


Comparative Study on Grape Berry Anthocyanins of Various Teinturier Varieties

 , , ,
, , ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Experimental Site and Plant Materials
2.3. Genetic Fingerprinting and Determination of the MybA1t Repeat Number at the Berry Color Locus (BCL)
2.4. Analysis of Organic Acids
2.5. Extraction of Berries’ Skin
2.6. Preparation of Juice Samples
2.7. Analysis of Anthocyanins
2.8. Statistical Analysis
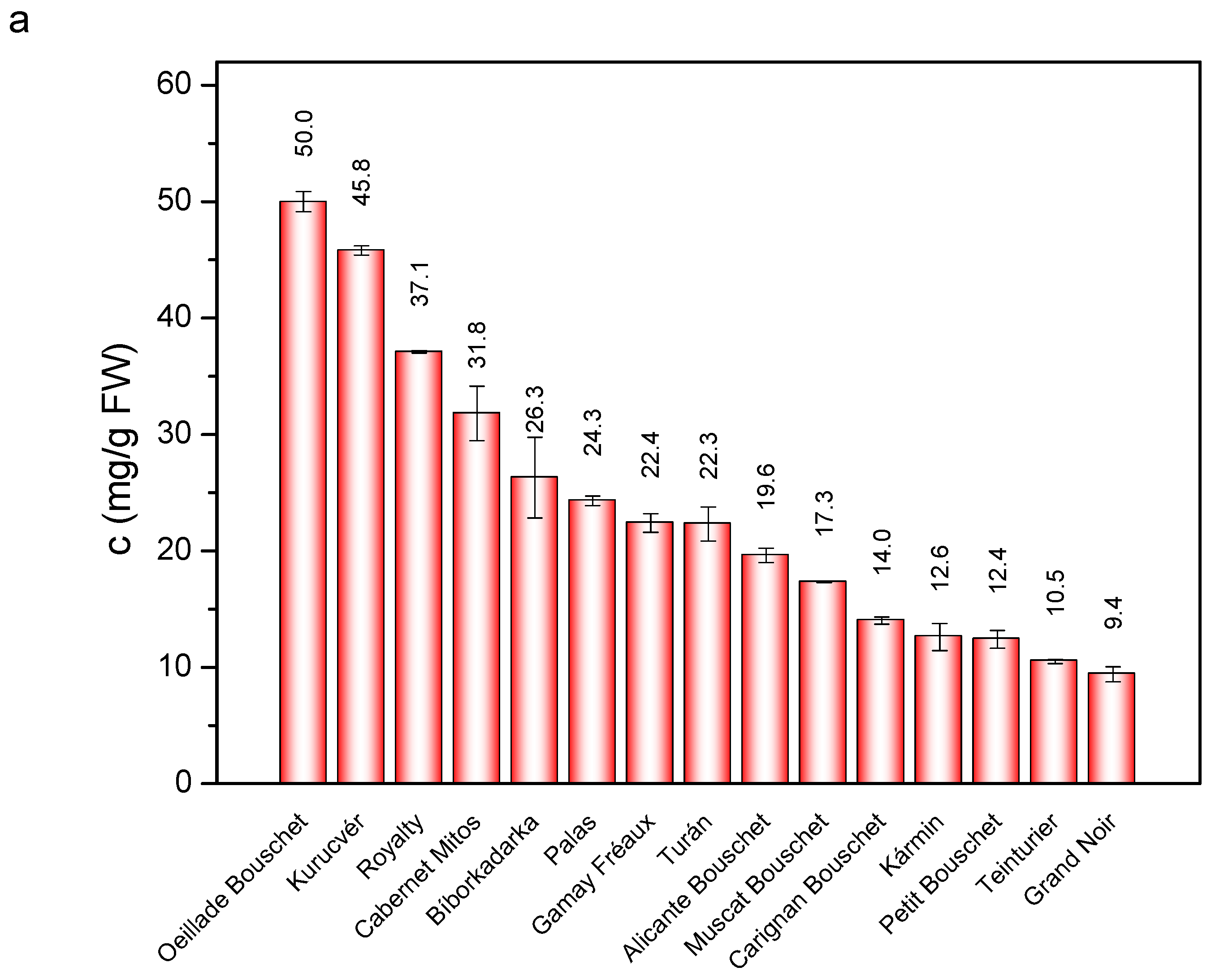
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Santiago, J.-L.; González, I.; Gago, P.; Alonso-Villaverde, V.; Boso, S.; Martínez, M. Identification of and relationships among a number ofteinturiergrapevines that expanded across Europe in the early 20th century. Aust. J. Grape Wine Res. 2008, 14, 223–229. [Google Scholar] [CrossRef]
- Tamura, H.; Hayashi, Y.; Sugisawa, H.; Kondo, T. Structure determination of acylated anthocyanins in Muscat Bailey a grapes by homonuclear Hartmann-Hahn (HOHAHA) spectroscopy and liquid chromatography-mass spectrometry. Phytochem. Anal. 1994, 5, 190–196. [Google Scholar] [CrossRef]
- Schwarz, M.; Winterhalter, P. Novel Aged Anthocyanins from Pinotage Wines: Isolation, Characterization, and Pathway of Formation. In Red Wine Color; Waterhouse, A.L., Kennedy, J.A., Eds.; American Chemical Society: Washington, DC, USA, 2004; Volume 886, pp. 179–197. [Google Scholar]
- Brouillard, R.; Delaporte, B. Chemistry of anthocyanin pigments. 2. Kinetic and thermodynamic study of proton transfer, hydration, and tautomeric reactions of malvidin 3-glucoside. J. Am. Chem. Soc. 1977, 99, 8461–8468. [Google Scholar] [CrossRef]
- Ockermann, P.; Headley, L.; Lizio, R.; Hansmann, J. A Review of the Properties of Anthocyanins and Their Influence on Factors Affecting Cardiometabolic and Cognitive Health. Nutrients 2021, 13, 2831. [Google Scholar] [CrossRef] [PubMed]
- Markakis, P. (Ed.) Anthocyanins as Food Colors, 1st ed.; Academic Press: Cambridge, MA, USA, 1982; p. 280. [Google Scholar]
- Santos-Buelga, C.; Mateus, N.; De Freitas, V. Anthocyanins. Plant Pigments and Beyond. J. Agric. Food Chem. 2014, 62, 6879–6884. [Google Scholar] [CrossRef] [PubMed]
- Yue, Q.; Xu, L.; Xiang, G.; Yu, X.; Yao, Y. Characterization of Gene Expression Profile, Phenolic Composition, and Antioxidant Capacity in Red-Fleshed Grape Berries and Their Wines. J. Agric. Food Chem. 2018, 66, 7190–7199. [Google Scholar] [CrossRef]
- Kalt, W.; Cassidy, A.; Howard, L.R.; Krikorian, R.; Stull, A.J.; Tremblay, F.; Zamora-Ros, R. Recent Research on the Health Benefits of Blueberries and Their Anthocyanins. Adv. Nutr. Int. Rev. J. 2020, 11, 224–236. [Google Scholar] [CrossRef]
- Li, D.; Wang, P.; Luo, Y.; Zhao, M.; Chen, F. Health benefits of anthocyanins and molecular mechanisms: Update from recent decade. Crit. Rev. Food Sci. Nutr. 2017, 57, 1729–1741. [Google Scholar] [CrossRef]
- Wang, X.; Yang, D.-Y.; Yang, L.-Q.; Zhao, W.-Z.; Cai, L.-Y.; Shi, H.-P. Anthocyanin Consumption and Risk of Colorectal Cancer: A Meta-Analysis of Observational Studies. J. Am. Coll. Nutr. 2019, 38, 470–477. [Google Scholar] [CrossRef]
- Alam, A.; Islam, P.; Subhan, N.; Rahman, M.; Khan, F.; Burrows, G.E.; Nahar, L.; Sarker, S.D. Potential health benefits of anthocyanins in oxidative stress related disorders. Phytochem. Rev. 2021, 20, 705–749. [Google Scholar] [CrossRef]
- Liu, C.; Sun, J.; Lu, Y.; Bo, Y. Effects of Anthocyanin on Serum Lipids in Dyslipidemia Patients: A Systematic Review and Meta-Analysis. PLoS ONE 2016, 11, e0162089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daneshzad, E.; Shab-Bidar, S.; Mohammadpour, Z.; Djafarian, K. Effect of anthocyanin supplementation on cardio-metabolic biomarkers: A systematic review and meta-analysis of randomized controlled trials. Clin. Nutr. 2019, 38, 1153–1165. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Ling, W.; Du, Z.; Chen, Y.; Li, D.; Deng, S.; Liu, Z.; Yang, L. Effects of Anthocyanins on Cardiometabolic Health: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Adv. Nutr. Int. Rev. J. 2017, 8, 684–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamakoshi, J.; Kataoka, S.; Koga, T.; Ariga, T. Proanthocyanidin-rich extract from grape seeds attenuates the development of aortic atherosclerosis in cholesterol-fed rabbits. Atherosclerosis 1999, 142, 139–149. [Google Scholar] [CrossRef]
- Brown, J.E.; Kelly, M.F. Inhibition of lipid peroxidation by anthocyanins, anthocyanidins and their phenolic degradation products. Eur. J. Lipid Sci. Technol. 2007, 109, 66–71. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Hong, J.-H.; Jung, H.K.; Jeong, Y.S.; Cho, K.-H. Grape skin and loquat leaf extracts and acai puree have potent anti-atherosclerotic and anti-diabetic activity in vitro and in vivo in hypercholesterolemic zebrafish. Int. J. Mol. Med. 2012, 30, 606–614. [Google Scholar] [CrossRef] [Green Version]
- Les, F.; Cásedas, G.; Gómez, C.; Moliner, C.; Valero, M.S.; López, V. The role of anthocyanins as antidiabetic agents: From molecular mechanisms to in vivo and human studies. J. Physiol. Biochem. 2021, 77, 109–131. [Google Scholar] [CrossRef]
- Pattananandecha, T.; Apichai, S.; Sirilun, S.; Julsrigival, J.; Sawangrat, K.; Ogata, F.; Kawasaki, N.; Sirithunyalug, B.; Saenjum, C. Anthocyanin Profile, Antioxidant, Anti-Inflammatory, and Antimicrobial against Foodborne Pathogens Activities of Purple Rice Cultivars in Northern Thailand. Molecules 2021, 26, 5234. [Google Scholar] [CrossRef]
- Azzini, E.; Giacometti, J.; Russo, G.L. Antiobesity Effects of Anthocyanins in Preclinical and Clinical Studies. Oxidative Med. Cell. Longev. 2017, 2017, 2740364. [Google Scholar] [CrossRef]
- Belwal, T.; Nabavi, S.F.; Nabavi, S.M.; Habtemariam, S. Dietary Anthocyanins and Insulin Resistance: When Food Becomes a Medicine. Nutrients 2017, 9, 1111. [Google Scholar] [CrossRef]
- Fallah, A.A.; Sarmast, E.; Fatehi, P.; Jafari, T. Impact of dietary anthocyanins on systemic and vascular inflammation: Systematic review and meta-analysis on randomised clinical trials. Food Chem. Toxicol. 2020, 135, 110922. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Choi, M.; Lee, M. Effects of Anthocyanin Supplementation on Reduction of Obesity Criteria: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Nutrients 2021, 13, 2121. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Ding, S.; Fei, Y.; Liu, G.; Jang, H.; Fang, J. Antimicrobial activity of anthocyanins and catechins against foodborne pathogens Escherichia coli and Salmonella. Food Control 2019, 106, 106712. [Google Scholar] [CrossRef]
- Igwe, E.O.; Charlton, K.E.; Probst, Y.C.; Kent, K.; Netzel, M.E. A systematic literature review of the effect of anthocyanins on gut microbiota populations. J. Hum. Nutr. Diet. 2019, 32, 53–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontaine, M.C.; Labbé, F.; Dussert, Y.; Delière, L.; Richart-Cervera, S.; Giraud, T.; Delmotte, F. Europe as a bridgehead in the worldwide invasion history of grapevine downy mildew, Plasmopara viticola. Curr. Biol. 2021, 31, 2155–2166.e4. [Google Scholar] [CrossRef]
- Castillo-Muñoz, N.; Fernández-González, M.; Gómez-Alonso, S.; García-Romero, E.; Hermosín-Gutiérrez, I. Red-Color Related Phenolic Composition of Garnacha Tintorera (Vitis vinifera L.) Grapes and Red Wines. J. Agric. Food Chem. 2009, 57, 7883–7891. [Google Scholar] [CrossRef]
- He, F.; Liang, N.-N.; Duan, C.-Q.; Wang, J.; Reeves, M. Accumulation of anthocyanins in the skin and pulp of five different teinturier grapes. Acta Hortic. 2017, 73–82. [Google Scholar] [CrossRef]
- Guan, L.; Dai, Z.; Wu, B.-H.; Wu, J.; Merlin, I.; Hilbert, G.; Renaud, C.; Gomès, E.; Edwards, E.; Li, S.-H.; et al. Anthocyanin biosynthesis is differentially regulated by light in the skin and flesh of white-fleshed and teinturier grape berries. Planta 2016, 243, 23–41. [Google Scholar] [CrossRef]
- Ageorges, A.; Fernandez, L.; Vialet, S.; Merdinoglu, D.; Terrier, N.; Romieu, C. Four specific isogenes of the anthocyanin metabolic pathway are systematically co-expressed with the red colour of grape berries. Plant Sci. 2006, 170, 372–383. [Google Scholar] [CrossRef]
- Röckel, F.; Moock, C.; Braun, U.; Schwander, F.; Cousins, P.; Maul, E.; Töpfer, R.; Hausmann, L. Color Intensity of the Red-Fleshed Berry Phenotype of Vitis vinifera Teinturier Grapes Varies Due to a 408 bp Duplication in the Promoter of VvmybA1. Genes 2020, 11, 891. [Google Scholar] [CrossRef]
- Maul, E.; Sudharma, K.N.; Kecke, S.; Marx, G.; Müller, C.; Audeguin, L.; Boselli, M.; Boursiquot, J.M.; Bucchetti, B.; Cabello, F.; et al. The European Vitis Database (www.eu-vitis.de)—A technical innovation through an online uploading and interactive modification system. Vitis 2012, 51, 79–85. [Google Scholar] [CrossRef]
- Huber, F.; Röckel, F.; Schwander, F.; Maul, E.; Eibach, R.; Cousins, P.; Töpfer, R. A view into American grapevine history: Vitis vinifera cv. ‘Sémillon’ is an ancestor of ‘Catawba’ and ‘Concord’. Vitis 2016, 55, 53–56. [Google Scholar] [CrossRef]
- Brouillard, R.; Chassaing, S.; Fougerousse, A. Why are grape/fresh wine anthocyanins so simple and why is it that red wine color lasts so long? Phytochemistry 2003, 64, 1179–1186. [Google Scholar] [CrossRef]
- Chen, W.-K.; Wang, Y.; Gao, X.-T.; Yang, X.-H.; He, F.; Duan, C.-Q.; Wang, J. Flavonoid and aromatic profiles of two Vitis vinifera L. teinturier grape cultivars. Aust. J. Grape Wine Res. 2018, 24, 379–389. [Google Scholar] [CrossRef]
- He, J.-J.; Liu, Y.-X.; Pan, Q.-H.; Cui, X.-Y.; Duan, C.-Q. Different Anthocyanin Profiles of the Skin and the Pulp of Yan73 (Muscat Hamburg × Alicante Bouschet) Grape Berries. Molecules 2010, 15, 1141–1153. [Google Scholar] [CrossRef]
- Xie, S.; Song, C.; Wang, X.; Liu, M.; Zhang, Z.; Xi, Z. Tissue- Specific Expression Analysis of Anthocyanin Biosynthetic Genes in White- and Red-Fleshed Grape Cultivars. Molecules 2015, 20, 22767–22780. [Google Scholar] [CrossRef] [Green Version]
- Guan, L.; Li, J.-H.; Fan, P.-G.; Chen, S.; Fang, J.-B.; Li, S.-H.; Wu, B.-H. Anthocyanin Accumulation in Various Organs of a Teinturier Cultivar (Vitis vinifera L.) during the Growing Season. Am. J. Enol. Vitic. 2012, 63, 177–184. [Google Scholar] [CrossRef]
- He, F.; He, J.-J.; Pan, Q.-H.; Duan, C.-Q. Mass-spectrometry evidence confirming the presence of pelargonidin-3-O-glucoside in the berry skins of Cabernet Sauvignon and Pinot Noir (Vitis vinifera L.). Aust. J. Grape Wine Res. 2010, 16, 464–468. [Google Scholar] [CrossRef]
- Kong, J.; Wu, J.; Guan, L.; Hilbert, G.; Delrot, S.; Fan, P.; Liang, Z.; Wu, B.; Matus, J.T.; Gomès, E.; et al. Metabolite analysis reveals distinct spatio-temporal accumulation of anthocyanins in two teinturier variants of cv. ‘Gamay’ grapevines (Vitis vinifera L.). Planta 2021, 253, 84. [Google Scholar] [CrossRef]
- Zhang, K.; Liu, Z.; Guan, L.; Zheng, T.; Jiu, S.; Zhu, X.; Jia, H.; Fang, J. Changes of Anthocyanin Component Biosynthesis in ‘Summer Black’ Grape Berries after the Red Flesh Mutation Occurred. J. Agric. Food Chem. 2018, 66, 9209–9218. [Google Scholar] [CrossRef]
- Carreño, J.; Almela, L.; Martínez, A.; Fernández-López, J.A. Chemotaxonomical Classification of Red Table Grapes based on Anthocyanin Profile and External Colour. LWT Food Sci. Technol. 1997, 30, 259–265. [Google Scholar] [CrossRef]
- Wang, H.; Race, A.E.J.; Shrikhande, A.J. Characterization of Anthocyanins in Grape Juices by Ion Trap Liquid Chromatography−Mass Spectrometry. J. Agric. Food Chem. 2003, 51, 1839–1844. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Pan, Q.-H.; Duan, C.-Q.; Wang, J. Anthocyanin profiling of the berry skins of five Vitis amurensis grapes and one related hybrid cultivar. Acta Hortic. 2014, 531–538. [Google Scholar] [CrossRef]
- Zhao, Q.; Duan, C.-Q.; Wang, J. Anthocyanins Profile of Grape Berries of Vitis amurensis, Its Hybrids and Their Wines. Int. J. Mol. Sci. 2010, 11, 2212–2228. [Google Scholar] [CrossRef] [Green Version]
- Stalmach, A.; Edwards, C.A.; Wightman, J.D.; Crozier, A. Identification of (Poly)phenolic Compounds in Concord Grape Juice and Their Metabolites in Human Plasma and Urine after Juice Consumption. J. Agric. Food Chem. 2011, 59, 9512–9522. [Google Scholar] [CrossRef]
- Xing, R.-R.; He, F.; Xiao, H.-L.; Duan, C.-Q.; Pan, Q.-H. Accumulation Pattern of Flavonoids in Cabernet Sauvignon Grapes Grown in a Low-Latitude and High-Altitude Region. S. Afr. J. Enol. Vitic. 2015, 36, 32–43. [Google Scholar] [CrossRef] [Green Version]
- Ju, Y.; Yang, L.; Yue, X.; Li, Y.; He, R.; Deng, S.; Yang, X.; Fang, Y. Anthocyanin profiles and color properties of red wines made from Vitis davidii and Vitis vinifera grapes. Food Sci. Hum. Wellness 2021, 10, 335–344. [Google Scholar] [CrossRef]
- Ma, Y.; Ma, X.; Gao, X.; Wu, W.; Zhou, B. Light Induced Regulation Pathway of Anthocyanin Biosynthesis in Plants. Int. J. Mol. Sci. 2021, 22, 11116. [Google Scholar] [CrossRef]
- He, F.; Mu, L.; Yan, G.-L.; Liang, N.-N.; Pan, Q.-H.; Wang, J.; Reeves, M.J.; Duan, C.-Q. Biosynthesis of Anthocyanins and Their Regulation in Colored Grapes. Molecules 2010, 15, 9057–9091. [Google Scholar] [CrossRef] [Green Version]
- Bogs, J.; Ebadi, A.; McDavid, D.; Robinson, S.P. Identification of the Flavonoid Hydroxylases from Grapevine and Their Regulation during Fruit Development. Plant Physiol. 2006, 140, 279–291. [Google Scholar] [CrossRef]
- Fraige, K.; Pereira-Filho, E.R.; Carrilho, E. Fingerprinting of anthocyanins from grapes produced in Brazil using HPLC–DAD–MS and exploratory analysis by principal component analysis. Food Chem. 2014, 145, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Budić-Leto, I.; Mucalo, A.; Ljubenkov, I.; Zdunić, G. Anthocyanin profile of wild grape Vitis vinifera in the eastern Adriatic region. Sci. Hortic. 2018, 238, 32–37. [Google Scholar] [CrossRef]
- Jin, Z.-M.; He, J.-J.; Bi, H.-Q.; Cui, X.-Y.; Duan, C.-Q. Phenolic Compound Profiles in Berry Skins from Nine Red Wine Grape Cultivars in Northwest China. Molecules 2009, 14, 4922–4935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.; Zhang, Y.; Lu, J. Phenolic Contents and Compositions in Skins of Red Wine Grape Cultivars among Various Genetic Backgrounds and Originations. Int. J. Mol. Sci. 2012, 13, 3492–3510. [Google Scholar] [CrossRef] [Green Version]
- Stój, A.; Kapusta, I.; Domagała, D. Classification of Red Wines Produced from Zweigelt and Rondo Grape Varieties Based on the Analysis of Phenolic Compounds by UPLC-PDA-MS/MS. Molecules 2020, 25, 1342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fanzone, M.; Zamora, F.; Jofré, V.; Assof, M.; Gómez-Cordovés, C.; Peña-Neira, A. Phenolic characterisation of red wines from different grape varieties cultivated in Mendoza province (Argentina). J. Sci. Food Agric. 2012, 92, 704–718. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Sharifi-Rad, J.; Cappellini, F.; Reiner, Z.; Zorzan, D.; Imran, M.; Sener, B.; Kilic, M.; El-Shazly, M.; Fahmy, N.M.; et al. The Therapeutic Potential of Anthocyanins: Current Approaches Based on Their Molecular Mechanism of Action. Front. Pharmacol. 2020, 11, 1300. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.; Qiao, X.; Chen, H.; Nan, H.; Zhang, Z. Coordinated Regulation of Grape Berry Flesh Color by Transcriptional Activators and Repressors. J. Agric. Food Chem. 2019, 67, 11815–11824. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ion Source | HESI |
|---|---|
| Spray voltage | +/− 3 keV |
| Capillary temperature | 350 °C |
| Probe heater temperature | 300 °C |
| Flow rate of sheath gas (N2) | 30 (a.u.) |
| Flow rate of auxiliary gas (N2) | 10 (a.u.) |
| Resolution of Full Scan | 35,000 |
| Resolution of All Ion Fragmentation | 17,500 |
| S-Lens RF Level: | 50% |
| Collision gas/energy | N2/30 eV |
| No. | RT (min) | Compound | [M + H]+ (amu) | Positive Fragment Ions | [M − H]− (amu) | Negative Fragment Ions |
|---|---|---|---|---|---|---|
| 1 | 5.73 ± 0.03 | Delphinidin-3-O-glucoside | 465.1022 | 303.0495 (100), 304.0527 (16) | 463.0889 | 300.0279 (100), 301.0344 (54) |
| 2 | 7.07 ± 0.03 | Petunidin-3,5-di-O-glucoside | 641.1701 | 317.0650 (100), 479.1170 (17), 318.0681 (16) | 639.1564 | 477.1032 (100), 315.0517 (67), 355.0669 (43) |
| 3 | 7.26 ± 0.04 | Cyanidin-3-O-glucoside | 449.1071 | 287.0544 (100), 288.0577 (17) | 447.0934 | 284.0327 (100), 285.0396 (87) |
| 4 | 8.65 ± 0.03 | Peonidin-3,5-di-O-glucoside | 625.1760 | 301.0703 (100), 302.0736 (18), 463.1230 (18) | 623.1627 | 479.1203 (100), 299.0564 (88), 461.1096 (84), 317.0668 (70) |
| 5 | 8.90 ± 0.05 | Pelargonidin-3-O-glucoside | 433.1123 | 271.0591 (100), 272.0618 (17) | 431.0982 | 269.0448 (100), 268.0378 (81), 270.0482 (16), 431.0982 (12) |
| 6 | 9.67 ± 0.04 | Petunidin-3-O-glucoside | 479.1182 | 317.0652 (100), 318.0685 (18) | 477.1043 | 314.0437 (100), 315.0508 (87), 300.0273 (21), 316.0542 (16) |
| 7 | 10.51 ± 0.05 | Malvidin-3,5-di-O-glucoside | 655.1864 | 331.0806 (100), 332.0838 (19), 493.1335 (18) | 653.1729 | 509.1306 (100), 329.0668 (60), 347.0775 (59), 491.1198 (54) |
| 8 | 11.51 ± 0.04 | Peonidin-3-O-glucoside | 463.1230 | 301.0702 (100), 302.0736 (17), 349.1827 (12) | 461.1094 | 299.0560 (100), 298.0486 (66), 284.0326 (21), 300.0602 (17) |
| 9 | 13.94 ± 0.05 | Malvidin-3-O-glucoside | 493.1337 | 331.0807 (100), 332.0839 (19) | 491.1200 | 329.0664 (100), 328.0592 (63), 314.0430 (19), 330.0698 (18) |
| 10 | 16.08 ± 0.04 | Delphinidin-3-O-(6-O-acetyl-glucoside) | 507.1132 | 303.0497 (100), 304.0530 (17) | 505.0994 | 300.0277 (100), 301.0346 (60) |
| 11 | 18.70 ± 0.05 | Malvidin-3-O-(6-O-acetyl-glucoside)-5-O-glucoside | 697.1964 | 331.0810 (100), 349.1829 (59), 535.1443 (34), 332.0844 (19) | 695.1840 | 509.1310 (100), 347.0779 (54), 329.0668 (49), 491.1180 (45) |
| 12 | 21.42 ± 0.05 | Petunidin-3-O-(6-O-acetyl-glucoside) | 521.1287 | 437.2350 (100), 317.0650 (88), 437.2350 (19), 318.0683 (15) | 519.1146 | 314.0436 (100), 315.0509 (96), 316.0542 (18) |
| 13 | 24.22 ± 0.06 | Peonidin-3-O-(6-O-acetyl-glucoside) | 505.1333 | 301.0700 (100), 302.0732 (17), 525.2872 (12) | 503.1197 | 299.0557 (100), 298.0483 (67), 300.0592 (17), 284.0323 (13) |
| 14 | 25.68 ± 0.06 | Delphinidin-3-O-(6-O-p-coumaroyl-glucoside) | 611.1389 | 303.0496 (100), 525.2875 (21), 304.0529 (17) | 609.1257 | 300.0278 (100), 301.0340 (41), 609.1265 (14) |
| 15 | 26.69 ± 0.06 | Malvidin-3-O-(6-O-acetyl-glucoside) | 535.1444 | 331.0807 (100), 525.2877 (28), 332.0841 (18) | 533.1306 | 329.0667 (100), 328.0592 (62), 330.0700 (18) |
| 16 | 28.44 ± 0.08 | Cyanidin-3-O-(6-O-p-coumaroyl-glucoside) | 595.1440 | 287.0544 (100), 301.0695 (33) | 593.1306 | 284.0327 (100), 285.0399 (73), 299.0564 (44) |
| 17 | 30.05 ± 0.08 | Malvidin-3-O-(6-O-coumaroyl-glucoside)-5-O-glucoside | 801.2231 | 331.0808 (100), 639.1703 (66), 525.2877 (46), 640.1735 (25) | 799.2097 | 509.1307 (100), 491.1201 (45), 637.1574 (39), 329.0670 (27) |
| 18 | 30.29 ± 0.05 | Malvidin-3-O-(6-O-cis-p-coumaroyl-glucoside) | 639.1700 | 331.0807 (100), 332.0829 (17) | 637.1567 | 329.0670 (100), 328.0583 (35), 330.0700 (18) |
| 19 | 30.85 ± 0.06 | Petunidin-3-O-(6-O-trans-p-coumaroyl-glucoside) | 625.1548 | 317.0651 (100), 318.0684 (17), 613.3401 (12) | 623.1414 | 314.0437 (100), 315.0506 (80), 623.1417 (15), 316.0543 (13) |
| 20 | 33.50 ± 0.06 | Peonidin-3-O-(6-O-trans-p-coumaroyl-glucoside) | 609.1592 | 301.0698 (100), 302.0730 (18) | 607.1461 | 299.0559 (100), 298.0484 (34), 300.0593 (17) |
| 21 | 35.30 ± 0.06 | Malvidin-3-O-(6-O-trans-p-coumaroyl-glucoside) | 639.1700 | 331.0806 (100), 332.0838 (19) | 637.1572 | 329.0668 (100), 328.0592 (32), 330.0702 (18) |
| Dph-glc | Cyd-glc | Plg-glc | Ptd-glc | Mvd-3,5-diglc | Pnd-glc | Mvd-glc | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cultivars | Skin | Juice | Skin | Juice | Skin | Juice | Skin | Juice | Skin | Juice | Skin | Juice | Skin | Juice |
| Alicante Bouschet | 451 ± 42 | trace | 132 ± 3 | 1.7 ± 0.1 | trace | trace | 781 ± 40 | Nd | nd | trace | 2790 ± 60 | 80.8 ± 3.3 | 10,005 ± 214 | 22.8 ± 0.3 |
| Bíborkadarka | 768 ± 110 | trace | 207 ± 25 | 2.5 ± 0.4 | trace | 2.2 ± 0.1 | 1218 ± 181 | Nd | nd | trace | 2964 ± 317 | 90.4 ± 4.6 | 13,505 ± 1867 | 32.0 ± 1.6 |
| Cabernet Mitos | 1817 ± 68 | trace | 365 ± 3 | 41.7 ± 1.8 | trace | trace | 2316 ± 75 | 11.2±0.6 | nd | trace | 3398 ± 227 | 381.6 ± 24.9 | 13,665 ± 1580 | 320.3 ± 6.3 |
| Carignan Bouchset | 144 ± 8 | trace | trace | 4.5 ± 0.2 | nd | trace | 284 ± 7 | trace | nd | nd | 631 ± 13 | 118.0 ± 3.7 | 5987 ± 25 | 47.0 ± 2.3 |
| Gamay Fréaux | 949 ± 21 | trace | 230 ± 1 | 5.5 ± 0.4 | trace | trace | 1537 ± 82 | trace | nd | trace | 2755 ± 37 | 153.8 ± 6.8 | 15,045 ± 748 | 79.9 ± 3.4 |
| Grand Noir | 170 ± 10 | trace | 78 ± 13 | 1.0 ± 0.1 | trace | 2.4 ± 0.2 | 306 ± 10 | Nd | nd | trace | 3468 ± 395 | 100.9 ± 1.3 | 3894 ± 219 | 30.7 ± 0.4 |
| Kármin | 363 ± 30 | trace | trace | 1.8 ± 0.0 | nd | 1.1 ± 0.1 | 532 ± 49 | nd | nd | trace | 495 ± 48 | 71.4 ± 2.5 | 6281 ± 634 | 28.0 ± 1.1 |
| Kurucvér | 7933 ± 35 | 3.4 ± 0.2 | 738 ± 37 | 21.0 ± 2.8 | trace | 4.1 ± 0.1 | 4993 ± 132 | 6.1 ± 0.2 | 2168 ± 142 | trace | 927 ± 52 | 42.4 ± 1.3 | 15,201 ± 311 | 85.7 ± 2.8 |
| Muscat Bouschet | 408 ± 29 | trace | 115 ± 2 | 1.7 ± 0.2 | trace | 1.2 ± 0.1 | 631 ± 10 | Nd | 543 ± 10 | trace | 2263 ± 36 | 74.7 ± 2.6 | 6991 ± 40 | 16.2 ± 0.7 |
| Oeillade Bouschet | 7032 ± 237 | 2.9 ± 0.2 | 1054 ± 87 | 18.8 ± 0.7 | trace | trace | 5893 ± 240 | 6. 2 ± 0.9 | nd | trace | 2772 ± 230 | 143.3 ± 2.1 | 19,102 ± 901 | 93.4 ± 2.7 |
| Palas | 1700 ± 43 | trace | 410 ± 14 | 19.1 ± 0.7 | trace | trace | 2301 ± 24 | 6.4 ± 0.9 | nd | trace | 3237 ± 13 | 226.8 ± 9.1 | 11,454 ± 145 | 210.4 ± 7.9 |
| Petit Bouschet | 211 ± 24 | trace | 118 ± 6 | 2.9 ± 0.2 | trace | 1.6 ± 0.1 | 411 ± 38 | nd | nd | nd | 3959 ± 130 | 78.9 ± 3.1 | 5860 ± 65 | 13.7 ± 0.3 |
| Royalty | 611 ± 6 | 3.4 ± 0.7 | trace | 18.4 ± 0.5 | nd | nd | 795 ± 120 | 7.6 ± 0.3 | 15,264 ± 271 | 349.7 ± 6.9 | 337 ± 26 | 78.3 ± 0.9 | 2207 ± 176 | 51.6 ± 1.0 |
| Teinturier | 327 ± 18 | trace | 100 ± 13 | 2.3 ± 0.1 | nd | trace | 500 ± 25 | nd | nd | nd | 1402 ± 25 | 91.6 ± 3.1 | 5590 ± 103 | 15.7 ± 0.2 |
| Turán | 693.5 ± 52.4 | 5.0 ± 0.3 | 178 ± 15 | 14.4 ± 1.0 | nd | 1.8 ± 0.1 | 1065 ± 43 | 7.3 ± 0.7 | nd | trace | 1263 ± 52 | 106.6 ± 5.4 | 6633 ± 229 | 82.3 ± 4.0 |

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kőrösi, L.; Molnár, S.; Teszlák, P.; Dörnyei, Á.; Maul, E.; Töpfer, R.; Marosvölgyi, T.; Szabó, É.; Röckel, F. Comparative Study on Grape Berry Anthocyanins of Various Teinturier Varieties. Foods 2022, 11, 3668. https://doi.org/10.3390/foods11223668
Kőrösi L, Molnár S, Teszlák P, Dörnyei Á, Maul E, Töpfer R, Marosvölgyi T, Szabó É, Röckel F. Comparative Study on Grape Berry Anthocyanins of Various Teinturier Varieties. Foods. 2022; 11(22):3668. https://doi.org/10.3390/foods11223668
Chicago/Turabian StyleKőrösi, László, Szilárd Molnár, Péter Teszlák, Ágnes Dörnyei, Erika Maul, Reinhard Töpfer, Tamás Marosvölgyi, Éva Szabó, and Franco Röckel. 2022. "Comparative Study on Grape Berry Anthocyanins of Various Teinturier Varieties" Foods 11, no. 22: 3668. https://doi.org/10.3390/foods11223668
APA StyleKőrösi, L., Molnár, S., Teszlák, P., Dörnyei, Á., Maul, E., Töpfer, R., Marosvölgyi, T., Szabó, É., & Röckel, F. (2022). Comparative Study on Grape Berry Anthocyanins of Various Teinturier Varieties. Foods, 11(22), 3668. https://doi.org/10.3390/foods11223668