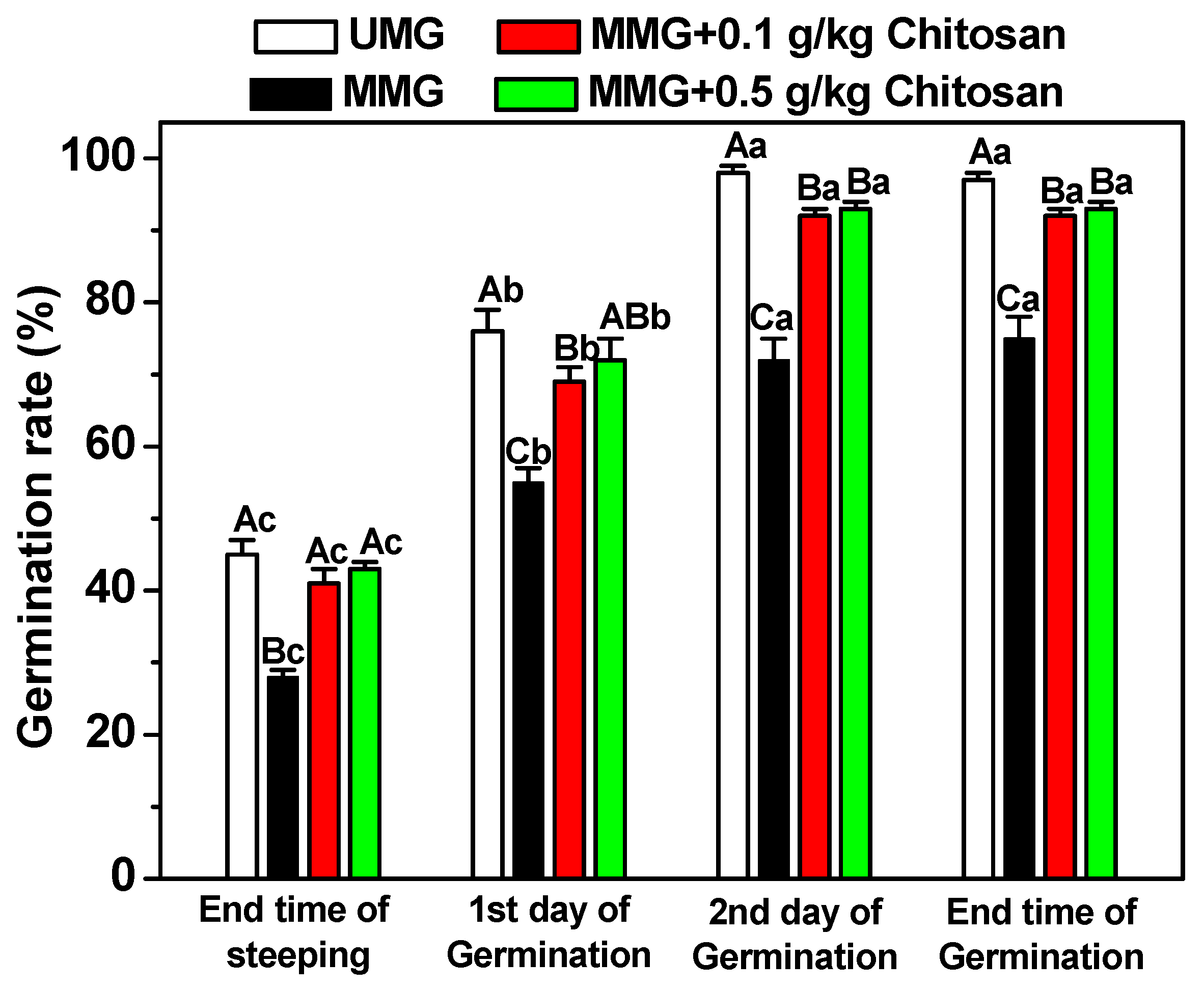
3.4. Effects of Chitosan on Germination Ability of Fusarium graminearum-Infected Barley
3.5. Effects of Chitosan on Quality Performance of Fusarium graminearum-Infected Malt
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pavel, J.; Vaculová, K.; Faltusová, Z.; Kučera, L.; Sedláčková, I.; Tvaružek, L.; Ovesná, J. Effect of fungicide treatment on Fusarium culmorum and Tri genes transcription in barley malt. Czech J. Food Sci. 2015, 33, 326–333. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, P.M.; Brosnan, B.; Furey, A.; Coffey, A.; Zannini, E. Lactic acid bacteria bioprotection applied to the malting process. Part II: Strain characterization and identification of antifungal compounds. Food Control 2015, 51, 433–443. [Google Scholar] [CrossRef]
- Steiner, E.; Gastl, M.; Becker, T. Protein changes during malting and brewing with focus on haze and foam formation: A review. Eur. Food Res. Technol. 2011, 232, 191–204. [Google Scholar] [CrossRef]
- Laitila, A.; Kotaviita, E.; Peltola, P.; Home, S.; Wilhelmson, A. Indigenous microbial community of barley greatly influences grain germination and malt quality. J. Inst. Brew. 2007, 113, 9–20. [Google Scholar] [CrossRef]
- Laitila, A.; Sarlin, T.; Kotaviita, E.; Huttunen, T.; Home, S.; Wilhelmson, A. Yeasts isolated from industrial malting can suppress Fusarium growth and formation of gushing factors. J. Ind. Microbiol.Biot. 2007, 34, 701–713. [Google Scholar] [CrossRef]
- Kaur, M.; Bowman, J.P.; Stewart, D.C.; Evans, D.E. The fungal community structure of barley malts from diverse geographical regions correlates with malt quality parameters. Int. J. Food Microbiol. 2015, 215, 71–78. [Google Scholar] [CrossRef]
- Justé, A.; Malfliet, S.; Lenaerts, L.; De Cooman, L.; Aerts, G.; Willems, K.A.; Lievens, B. Microflora during malting of barley: Overview and impact on malt quality. Brew. Sci. 2011, 64, 22–30. [Google Scholar]
- Oliveira, P.M.; Mauch, A.; Jacob, F.; Waters, D.M.; Arendt, E.K. Fundamental study on the influence of Fusarium infection on quality and ultrastructure of barley malt. Int. J. Food Microbiol. 2012, 156, 32–43. [Google Scholar] [CrossRef]
- Kumar, K.N.; Venkataramana, M.; Allen, J.A.; Chandranayaka, S.; Murali, H.S.; Batra, H.V. Role of Curcuma longa L. essential oil in controlling the growth andzearalenone production of Fusarium graminearum. LWT-Food Sci. Technol. 2016, 69, 522–528. [Google Scholar] [CrossRef]
- Wolf-Hall, C.E. Mold and mycotoxin problems encountered during malting and brewing. Int. J. Food Microbiol. 2007, 119, 89–94. [Google Scholar] [CrossRef]
- Chen, D.; Chen, P.; Cheng, Y.; Peng, P.; Liu, J.; Ma, Y.; Liu, Y.; Ruan, R. Deoxynivalenol decontamination in raw and germinating barley treated by plasma-activated water and intense pulsed light. Food Bioprocess Technol. 2019, 12, 246–254. [Google Scholar] [CrossRef]
- Peng, Z.; Feng, W.; Cai, G.; Wu, D.; Lu, J. Enhancement effect of chitosan coating on inhibition of deoxynivalenol accumulation by Litsea cubeba essential oil emulsion during malting. Foods 2021, 10, 3051. [Google Scholar] [CrossRef] [PubMed]
- Xing, K.; Zhu, X.; Peng, X.; Qin, S. Chitosan antimicrobial and eliciting properties for pest control in agriculture: A review. Agron. Sustain. Dev. 2015, 35, 569–588. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yin, L.; Rasheed, H.A.; Xia, X.; Tekliye, M.; Li, Z.; Li, J.; Dong, M. Effects of chitosan on the physicochemical properties, in vitro starch digestibility, antimicrobial potentials, and antioxidant activities of purple highland barley noodles. LWT-Food Sci. Technol. 2020, 132, 109802–109807. [Google Scholar] [CrossRef]
- Hsu, S.H.; Chang, Y.B.; Tsai, C.L.; Fu, K.Y.; Wang, S.H.; Tseng, H.J. Characterization and biocompatibility of chitosan nanocomposites. Colloid. Surface B 2011, 85, 198–206. [Google Scholar] [CrossRef]
- Kim, H.; Tator, C.H.; Shoichet, M.S. Chitosan implants in the rat spinal cord: Biocompatibility and biodegradation. J. Biomed. Mater. Res. A 2011, 97, 395–404. [Google Scholar] [CrossRef]
- Rabea, E.I.; Badawy, M.E.I.; Steurbaut, W.; Stevens, C.V. In vitro assessment of N-(benzyl) chitosan derivatives against some plant pathogenic bacteria and fungi. Eur. Polym. J. 2009, 45, 237–245. [Google Scholar] [CrossRef]
- Li, S.J.; Zhu, T.H. Biochemical response and induced resistance against anthracnose (Colletortrichumcamelliae) of camellia (Camellia pitardii) by chitosan oligosaccharide application. Forest Pathol. 2013, 43, 67–76. [Google Scholar] [CrossRef]
- Mansilla, A.Y.; Albertengo, L.; Rodríguez, M.S.; Debbaudt, A.; Zúňiga, A.; Casalongué, C.A. Evidence on antimicrobial properties and mode of action of a chitosan obtained from crustacean exoskeletons on Pseudomonas syringae pv. tomato DC3000. App. Microbiol. Biot. 2013, 97, 6957–6966. [Google Scholar] [CrossRef]
- Al-Hetar, M.Y.; Abidin, M.A.Z.; Sariah, M.; Wong, M.Y. Antifungal activity of chitosan against Fusarium oxysporum f. sp. cubense. J. Appl. Polym. Sci. 2011, 120, 2434–2439. [Google Scholar] [CrossRef]
- De Rodríguez, D.J.; Trejo-González, F.A.; Rodríguez-García, R.; Díaz-Jimenez, M.L.V.; Sáenz-Galindo, A.; Hernández-Castillo, F.D.; Villarreal-Quintanilla, J.A.; Peňa-Ramos, F.M. Antifungal activity in vitro of Rhus muelleri against Fusarium oxysporum f. sp. lycopersici. Ind. Crops Prod. 2015, 75, 150–158. [Google Scholar] [CrossRef]
- Im, H.; Kim, K.M.; Lee, S.; Ryu, C. Functional metagenome mining of soil for a novel gentamicin resistance gene. J. Molecul. Microb. Biotech. 2016, 26, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Mauch, A.; Dal Bello, F.; Coffey, A.; Arendt, E.K. The use of Lactobacillus brevis PS1 to in vitro inhibit the outgrowth of Fusarium culmorum and other common Fusarium species found on barley. Int. J. Food Microbiol. 2010, 141, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Vegi, A.; Schwarz, P.; Wolf-Hall, C.E. Quantification of Tri5 gene, expression, and deoxynivalenol production during the malting of barley. Int. J. Food Microbiol. 2011, 150, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, P.M.; Brosnan, B.; Jacob, F.; Coffey, A.; Zannini, E.; Arendt, E.K. Lactic acid bacteria bioprotection applied to the malting process. Part I: Substrate impact and mycotoxin reduction. Food Control 2015, 51, 444–452. [Google Scholar] [CrossRef]
- Boenisch, M.J.; Schäfer, W. Fusarium graminearum forms mycotoxin producting infection structures on wheat. BMC Plant Biol. 2011, 11, 110. [Google Scholar] [CrossRef] [Green Version]
- Lan, W.; Wang, W.; Yu, Z.; Qin, Y.; Luan, J.; Li, X. Enhanced germination of barley (Hordeum vulgare L.) using chitooligosaccharide as an elicitor in seed priming to improve malt quality. Biotechnol. Lett. 2016, 38, 1935–1940. [Google Scholar] [CrossRef]
- American Society of Brewing Chemists (ASBC). ASBC Analysis Method, 14th ed.; ASBC Press: Saint Paul, MN, USA, 2008; pp. 25–55. [Google Scholar]
- European Brewery Convention (EBC). Analytica-EBC, 3rd ed.; Verlag Hans Getränke-Fachverlag Press: Nürnberg, Germany, 2004; pp. 59–72. [Google Scholar]
- Hope, R.; Magan, N. Two-dimensional environmental profiles of growth, deoxynivalenol and nivalenol production by Fusarium culmorum on a wheat-based substrate. Lett. Appl. Microbiol. 2003, 37, 70–74. [Google Scholar] [CrossRef]
- Dananjaya, S.H.S.; Erandani, W.K.C.U.; Kim, C.; Nikapitiya, C.; Lee, J.; De Zoysa, M. Comparative study on antifungal activities of chitosan nanoparticles and chitosan silver nano composites against Fusarium oxysporum species complex. Int. J. Biol.Macromol. 2017, 105, 478–488. [Google Scholar] [CrossRef]
- Kheiri, A.; Moosawijorf, S.A.; Mallihipour, A.; Saremi, H.; Nikkhah, M. Application of chitosan and chitosan nanoparticles for the control of Fusarium head blight of wheat (Fusarium graminearum) in vitro and greenhouse. Int. J. Biol.Macromol. 2016, 93, 1261–1272. [Google Scholar] [CrossRef]
- Khan, M.R.; Doohan, F.M. Comparison of the efficacy of chitosan with that of a fluorescent pseudomonad for the control of Fusarium head blight disease of cereals and associated mycotoxin contamination of grain. Biol. Control 2009, 48, 48–54. [Google Scholar] [CrossRef]
- Li, X.F.; Feng, X.Q.; Yang, S.; Wang, T.P.; Su, Z.X. Effects of molecular weight and concentration of chitosan on antifungal activity against Aspergillus Niger. Iran. J. Sci.Technol. 2008, 17, 843–852. [Google Scholar]
- Tsai, G.J.; Su, W.H.; Chen, H.S.; Pan, C.L. Antimicrobial activity of shrimp chitin and chitosan from different treatments and applications of fish preservation. Fish. Sci. 2002, 68, 170–177. [Google Scholar] [CrossRef] [Green Version]
- Park, S.I.; Daeschel, M.A.; Zhao, Y. Functional properties of antimicrobial lysozyme-chitosan composite films. J. Food Sci. 2004, 69, 215–221. [Google Scholar] [CrossRef]
- Zhang, L.Y.; Zhu, J.F. Study on antimicrobial activity of chitosan with different molecular weights. Carbohyd. Polym. 2003, 54, 527–530. [Google Scholar] [CrossRef]
- Hernández-Lauzardo, A.N.; Bautista-Baňos, S.; Velázquez-del Valle, M.G.; Méndez-Montealvo, M.G.; Sánchez-Rivera, M.M.; Bello-Pérez, L.A. Antifungal effects of chitosan with different molecular weights on in vitro development of Rhizopus stolonifer (Ehrenb.:Fr.) Vuill. Carbohyd. Polym. 2008, 73, 541–547. [Google Scholar] [CrossRef]
- Younes, I.; Sellimi, S.; Rinaudo, M.; Jellouli, K.; Nasri, M.N. Influence of acetylation degree and molecular weight of homogeneous chitosans on antibacterial and antifungal activities. Int. J. Food Microbiol. 2014, 185, 57–63. [Google Scholar] [CrossRef]
- Liu, J.; Tian, S.; Meng, X.; Xu, Y. Effects of chitosan on control of postharvest diseases and physiological responses of tomato fruit. Postharvest Biol. Tec. 2007, 44, 300–306. [Google Scholar] [CrossRef]
- Yu, J.; Yin, H.; Dong, J.; Zhang, C.; Zhang, B.; Jin, Z.; Cao, Y. Pullulation of toxigenic Fusarium and deoxynivalenol in the malting of de minimis infected barley (Hordeum vulgare). LWT-Food Sci. Technol. 2019, 113, 108242–108262. [Google Scholar] [CrossRef]
- Reddy, M.V.B.; Arul, J.; Angers, P.; Couture, L. Chitosan treatment of wheat seeds induces resistance to Fusarium graminearum and improves seeds quality. J. Agr. Food Chem. 1999, 47, 67–72. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chitosan (k Da) | Mycelial Growth | Spore Germination |
|---|---|---|
| 5.1 | 12.8 ± 0.4 a | 3.6 ± 0.2 c |
| 30.2 | 4.6 ± 0.1 b | 2.0 ± 0.1 f |
| 102.7 | 3.1 ± 0.2 d | 1.6 ± 0.1 g |
| 189.3 | 2.6 ± 0.1 e | 1.6 ± 0.1 g |
| Parameters | UMG | MMG | MMG + 0.1 g/kg Chitosan | MMG + 0.5 g/kg Chitosan |
|---|---|---|---|---|
| Malt water (%) | 4.5 ± 0.2 a | 4.3 ± 0.2 a | 4.4 ± 0.1 a | 4.4 ± 0.3 a |
| Brittleness (%) | 89.5 ± 0.5 a | 67.5 ± 0.4 b | 86.1 ± 0.5 a | 85.2 ± 0.4 a |
| Kolbach Index (%) | 42 ± 1 a | 38 ± 1 b | 42 ± 2 a | 43 ± 2 a |
| α-amino nitrogen (%) | 95 ± 3 b | 82 ± 2 c | 110 ± 4 a | 100 ± 4 b |
| Extract (%) | 77.5 ± 0.3 a | 54.3 ± 0.2 b | 77.8 ± 0.4 a | 76.8 ± 0.6 a |
| Diastatic power (oWK) | 198 ± 5 a | 165 ± 4 b | 204 ± 4 a | 198 ± 4 a |
| Colority (EBC) | 3.5 ± 0.0 b | 4.0 ± 0.0 a | 3.5 ± 0.0 b | 3.5 ± 0.0 b |
| Turbidity (EBC) | 0.4 ± 0.1 b | 1.5 ± 0.2 a | 0.5 ± 0.1 b | 0.6± 0.1 b |
| β-glucan content (mg/L) | 79 ± 3 b | 99 ± 3 a | 70 ± 2 c | 73 ± 2 c |
| Viscosity (mPa·s) | 1.4 ± 0.1 ab | 1.6 ± 0.1 a | 1.4 ± 0.1 ab | 1.4 ± 0.0 b |
| Glucose content (g/L) | 9.3 ± 0.2 a | 8.1 ± 0.1 b | 9.2 ± 0.2 a | 9.2 ± 0.1 a |
| Maltose content (g/L) | 45.6 ± 2.1 c | 35.4 ± 2.4 d | 54.8 ± 1.5 a | 50.2 ± 1.3 b |
| Maltotriose content (g/L) | 8.0 ± 0.2 a | 6.1 ± 0.1 b | 8.0 ± 0.3 a | 8.1 ± 0.5 a |
| pH | 6.0 ± 0.1 a | 5.8 ± 0.1 a | 5.9 ± 0.1 a | 6.0 ± 0.1 a |
| Total acid (mL/100 mL) | 1.4 ± 0.1 a | 1.5 ± 0.0 a | 1.5 ± 0.0 a | 1.4 ± 0.1 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luan, J.; Wei, X.; Li, Z.; Tang, W.; Yang, F.; Yu, Z.; Li, X. Inhibition of Chitosan with Different Molecular Weights on Barley-Borne Fusarium graminearum during Barley Malting Process for Improving Malt Quality. Foods 2022, 11, 3058. https://doi.org/10.3390/foods11193058
Luan J, Wei X, Li Z, Tang W, Yang F, Yu Z, Li X. Inhibition of Chitosan with Different Molecular Weights on Barley-Borne Fusarium graminearum during Barley Malting Process for Improving Malt Quality. Foods. 2022; 11(19):3058. https://doi.org/10.3390/foods11193058
Chicago/Turabian StyleLuan, Jing, Xu Wei, Zhefeng Li, Wenzhu Tang, Fan Yang, Zhimin Yu, and Xianzhen Li. 2022. "Inhibition of Chitosan with Different Molecular Weights on Barley-Borne Fusarium graminearum during Barley Malting Process for Improving Malt Quality" Foods 11, no. 19: 3058. https://doi.org/10.3390/foods11193058
APA StyleLuan, J., Wei, X., Li, Z., Tang, W., Yang, F., Yu, Z., & Li, X. (2022). Inhibition of Chitosan with Different Molecular Weights on Barley-Borne Fusarium graminearum during Barley Malting Process for Improving Malt Quality. Foods, 11(19), 3058. https://doi.org/10.3390/foods11193058