
Comparative Transcriptome Analysis to Investigate the Immunotoxicity Mechanism Triggered by Dimethomorph on Human Jurkat T Cell Lines


 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Jurkat Cell Culture
2.3. Cell Viability Assay
2.4. Cell Apoptosis Analysis
2.5. Transcriptome Analysis
2.5.1. RNA Extraction and High-Throughput Sequencing
2.5.2. Read Mapping and Differential Expression Analysis
2.6. Quantitative Real-Time PCR
2.7. Statistical Analysis
3. Results and Discussion
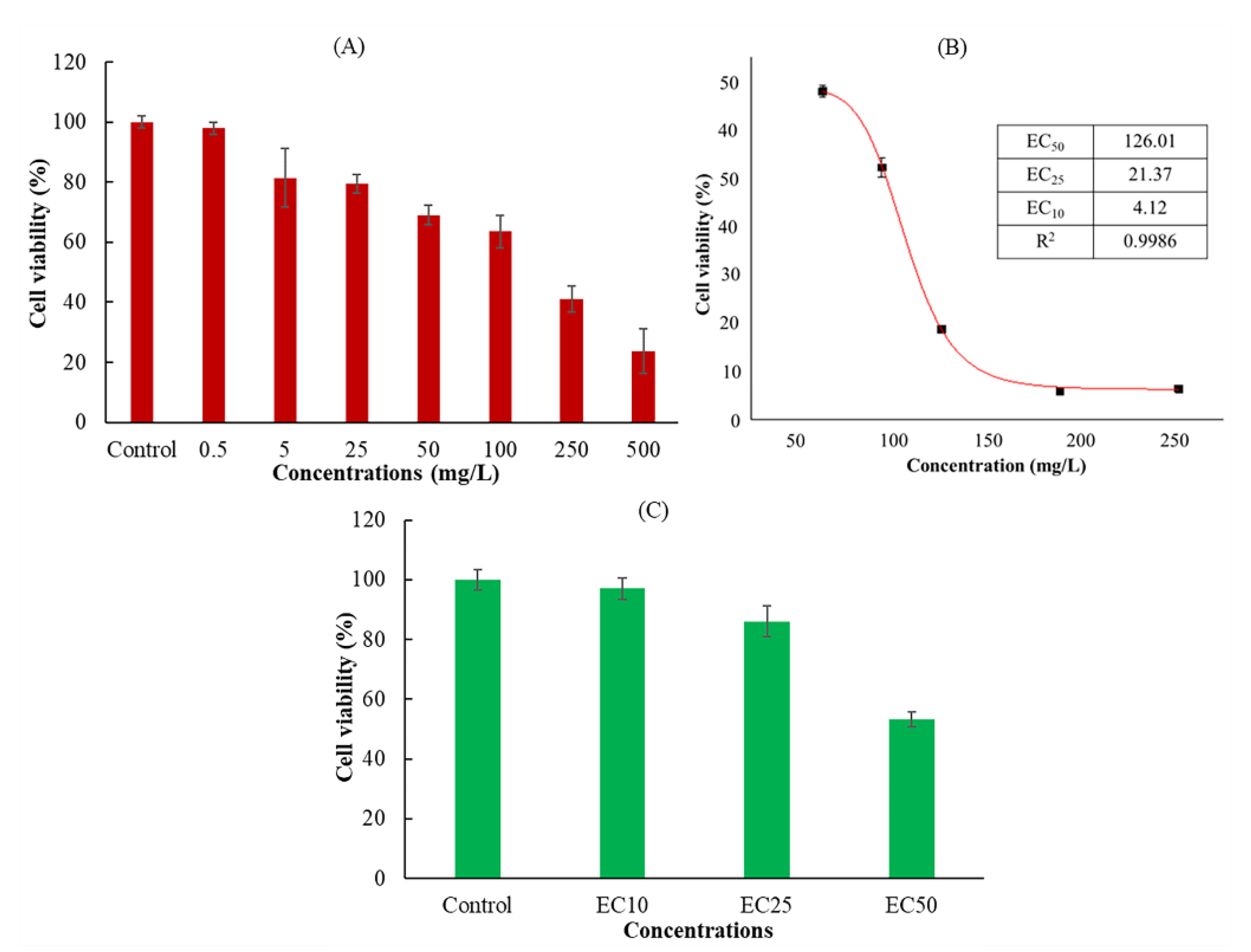
3.1. Effect of Dimethomorph on Cell Viability
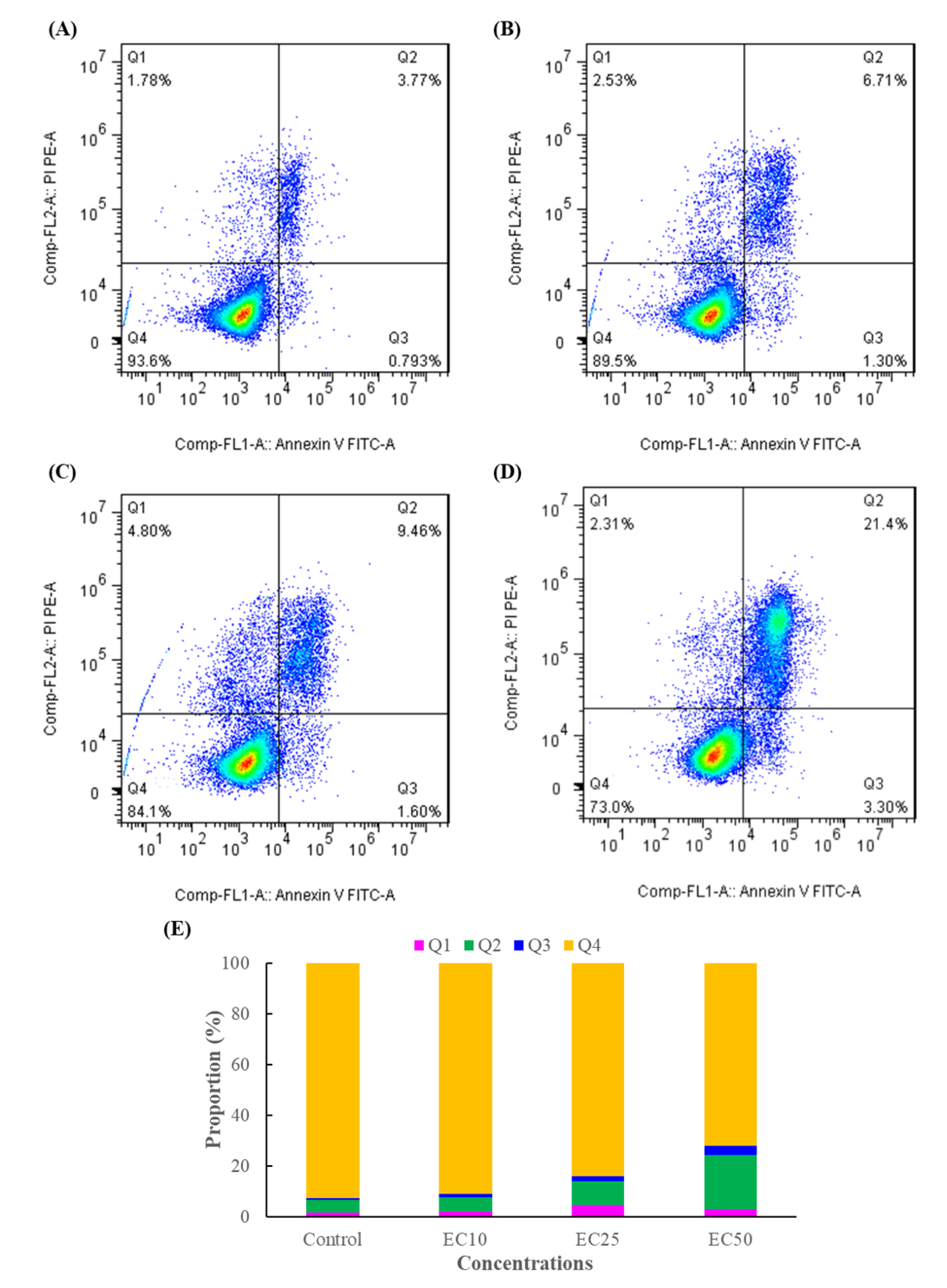
3.2. Effect of DMM on Cell Apoptosis
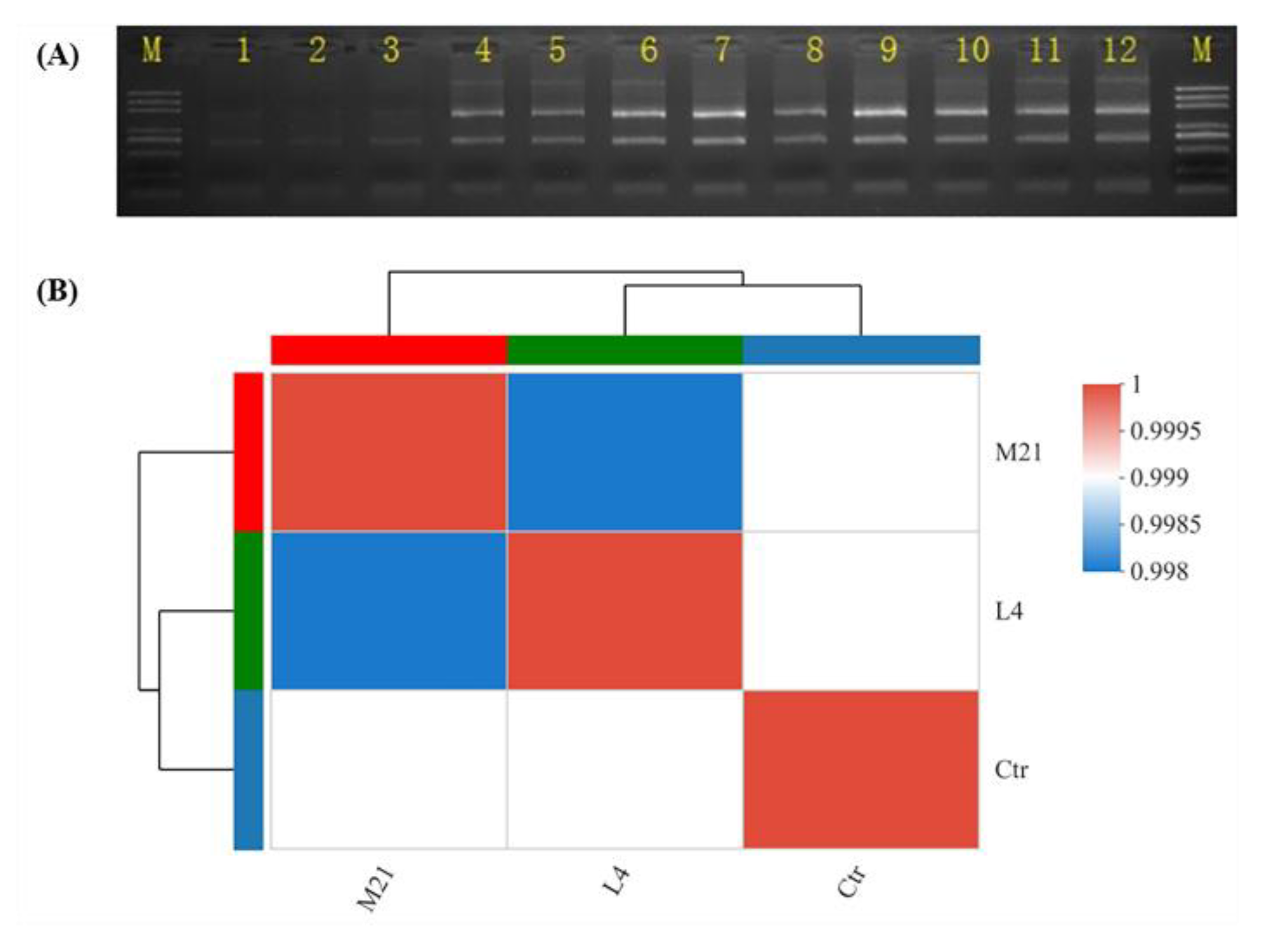
3.3. RNA Extraction and Quality Evaluation
3.4. RNA Sequencing Data Assessment
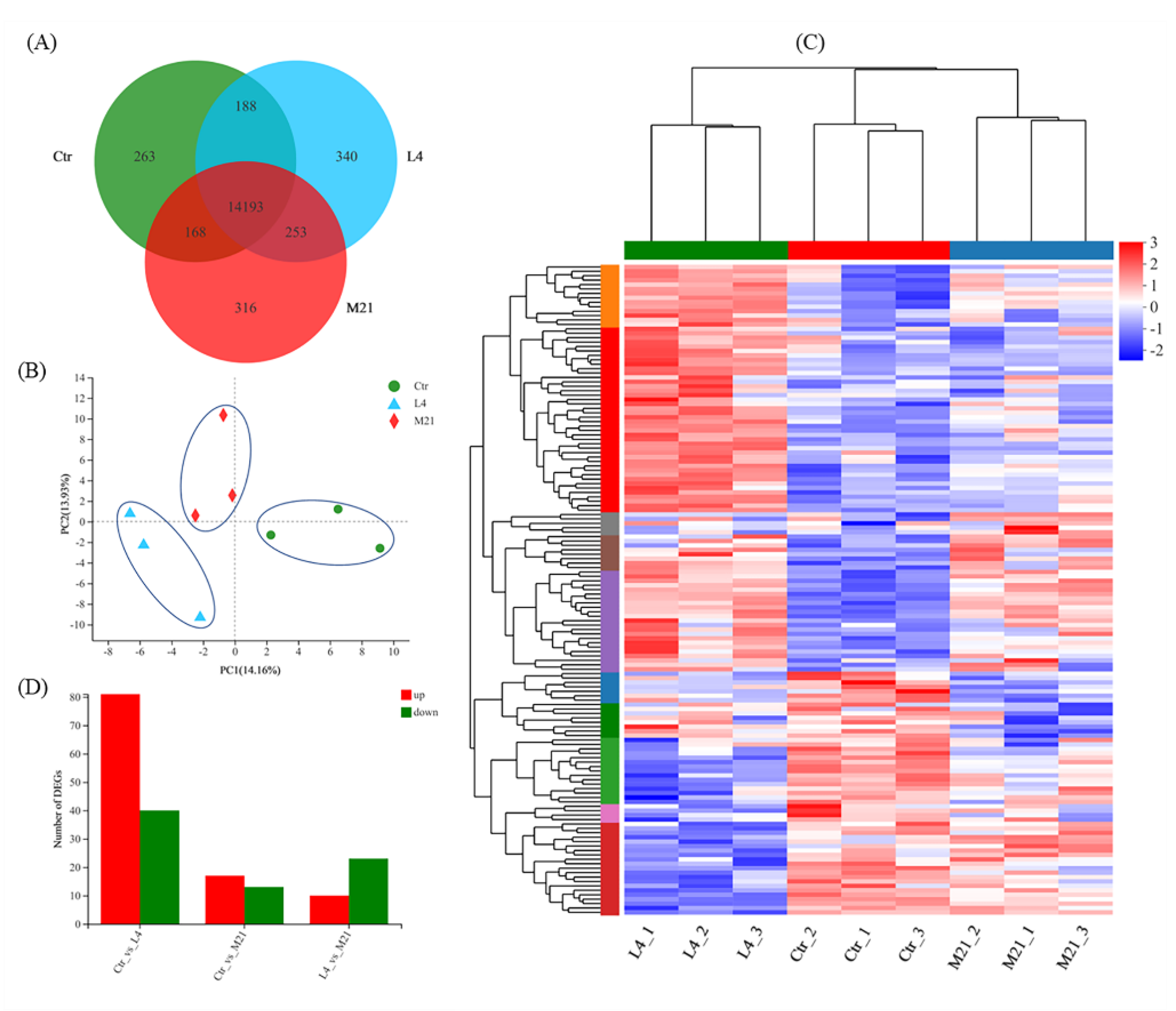
3.5. Gene Expression Overview
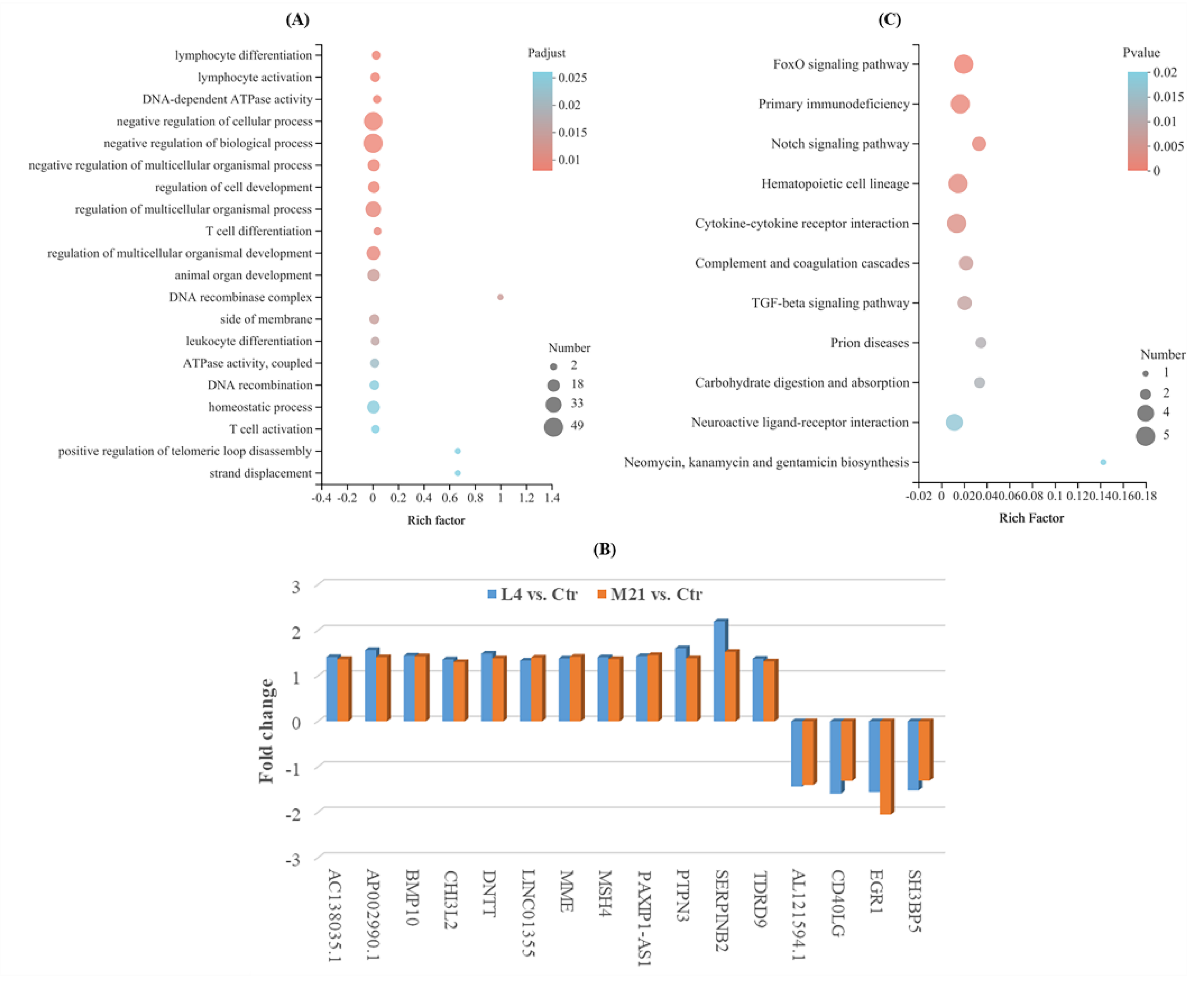
3.6. Gene Ontology (GO) Analysis of Differentially Expressed Genes (DEGs)
3.7. KEGG Enrichment Analysis of DEGs
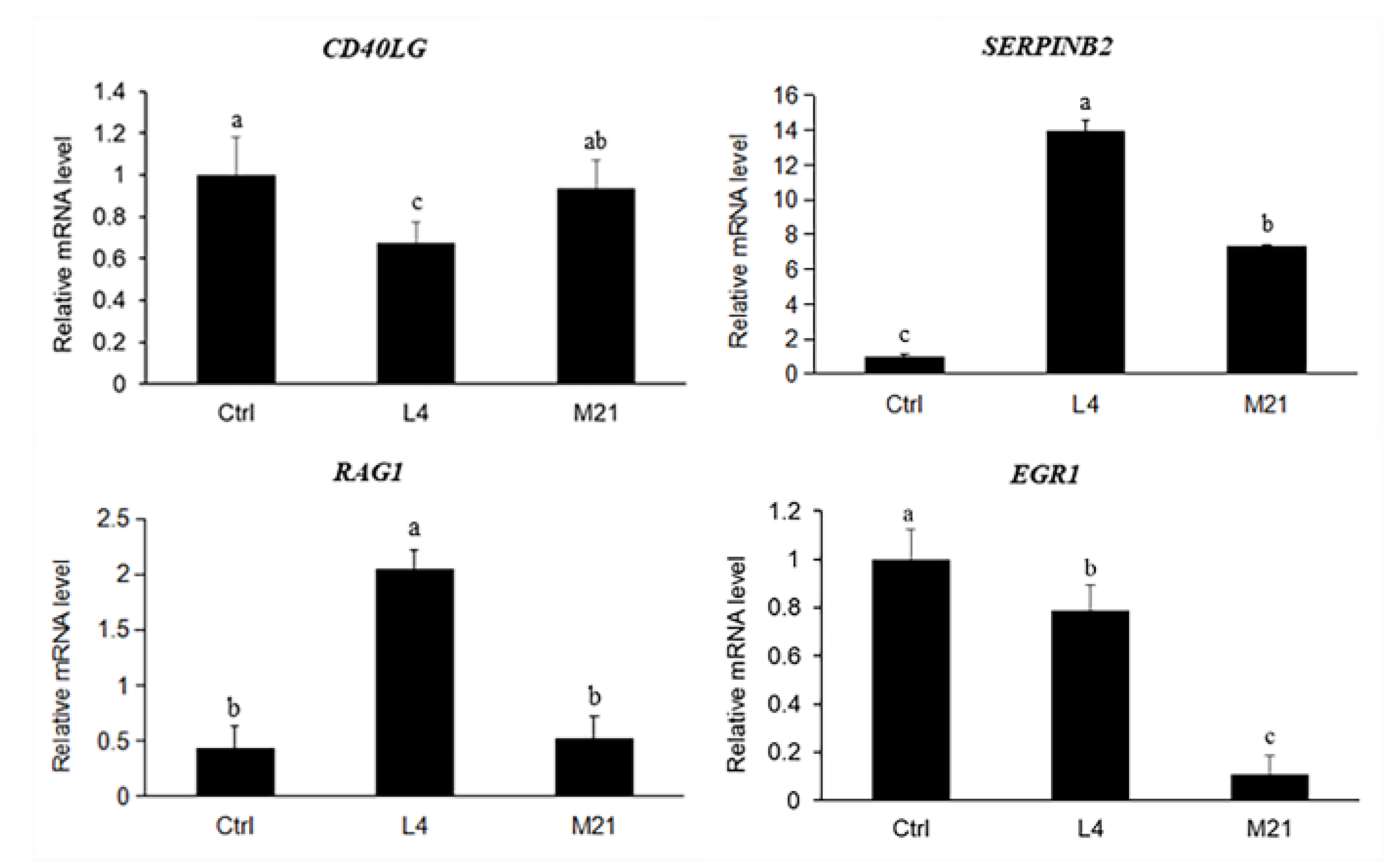
3.8. Target Gene Screening and Quantitative Real-Time PCR Validation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lawler, S.P. Environmental safety review of methoprene and bacterially-derived pesticides commonly used for sustained mosquito control. Ecotox. Environ. Safe. 2017, 139, 335–343. [Google Scholar] [CrossRef]
- Tudi, M.; Ruan, H.D.; Wang, L.; Lyu, J.; Sadler, R.; Connell, D.; Chu, C.; Phung, D.T. Agriculture development, pesticide application and its impact on the environment. Int. J. Environ. Res. Pub. He. 2021, 18, 1112. [Google Scholar] [CrossRef] [PubMed]
- Ji, C.Y.; Magnuson, J.T.; Zhang, W.; Zhao, M.R. New insight into the enantioselective cytotoxicity of cypermethrin: Imbalance between cell cycle and apoptosis. J. Hazard. Mater. 2021, 403, 123893. [Google Scholar] [CrossRef]
- Kim, K.H.; Kabir, E.; Jahan, S.A. Exposure to pesticides and the associated human health effects. Sci. Total Environ. 2017, 575, 525–535. [Google Scholar] [CrossRef]
- Fu, H.Y.; Tan, P.; Wang, R.J.; Li, S.N.; Liu, H.Z.; Yang, Y.; Wu, Z.L. Advances in organophosphorus pesticides pollution: Current status and challenges in ecotoxicological, sustainable agriculture, and degradation strategies. J. Hazard. Mater. 2022, 424, 127494. [Google Scholar] [CrossRef] [PubMed]
- Aktar, W.; Sengupta, D.; Chowdhury, A. Impact of pesticides use in agriculture: Their benefits and hazards. Interdiscipl. Toxicol. 2009, 2, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, M.M.; Chen, C.; Yang, G.L.; Wang, Y.H.; Wang, T.C.; Li, Y.; Qian, Y.Z. Combined anti-androgenic effects of mixtures of agricultural pesticides using in vitro and in silico methods. Ecotox. Environ. Safe. 2019, 186, 109652. [Google Scholar] [CrossRef] [PubMed]
- Arab, A.; Mostafalou, S. Neurotoxicity of pesticides in the context of CNS chronic diseases. Int. J. Environ. Health R. 2021, 32, 2718–2755. [Google Scholar] [CrossRef] [PubMed]
- Lerro, C.C.; Freeman, L.E.B.; DellaValle, C.T.; Andreotti, G.; Hofmann, J.N.; Koutros, S.; Parks, C.G.; Shrestha, S.; Alavanja, M.C.R.; Blair, A.; et al. Pesticide exposure and incident thyroid cancer among male pesticide applicators in agricultural health study. Environ. Int. 2021, 146, 106187. [Google Scholar] [CrossRef]
- Damalas, C.A.; Eleftherohorinos, I.G. Pesticide exposure, safety issues, and risk assessment indicators. Int. J. Environ. Res. Public Health 2011, 8, 1402–1419. [Google Scholar] [CrossRef] [PubMed]
- Dhouib, I.; Jallouli, M.; Annabi, A.; Marzouki, S.; Gharbi, N.; Elfazaa, S.; Lasram, M.M. From immunotoxicity to carcinogenicity: The effects of carbamate pesticides on the immune system. Environ. Sci. Pollut. R. 2016, 23, 9448–9458. [Google Scholar] [CrossRef] [PubMed]
- Naasri, S.; Helali, I.; Aouni, M.; Mastouri, M.; Harizi, H. N-acetylcysteine reduced the immunotoxicity effects induced in vitro by azoxystrobin and iprodione fungicides in mice. Environ. Toxicol. 2021, 36, 562–571. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.H.; Choi, K.C. Adverse effects of pesticides on the functions of immune system. Comp. Biochem. Phys. C 2020, 235, 108789. [Google Scholar]
- Hochstenbach, K.; Leeuwen, D.M.V.; Gmuender, H.; Stølevik, S.B.; Nygaard, U.C.; Løvik, M.; Granum, B.; Namork, E.; van Delft, J.H.M.; van Loveren, H. Transcriptomic profile indicative of immunotoxic exposure: In Vitro studies in peripheral blood mononuclear cells. Toxicol. Sci. 2010, 118, 19–30. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, S.C.; Sa-Correia, I. Yeast toxicogenomics: Lessons from a eukaryotic cell model and cell factory. Curr. Opin. Biotech. 2015, 33, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Rao, T.N.; Balaji, A.P.B.; Panagal, M.; Parvatamma, B.; Selvaraj, B.; Panneerselvam, S.; Aruni, W.; Subramanian, K.; Renuga, P.S.; Pandian, S. Nanoremediation of dimethomorph in water samples using magnesium aluminate nanoparticles. Environ. Technol. Inno. 2020, 20, 101176. [Google Scholar] [CrossRef]
- Wang, C.X.; Zhang, Q.M.; Wang, F.F.; Liang, W.X. Toxicological effects of dimethomorph on soil enzymatic activity and soil earthworm (Eisenia fetida). Chemosphere 2017, 169, 316–323. [Google Scholar] [CrossRef]
- Zhang, C.; Li, J.; Wu, X.; Long, Y.; An, H.; Pan, X.; Li, M.; Dong, F.; Zheng, Y. Rapid degradation of dimethomorph in polluted water and soil by Bacillus cereus WL08 immobilized on bamboo charcoal-sodium alginate. J. Hazard. Mater. 2020, 398, 122806. [Google Scholar] [CrossRef]
- Lin, S.; Tang, T.; Cang, T.; Yu, S.Q.; Ying, Z.T.; Gu, S.J.; Zhang, Q. The distributions of three fungicides in vegetables and their potential health risks in Zhejiang, China: A 3-year study (2015–2017). Environ. Pollut. 2020, 267, 115481. [Google Scholar] [CrossRef]
- Yang, L.P.; Zheng, Q.; Lin, S.K.; Wang, Y.Q.; Zhu, Q.Z.; Cheng, D.M.; Chen, J.J.; Zhang, Z.X. Dissipation and residue of dimethomorph in potato plants produced and dietary intake risk assessment. Int. J. Environ. Anal. Chem. 2020, 102, 1332–1344. [Google Scholar] [CrossRef]
- Shabeer, A.; Banerjee, T.P.K.; Jadhav, M.; Girame, R.; Utture, S.; Hingmire, S.; Oulkar, D. Residue dissipation and processing factor for dimethomorph, famoxadone and cymoxanil during raisin preparation. Food Chem. 2015, 170, 180–185. [Google Scholar] [CrossRef]
- Kabir, M.H.; Abd El-Aty, A.M.; Rahman, M.M.; Chung, H.S.; Lee, H.S.; Kim, M.R.; Chang, B.J.; Wang, J.; Shin, H.C.; Shim, J.H. Residual dynamic and risk assessment of dimethomorph in Swiss chard grown at two different sites. Biomed. Chromatogr. 2018, 32, e4053. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.; Berger, L.F.; Hendriksen, P.J.M.; Peijnenburg, A.A.C.M.; van Loveren, H.; Volger, O.L. Transcriptome-based functional classifiers for direct immunotoxicity. Arch. Toxicol. 2014, 88, 673–689. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.C.; Xu, J.B.; Brinkhof, B.; Wang, H.; Cui, Z.F.; Huang, W.E.; Ye, H. A single-cell Raman-based platform to identify developmental stages of human pluripotent stem cell-derived neurons. PNAS 2020, 117, 18412–18423. [Google Scholar] [CrossRef]
- Escriva, L.; Jennen, D.; Caiment, F.; Manyes, L. Transcriptomic study of the toxic mechanism triggered by beauvericin in Jurkat cells. Toxicol. Lett. 2018, 284, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-C.; Luo, Y.; Meng, F.-B.; Li, J.; Chen, W.-J.; Liu, D.-Y.; Zou, L.-H.; Zhou, L. Preparation and characterization of feruloylated oat β-glucan with antioxidant activity and colon-targeted delivery. Carbohyd. Polym. 2022, 279, 119002. [Google Scholar] [CrossRef]
- Zhang, J.K.; Williams, T.D.; Abdallah, M.A.E.; Harrad, S.; Chipman, J.K.; Viant, M.R. Transcriptomic and metabolomic approaches to investigate the molecular responses of human cell lines exposed to the flame retardant hexabromocyclododecane (HBCD). Toxicol. Vitro 2015, 29, 2116–2123. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.-B.; Gou, Z.-Z.; Li, Y.-C.; Zou, L.-H.; Chen, W.-J.; Liu, D.-Y. The efficiency of lemon essential oil-based nanoemulsions on the inhibition of Phomopsis sp. and reduction of postharvest decay of kiwifruit. Foods 2022, 11, 1510. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.P.; Hu, Z.K.; Ye, N.; Yin, H.F. IsoSplitter: Identification and characterization of alternative splicing sites without a reference genome. RNA 2021, 27, 868–875. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.H.; Zhou, X.G.; Deng, Y.C.; Gong, D.X.; Luo, H.F.; Zhu, P. Dissipation behavior, residue distribution, and dietary risk assessment of fluopimomide and dimethomorph in taro using HPLC-MS/MS. Environ. Sci. Pollut. R. 2021, 28, 43956–43969. [Google Scholar] [CrossRef] [PubMed]
- Alessandra, V.; Vincenzo, L.M.; Alessandro, V.; Antonio, D.S. Rectal neuroendocrine cell proliferation in a patient with ulcerative colitis treated with adalimumab. Eur. J. Gastroen. Hepat. 2021, 33, 766–768. [Google Scholar]
- Zheng, Q.; Lin, Z.; Li, X.; Xin, X.; Wu, M.; An, J.; Gui, X.; Li, T.; Pu, H.; Li, H.; et al. Inflammatory cytokine IL6 cooperates with CUDR to aggravate hepatocyte-like stem cells malignant transformation through NF-κB signaling. Sci. Rep. 2016, 6, 36843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.F.; Shui, K.J.; Li, H.Y.; Hu, M.Y.; Zhong, G.H. Antiproliferative effect of azadirachtin A on Spodoptera litura Sl-1 cell line through cell cycle arrest and apoptosis induced by up-regulation of p53. Pestic. Biochem. Phys. 2011, 99, 16–24. [Google Scholar] [CrossRef]
- Wang, X.L.; Qiu, J.; Xu, Y.Y.; Liao, G.Q.; Jia, Q.; Pan, Y.C.; Wang, T.C.; Qian, Y.Z. Integrated non-targeted lipidomics and metabolomics analyses for fluctuations of neonicotinoids imidacloprid and acetamiprid on Neuro-2a cells. Environ. Pollut. 2021, 284, 117327. [Google Scholar] [CrossRef] [PubMed]
- Gallego Romero, I.; Pai, A.A.; Tung, J.; Gilad, Y. RNA-seq: Impact of RNA degradation on transcript quantification. BMC Biol. 2014, 12, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ISO10993-10995; Biological Evaluation of Medical Devices-Part5: Tests for in vitro Cytotoxicity. International Organization for Standardization: Geneva, Switzerland, 2009.
- Fan, H.; Lv, Z.P.; Gan, L.P.; Ning, C.; Li, Z.; Yang, M.H.; Zhang, B.B.; Song, B.C.; Li, G.; Tang, D.Z.; et al. A Novel lncRNA regulates the toll-like receptor signaling pathway and related immune function by stabilizing FOS mRNA as a competitive endogenous RNA. Front. Immunol. 2019, 10, 838. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.-B.; Zhou, L.; Li, J.-J.; Li, Y.-C.; Wang, M.; Zou, L.-H.; Liu, D.-Y.; Chen, W.-J. The combined effect of protein hydrolysis and Lactobacillus plantarum fermentation on antioxidant activity and metabolomic profiles of quinoa beverage. Food Res. Int. 2022, 157, 111416. [Google Scholar] [CrossRef] [PubMed]
- Park, D.W.; Kim, K.G.; Choi, E.A.; Kang, G.R.; Kim, T.S.; Yang, Y.S.; Moon, S.J.; Ha, D.R.; Kim, E.S.; Cho, B.S. Pesticide residues in leafy vegetables, stalk and stem vegetables from South Korea: A long-term study on safety and health risk assessment. Food Addit. Contam. A 2016, 33, 105–118. [Google Scholar] [CrossRef]
- Xu, Z.; Li, L.; Xu, Y.; Wang, S.; Zhang, X.; Tang, T.; Yu, J.; Zhao, H.; Wu, S.; Zhang, C.; et al. Pesticide multi-residues in Dendrobium officinale Kimura et Migo: Method validation, residue levels and dietary exposure risk assessment. Food Chem. 2021, 343, 128490. [Google Scholar] [CrossRef] [PubMed]
- Kwok, M.L.; Meng, Q.; Hu, X.L.; Chung, C.T.; Chan, K.M. Whole-transcriptome sequencing (RNA-seq) study of the ZFL zebrafish liver cell line after acute exposure to Cd2+ ions. Aquat. Toxicol. 2020, 228, 105628. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.Q.; Pei, J.Y.; Li, R.H.; Tian, L.R.; He, X.; Li, Y.P. CD40LG as a prognostic molecular marker regulates tumor microenvironment through immune process in breast cancer. Int. J. Gen. Med. 2021, 14, 8833–8846. [Google Scholar] [CrossRef] [PubMed]
- Sukhatme, V.P.; Cao, X.M.; Chang, L.C.; Tsai-Morris, C.H.; Stamenkovich, D.; Ferreira, P.C.; Cohen, D.R.; Edwards, S.A.; Shows, T.B.; Curran, T.; et al. A zinc finger-encoding gene coregulated with c-fos during growth and differentiation, and after cellular depolarization. Cell 1988, 53, 37–43. [Google Scholar] [CrossRef]
- Wang, B.; Guo, H.F.; Yu, H.Q.; Chen, Y.; Xu, H.Y.; Zhao, G. The role of the transcription factor EGR1 in cancer. Front. Oncol. 2021, 11, 642547. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.G.; Cao, J.T.; Liu, G.Y.; Zhang, H.L.; Liu, X.L. Comparative transcriptome profiling of skeletal muscle from black muscovy duck at different growth stages using RNA-seq. Genes 2020, 11, 1228. [Google Scholar] [CrossRef] [PubMed]
- Oppenheim, J.J. Cytokines: Past, Present, and Future. Int. J. Hematol. 2001, 74, 3–8. [Google Scholar] [CrossRef]
- Ma, Z.; Xin, Z.; Hu, W.; Jiang, S.; Yang, Z.; Yan, X.; Li, X.; Yang, Y.; Chen, F. Forkhead box O proteins: Crucial regulators of cancer EMT. Semin. Cancer Biol. 2018, 50, 21–31. [Google Scholar] [CrossRef]
- Liang, Y.; Xie, S.B.; Wu, C.H.; Hu, Y.; Zhang, Q.; Li, S.; Fan, Y.G.; Leng, R.X.; Pan, H.F.; Xiong, H.B.; et al. Coagulation cascade and complement system in systemic lupus erythematosus. Oncotarget 2018, 9, 14862–14881. [Google Scholar] [CrossRef] [Green Version]
- Schroder, W.A.; Major, L.; Suhrbier, A. The Role of SerpinB2 in Immunity. Crit. Rev. Immunol. 2011, 31, 15–30. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Concentration(ng/μL) | Content (μg) | OD260/280 | OD260/230 | RIN |
|---|---|---|---|---|---|
| H126-1 | 115.00 | 4.03 | 2.05 | 2.11 | 7.40 |
| H126-2 | 112.00 | 3.92 | 2.03 | 2.01 | 7.80 |
| H126-3 | 96.20 | 3.37 | 2.04 | 2.10 | 7.60 |
| M21-1 | 1545.90 | 54.11 | 2.02 | 2.21 | 10.00 |
| M21-2 | 1731.70 | 60.61 | 2.01 | 2.18 | 9.90 |
| M21-3 | 1678.50 | 58.75 | 2.01 | 2.17 | 10.00 |
| L4-1 | 2141.30 | 74.95 | 1.99 | 2.14 | 10.00 |
| L4-2 | 2090.50 | 73.17 | 1.99 | 2.13 | 10.00 |
| L4-3 | 2133.00 | 74.66 | 2.00 | 2.13 | 9.50 |
| Ctr-1 | 2020.60 | 70.72 | 2.00 | 2.15 | 9.90 |
| Ctr-2 | 1862.20 | 65.18 | 2.01 | 2.17 | 9.60 |
| Ctr-3 | 1892.30 | 66.23 | 2.01 | 2.18 | 9.90 |
| Sample | Raw Reads | Clean Reads | Clean Bases | Error Rate (%) | Q20 (%) | Q30 (%) |
|---|---|---|---|---|---|---|
| Ctr_1 | 69321808 | 68648290 | 10248272849 | 0.0247 | 98.15 | 94.41 |
| Ctr_2 | 56231964 | 55657530 | 8295729777 | 0.0249 | 98.07 | 94.21 |
| Ctr_3 | 62512340 | 61926524 | 9209389908 | 0.0245 | 98.25 | 94.67 |
| L4_1 | 65369620 | 64580756 | 9635172686 | 0.0244 | 98.26 | 94.71 |
| L4_2 | 51616818 | 51120046 | 7630992231 | 0.0246 | 98.21 | 94.57 |
| L4_3 | 58748206 | 58167602 | 8680297728 | 0.0249 | 98.08 | 94.18 |
| M21_1 | 52328192 | 51754638 | 7726615557 | 0.0249 | 98.07 | 94.23 |
| M21_2 | 56519460 | 56045588 | 8350478112 | 0.0241 | 98.4 | 95.03 |
| M21_3 | 54012066 | 53454568 | 7983424171 | 0.0248 | 98.1 | 94.31 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.-C.; Liu, S.-Y.; Meng, F.-B.; Xu, S.-H.; Qiu, J.; Qian, Y.-Z.; Xu, Y.-Y.; Li, Y. Comparative Transcriptome Analysis to Investigate the Immunotoxicity Mechanism Triggered by Dimethomorph on Human Jurkat T Cell Lines. Foods 2022, 11, 3848. https://doi.org/10.3390/foods11233848
Li Y-C, Liu S-Y, Meng F-B, Xu S-H, Qiu J, Qian Y-Z, Xu Y-Y, Li Y. Comparative Transcriptome Analysis to Investigate the Immunotoxicity Mechanism Triggered by Dimethomorph on Human Jurkat T Cell Lines. Foods. 2022; 11(23):3848. https://doi.org/10.3390/foods11233848
Chicago/Turabian StyleLi, Yun-Cheng, Shu-Yan Liu, Fan-Bing Meng, Shu-Hui Xu, Jing Qiu, Yong-Zhong Qian, Yan-Yang Xu, and Yun Li. 2022. "Comparative Transcriptome Analysis to Investigate the Immunotoxicity Mechanism Triggered by Dimethomorph on Human Jurkat T Cell Lines" Foods 11, no. 23: 3848. https://doi.org/10.3390/foods11233848
APA StyleLi, Y. -C., Liu, S. -Y., Meng, F. -B., Xu, S. -H., Qiu, J., Qian, Y. -Z., Xu, Y. -Y., & Li, Y. (2022). Comparative Transcriptome Analysis to Investigate the Immunotoxicity Mechanism Triggered by Dimethomorph on Human Jurkat T Cell Lines. Foods, 11(23), 3848. https://doi.org/10.3390/foods11233848