
Protein Characteristics and Bioactivity of Fish Protein Hydrolysates from Tra Catfish (Pangasius hypophthalmus) Side Stream Isolates

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Raw Materials and Sampling
2.1.1. Preparation of Fish Protein Isolates (FPIs)
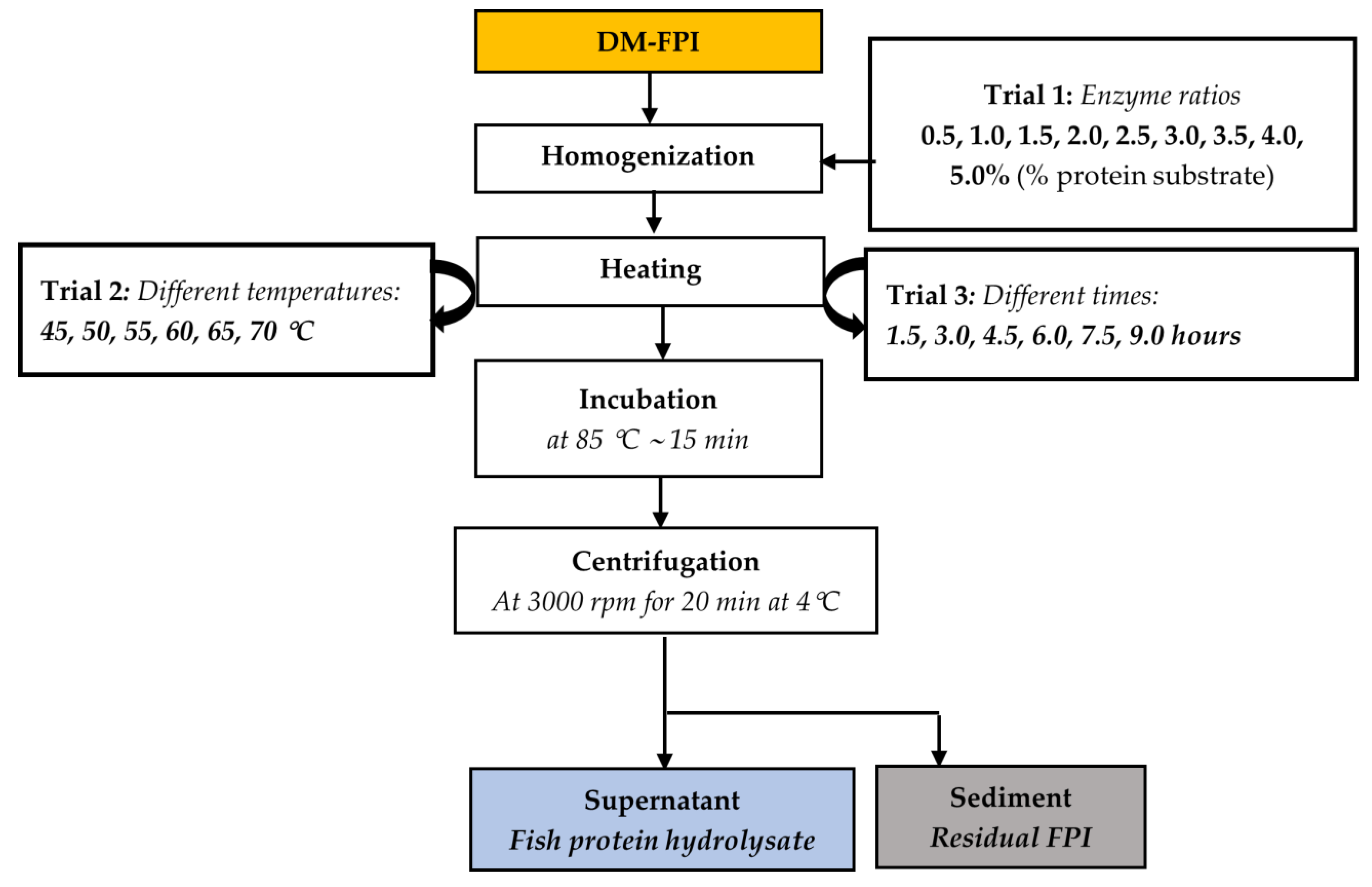
2.1.2. Preparation of Fish Protein Hydrolysate (FPH) from DM-FPI
2.1.3. FPH Preparation from Different FPIs at the Optimal Conditions
2.1.4. Chemicals
2.2. Analyses
2.2.1. Proximate Composition of the Tra Catfish Side-Streams and their Corresponding FPIs
2.2.2. Amino Acid Analysis
2.2.3. Degree of Hydrolysis (DH)
2.2.4. Protein Recovery
2.2.5. Antioxidant Activities
Determination of DPPH-RSA
Determination of TRPC
2.2.6. Statistical Analysis
3. Results and Discussion
3.1. Proximate Composition of the Raw Materials and FPIs
3.2. Optimization of FPH Processing from DM-FPIs
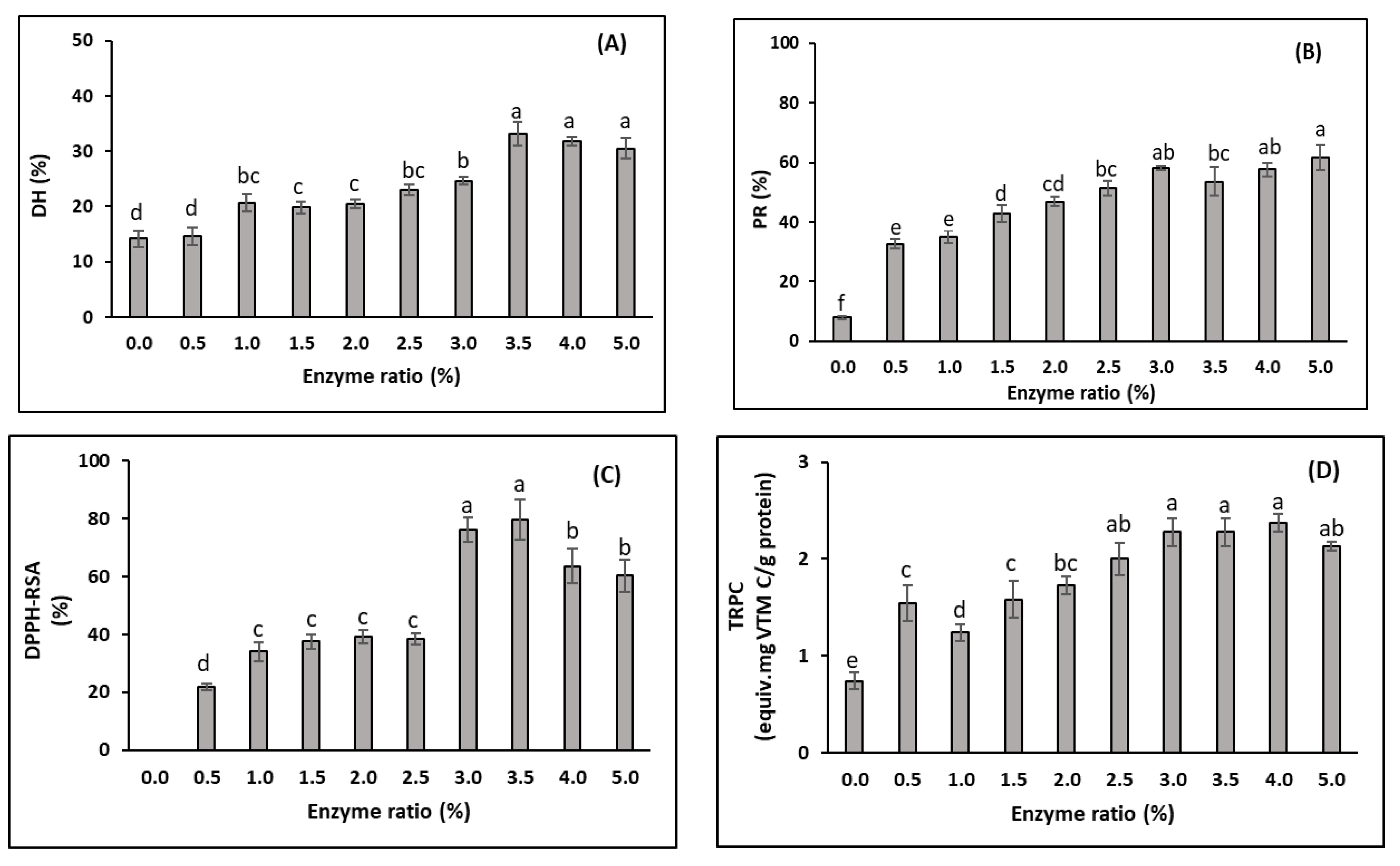
3.2.1. Effects of Enzyme Ratios (Trial 1)
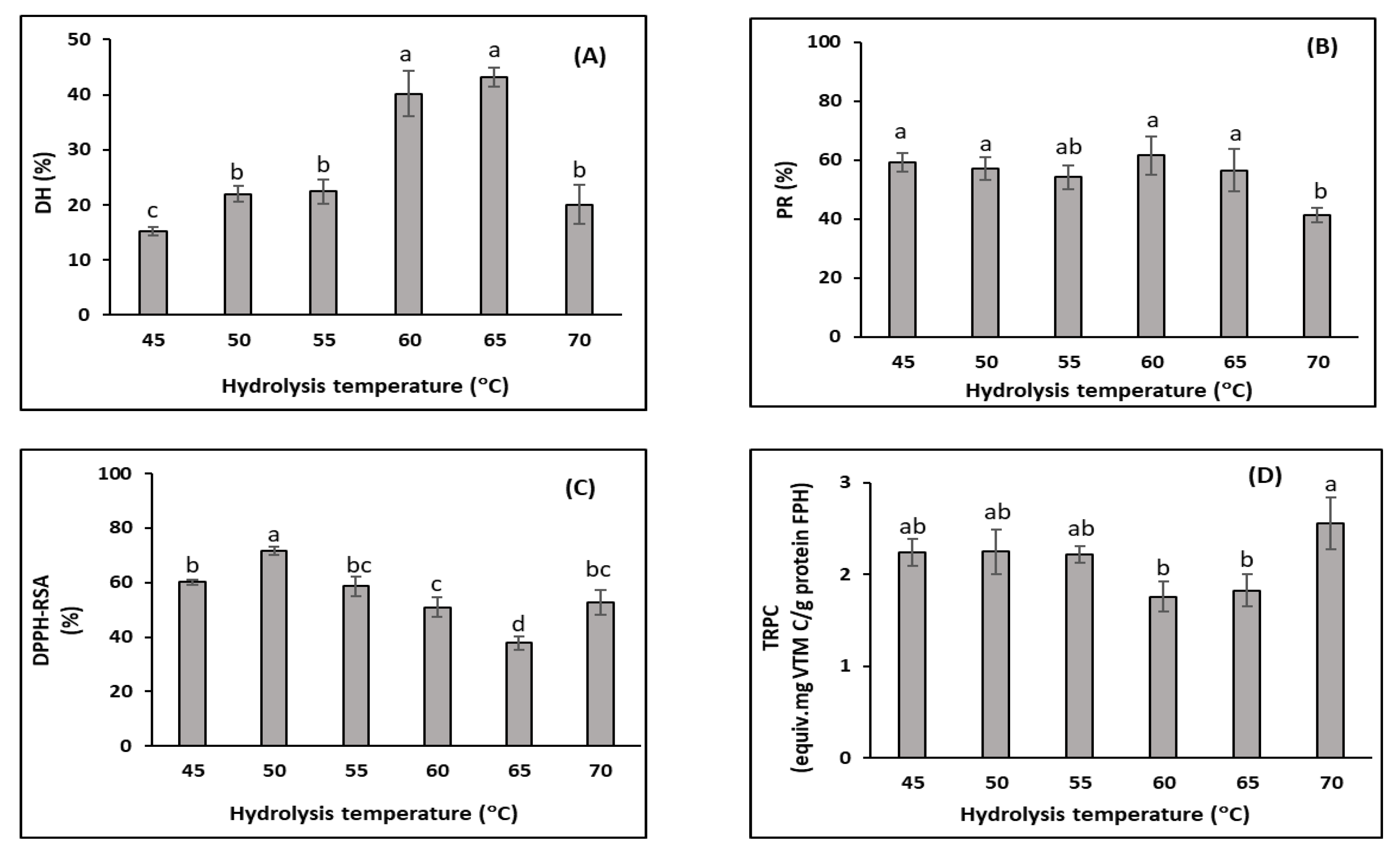
3.2.2. Effects of Hydrolysis Temperature (Trial 2)
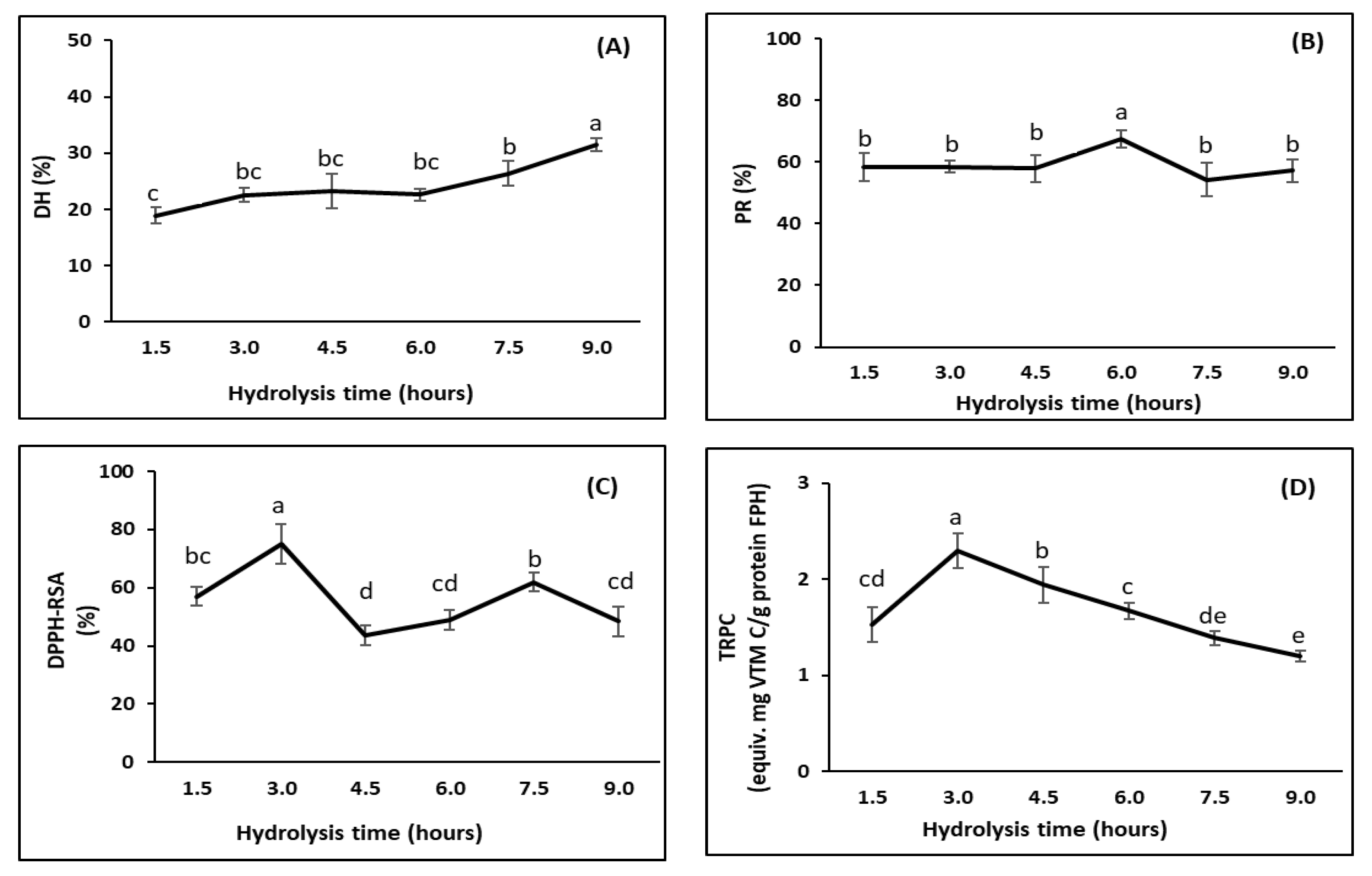
3.2.3. Effects of Hydrolysis Time (Trial 3)
3.3. Comparison of the FPHs Prepared from Different FPIs
3.3.1. Amino Acid Composition
3.3.2. DH, PR, and Antioxidant Activities
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lands, W.E. Fish, Omega-3 and Human Health; AOCS Press: New York, NY, USA, 2005. [Google Scholar]
- Dale, H.F.; Madsen, L.; Lied, G.A. Fish-derived proteins and their potential to improve human health. Nutr. Rev. 2019, 77, 572–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, R.; Yu, Q.; Shen, Y.; Chu, Q.; Chen, G.; Fen, S.; Yang, M.; Yuan, L.; McClements, D.J.; Sun, Q. Production, bioactive properties, and potential applications of fish protein hydrolysates: Developments and challenges. Trends Food Sci. Technol. 2021, 110, 687–699. [Google Scholar] [CrossRef]
- Lees, M.J.; Carson, B.P. The potential role of fish-derived protein hydrolysates on metabolic health, skeletal muscle mass and function in ageing. Nutrients 2020, 12, 2434. [Google Scholar] [CrossRef]
- Henchion, M.; Hayes, M.; Mullen, A.M.; Fenelon, M.; Tiwari, B. Future Protein Supply and Demand: Strategies and Factors Influencing a Sustainable Equilibrium. Foods 2017, 6, 53. [Google Scholar] [CrossRef] [Green Version]
- Shahidi, F.; Ambigaipalan, P. Novel functional food ingredients from marine sources. Curr. Opin. Food Sci. 2015, 2, 123–129. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture 2020. Sustainability in Action; FAO: Rome, Italy, 2020. [Google Scholar]
- Nguyen, H.T.; Hilmarsdóttir, G.S.; Tómasson, T.; Arason, S.; Gudjónsdóttir, M. Changes in Protein and Non-Protein Nitrogen Compounds during Fishmeal Processing—Identification of Unoptimized Processing Steps. Processes 2022, 10, 621. [Google Scholar] [CrossRef]
- Barrett, C.B. Overcoming global food security challenges through science and solidarity. Am. J. Agric. Econ. 2021, 103, 422–447. [Google Scholar] [CrossRef]
- Välimaa, A.-L.; Mäkinen, S.; Mattila, P.; Marnila, P.; Pihlanto, A.; Mäki, M.; Hiidenhovi, J. Fish and fish side streams are valuable sources of high-value components. Food Qual. Saf. 2019, 3, 209–226. [Google Scholar] [CrossRef] [Green Version]
- Galanakis, C.M. Recovery of high added-value components from food wastes: Conventional, emerging technologies and commercialized applications. Trends Food Sci. Technol. 2012, 26, 68–87. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Bao, H.N.D.; Dang, H.T.T.; Tómasson, T.; Arason, S.; Gudjónsdóttir, M. Protein Recovery of Tra Catfish (Pangasius hypophthalmus) Protein-Rich Side Streams by the pH-Shift Method. Foods 2022, 11, 1531. [Google Scholar] [CrossRef]
- Thuy, N.T.; Loc, N.T.; Lindberg, J.; Ogle, B. Survey of the production, processing and nutritive value of catfish by-product meals in the Mekong Delta of Vietnam. Livest. Res. Rur. Dev. 2007, 19, 124. [Google Scholar]
- Nam, P.V.; Van Hoa, N.; Anh, T.T.L.; Trung, T.S. Towards zero-waste recovery of bioactive compounds from catfish (Pangasius hypophthalmus) by-products using an enzymatic method. Waste Biomass Valorization 2020, 11, 4195–4206. [Google Scholar] [CrossRef]
- Dang, H.T.T.; Gudjónsdóttir, M.; Tómasson, T.; Nguyen, M.V.; Karlsdóttir, M.G.; Arason, S. Influence of processing additives, packaging and storage conditions on the physicochemical stability of frozen Tra catfish (Pangasius hypophthalmus) fillets. J. Food Eng. 2018, 238, 148–155. [Google Scholar] [CrossRef]
- Thong, N.T.; Ankamah-Yeboah, I.; Bronnmann, J.; Nielsen, M.; Roth, E.; Schulze-Ehlers, B. Price transmission in the pangasius value chain from Vietnam to Germany. Aquacult. Rep. 2020, 16, 100266. [Google Scholar] [CrossRef]
- Tapal, A.; Tiku, P.K. Chapter 27—Nutritional and Nutraceutical Improvement by Enzymatic Modification of Food Proteins. In Enzymes in Food Biotechnology; Kuddus, M., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 471–481. [Google Scholar]
- Kristinsson, H.G.; Rasco, B.A. Fish protein hydrolysates: Production, biochemical, and functional properties. Crit. Rev. Food Sci. Nutr. 2000, 40, 43–81. [Google Scholar] [CrossRef]
- Caruso, G.; Floris, R.; Serangeli, C.; Di Paola, L. Fishery Wastes as a Yet Undiscovered Treasure from the Sea: Biomolecules Sources, Extraction Methods and Valorization. Mar. Drugs 2020, 18, 622. [Google Scholar] [CrossRef]
- Peighambardoust, S.H.; Karami, Z.; Pateiro, M.; Lorenzo, J.M. A review on health-promoting, biological, and functional aspects of bioactive peptides in food applications. Biomolecules 2021, 11, 631. [Google Scholar] [CrossRef]
- Intarasirisawat, R.; Benjakul, S.; Visessanguan, W.; Wu, J. Effects of skipjack roe protein hydrolysate on properties and oxidative stability of fish emulsion sausage. LWT-Food Sci. Technol. 2014, 58, 280–286. [Google Scholar] [CrossRef]
- Zamora-Sillero, J.; Gharsallaoui, A.; Prentice, C. Peptides from fish by-product protein hydrolysates and its functional properties: An overview. Mar. Biotechnol. 2018, 20, 118–130. [Google Scholar] [CrossRef]
- Atef, M.; Mahdi Ojagh, S. Health benefits and food applications of bioactive compounds from fish byproducts: A review. J. Funct. Foods 2017, 35, 673–681. [Google Scholar] [CrossRef]
- Aluko, R. Amino acids, peptides, and proteins as antioxidants for food preservation. In Handbook of Antioxidants for Food Preservation; Elsevier Inc.: Cambridge, UK, 2015; pp. 105–140. [Google Scholar]
- Ahn, C.-B.; Kim, J.-G.; Je, J.-Y. Purification and antioxidant properties of octapeptide from salmon byproduct protein hydrolysate by gastrointestinal digestion. Food Chem. 2014, 147, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Ahn, C.-B.; Je, J.-Y.; Cho, Y.-S. Antioxidant and anti-inflammatory peptide fraction from salmon byproduct protein hydrolysates by peptic hydrolysis. Int. Food Res. J. 2012, 49, 92–98. [Google Scholar] [CrossRef]
- Walker, P.J.; Winton, J.R. Emerging viral diseases of fish and shrimp. Vet. Res. 2010, 41, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddik, M.A.; Howieson, J.; Fotedar, R.; Partridge, G.J. Enzymatic fish protein hydrolysates in finfish aquaculture: A review. Rev. Aquac. 2021, 13, 406–430. [Google Scholar] [CrossRef]
- Tang, H.G.; Wu, T.X.; Zhao, Z.Y.; Pan, X.D. Effects of fish protein hydrolysate on growth performance and humoral immune response in large yellow croaker (Pseudosciaena crocea R.). J. Zhejiang Univ. Sci. 2008, 9, 684–690. [Google Scholar] [CrossRef] [Green Version]
- Siddik, M.A.B.; Chaklader, M.R.; Foysal, M.J.; Howieson, J.; Fotedar, R.; Gupta, S.K. Influence of fish protein hydrolysate produced from industrial residues on antioxidant activity, cytokine expression and gut microbial communities in juvenile barramundi Lates calcarifer. Fish Shellfish Immunol. 2020, 97, 465–473. [Google Scholar] [CrossRef]
- Halim, N.R.A.; Yusof, H.M.; Sarbon, N.M. Functional and bioactive properties of fish protein hydolysates and peptides: A comprehensive review. Trends Food Sci. Technol. 2016, 51, 24–33. [Google Scholar] [CrossRef]
- Samaranayaka, A.G.; Li-Chan, E.C. Autolysis-assisted production of fish protein hydrolysates with antioxidant properties from Pacific hake (Merluccius productus). Food Chem. 2008, 107, 768–776. [Google Scholar] [CrossRef]
- Kim, S.K.; Perera, U.; Rajapakse, N.; Kim, S. Seafood Processing By-Products; Springer: New York, NY, USA, 2016. [Google Scholar]
- Thiansilakul, Y.; Benjakul, S.; Shahidi, F. Compositions, functional properties and antioxidative activity of protein hydrolysates prepared from round scad (Decapterus maruadsi). Food Chem. 2007, 103, 1385–1394. [Google Scholar] [CrossRef]
- Šližytė, R.; Mozuraitytė, R.; Martínez-Alvarez, O.; Falch, E.; Fouchereau-Peron, M.; Rustad, T. Functional, bioactive and antioxidative properties of hydrolysates obtained from cod (Gadus morhua) backbones. Process Biochem. 2009, 44, 668–677. [Google Scholar] [CrossRef]
- Shafiee, S.; Goli, M.; Khoshkhoo, Z.; Hosseini, S.E. Optimization of hydrolysis conditions (temperature, time, and concentration of alkalase) of rainbow trout viscera using the response surface methodology. J. Food Process. Preserv 2021, 45, e15456. [Google Scholar] [CrossRef]
- Kristinsson, H.G.; Rasco, B.A. Kinetics of the hydrolysis of Atlantic salmon (Salmo salar) muscle proteins by alkaline proteases and a visceral serine protease mixture. Process Biochem. 2000, 36, 131–139. [Google Scholar] [CrossRef]
- Liceaga-Gesualdo, A.; Li-Chan, E. Functional properties of fish protein hydrolysate from herring (Clupea harengus). J. Food Sci. 1999, 64, 1000–1004. [Google Scholar] [CrossRef]
- Dong, S.; Zeng, M.; Wang, D.; Liu, Z.; Zhao, Y.; Yang, H. Antioxidant and biochemical properties of protein hydrolysates prepared from Silver carp (Hypophthalmichthys molitrix). Food Chem. 2008, 107, 1485–1493. [Google Scholar] [CrossRef]
- Undeland, I.; Kristinsson, H.G.; Hultin, H.O. Hemoglobin-mediated oxidation of washed minced cod muscle phospholipids: Effect of pH and hemoglobin source. J. Agric. Food Chem. 2004, 52, 4444–4451. [Google Scholar] [CrossRef]
- Hultin, H.; Kristinsson, H.; Lanier, T.; Park, J. Process for recovery of functional proteins by pH shifts. In Surimi and Surimi Seafood; Park, J.W., Ed.; CRC Press: Boca Raton, FL, USA, 2005; pp. 107–139. [Google Scholar]
- Abdollahi, M.; Marmon, S.; Chaijan, M.; Undeland, I. Tuning the pH-shift protein-isolation method for maximum hemoglobin-removal from blood rich fish muscle. Food Chem. 2016, 212, 213–224. [Google Scholar] [CrossRef]
- Intarasirisawat, R.; Benjakul, S.; Visessanguan, W.; Wu, J. Antioxidative and functional properties of protein hydrolysate from defatted skipjack (Katsuwonous pelamis) roe. Food Chem. 2012, 135, 3039–3048. [Google Scholar] [CrossRef]
- Klompong, V.; Benjakul, S.; Kantachote, D.; Shahidi, F. Antioxidative activity and functional properties of protein hydrolysate of yellow stripe trevally (Selaroides leptolepis) as influenced by the degree of hydrolysis and enzyme type. Food Chem. 2007, 102, 1317–1327. [Google Scholar] [CrossRef]
- Mackie, I.M. General review of fish protein hydrolysates. Anim. Feed Sci. Technol. 1982, 7, 113–124. [Google Scholar] [CrossRef]
- Hultin, H.O.; Kelleher, S.D. High Efficiency Alkaline Protein Extraction. U.S. Patent 6136959A, 24 October 2000. [Google Scholar]
- Kakko, T.; Damerau, A.; Nisov, A.; Puganen, A.; Tuomasjukka, S.; Honkapää, K.; Tarvainen, M.; Yang, B. Quality of Protein Isolates and Hydrolysates from Baltic Herring (Clupea harengus membras) and Roach (Rutilus rutilus) Produced by pH-Shift Processes and Enzymatic Hydrolysis. Foods 2022, 11, 230. [Google Scholar] [CrossRef]
- Nisov, A.; Kakko, T.; Alakomi, H.L.; Lantto, R.; Honkapää, K. Comparison of enzymatic and pH shift methods to extract protein from whole Baltic herring (Clupea harengus membras) and roach (Rutilus rutilus). Food Chem. 2022, 373, 131524. [Google Scholar] [CrossRef] [PubMed]
- ISO 6496:1999; Animal Feeding Stuffs: Determination of Moisture and Other Volatile Matter Content. International Organization for Standarddization: Geneva, Switzerland, 1999.
- ISO 5983-2:2009. Determination of Nitrogen Content and Calculation of Crude Protein Content—Part 2: Block Digestion and Steam Distillation Method. International Organization for Standardization: Geneva, Switzerland, 2009.
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar]
- AOAC. Official Methods of Analysis: Official Method for Ash. In Method No. 936.03; Association of Official Analytical Chemists: Washington, DC, USA, 2000. [Google Scholar]
- ISO 13903:2005; Animal Feeding Stuffs: Determination of Amino acids Content. International Organization for Standardization: Geneva, Switzerland, 2005.
- Goodwin, J.F. The colorimetric estimation of plasma amino nitrogen with DNFB. Clin. Chem. 1968, 14, 1080–1090. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.M.; da Fonseca, R.d.S.; Prentice, C. Comparing the hydrolysis degree of industrialization byproducts of Withemouth croaker (Micropogonias furnieri) using microbial enzymes. Int. Food Res. J. 2014, 21, 1757–1761. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Mæhre, H.K.; Dalheim, L.; Edvinsen, G.K.; Elvevoll, E.O.; Jensen, I.-J. Protein determination—Method matters. Foods 2018, 7, 5. [Google Scholar] [CrossRef] [Green Version]
- Fu, H.Y.; Shieh, D.E.; Ho, C.T. Antioxidant and free radical scavenging activities of edible mushrooms. J. Food Lipids 2002, 9, 35–43. [Google Scholar] [CrossRef]
- Oyaizu, M. Studies on products of browning reaction antioxidative activities of products of browning reaction prepared from glucosamine. Jpn. J. Nutr. 1986, 44, 307–315. [Google Scholar] [CrossRef] [Green Version]
- Kristinsson, H.G.; Liang, Y. Effect of pH-shift processing and surimi processing on Atlantic croaker (Micropogonias undulates) muscle proteins. J. Food Sci. 2006, 71, C304–C312. [Google Scholar] [CrossRef]
- Gehring, C.K.; Gigliotti, J.C.; Moritz, J.S.; Tou, J.C.; Jaczynski, J. Functional and nutritional characteristics of proteins and lipids recovered by isoelectric processing of fish by-products and low-value fish: A review. Food Chem. 2011, 124, 422–431. [Google Scholar] [CrossRef]
- Halldorsdottir, S.M.; Sveinsdottir, H.; Freysdottir, J.; Kristinsson, H.G. Oxidative processes during enzymatic hydrolysis of cod protein and their influence on antioxidant and immunomodulating ability. Food Chem. 2014, 142, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Khantaphant, S.; Benjakul, S.; Ghomi, M.R. The effects of pretreatments on antioxidative activities of protein hydrolysate from the muscle of brownstripe red snapper (Lutjanus vitta). LWT-Food Sci. Technol. 2011, 44, 1139–1148. [Google Scholar] [CrossRef]
- Halldórsdóttir, S.M.; Hamaguchi, P.Y.; Sveinsdóttir, H.; Kristinsson, H.G.; Bergsson, A.B.; Thorkelsson, G. Properties of hydrolysed saithe protein isolates prepared via pH shift process with and without dewatering. LWT-Food Sci. Technol. 2011, 44, 1999–2004. [Google Scholar] [CrossRef]
- Benjakul, S.; Morrissey, M.T. Protein hydrolysates from Pacific whiting solid wastes. J. Agric. Food Chem. 1997, 45, 3423–3430. [Google Scholar] [CrossRef]
- Wei, C.-K.; Thakur, K.; Liu, D.-H.; Zhang, J.-G.; Wei, Z.-J. Enzymatic hydrolysis of flaxseed (Linum usitatissimum L.) protein and sensory characterization of Maillard reaction products. Food Chem. 2018, 263, 186–193. [Google Scholar] [CrossRef]
- Tacias-Pascacio, V.G.; Morellon-Sterling, R.; Siar, E.-H.; Tavano, O.; Berenguer-Murcia, Á.; Fernandez-Lafuente, R. Use of Alcalase in the production of bioactive peptides: A review. Int. J. Biol. Macromol. 2020, 165, 2143–2196. [Google Scholar] [CrossRef]
- Nothling, M.D.; Xiao, Z.; Bhaskaran, A.; Blyth, M.T.; Bennett, C.W.; Coote, M.L.; Connal, L.A. Synthetic catalysts inspired by hydrolytic enzymes. ACS Catal. 2018, 9, 168–187. [Google Scholar] [CrossRef]
- Guerard, F.; Guimas, L.; Binet, A. Production of tuna waste hydrolysates by a commercial neutral protease preparation. J. Mol. Catal. B Enzym. 2002, 19–20, 489–498. [Google Scholar] [CrossRef]
- Noman, A.; Xu, Y.; Al-Bukhaiti, W.Q.; Abed, S.M.; Ali, A.H.; Ramadhan, A.H.; Xia, W. Influence of enzymatic hydrolysis conditions on the degree of hydrolysis and functional properties of protein hydrolysate obtained from Chinese sturgeon (Acipenser sinensis) by using papain enzyme. Process Biochem. 2018, 67, 19–28. [Google Scholar] [CrossRef]
- Najafian, L.; Babji, A.S. Production of bioactive peptides using enzymatic hydrolysis and identification antioxidative peptides from patin (Pangasius sutchi) sarcoplasmic protein hydolysate. J. Funct. Foods 2014, 9, 280–289. [Google Scholar] [CrossRef]
- Chalamaiah, M.; Hemalatha, R.; Jyothirmayi, T. Fish protein hydrolysates: Proximate composition, amino acid composition, antioxidant activities and applications: A review. Food Chem. 2012, 135, 3020–3038. [Google Scholar] [CrossRef] [PubMed]
- Tanuja, S.; Haridas, A.; Zynudheen, A.; Joshy, C. Functional and antioxidative properties of fish protein hydrolysate (FPH) produced from the frame meat of striped catfish Pangasianodon hypophthalmus (Sauvage, 1878) using alkaline protease alcalase. Indian J. Fish 2014, 61, 82–89. [Google Scholar]
- Eisenthal, R.; Peterson, M.E.; Daniel, R.M.; Danson, M.J. The thermal behaviour of enzyme activity: Implications for biotechnology. Trends Biotechnol. 2006, 24, 289–292. [Google Scholar] [CrossRef]
- Daniel, R.M.; Danson, M.J. A new understanding of how temperature affects the catalytic activity of enzymes. Trends Biochem. Sci. 2010, 35, 584–591. [Google Scholar] [CrossRef] [PubMed]
- Peterson, M.E.; Daniel, R.M.; Danson, M.J.; Eisenthal, R. The dependence of enzyme activity on temperature: Determination and validation of parameters. Biochem. J. 2007, 402, 331–337. [Google Scholar] [CrossRef]
- Amiza, M.; Ow, Y.; Faazaz, A. Physicochemical properties of silver catfish (Pangasius sp.) frame hydrolysate. Int. Food Res. J. 2013, 20, 1255. [Google Scholar]
- Sabeena Farvin, K.H.; Andersen, L.L.; Otte, J.; Nielsen, H.H.; Jessen, F.; Jacobsen, C. Antioxidant activity of cod (Gadus morhua) protein hydrolysates: Fractionation and characterisation of peptide fractions. Food Chem. 2016, 204, 409–419. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.-C.; Chen, H.-M.; Shiau, C.-Y. Free amino acids and peptides as related to antioxidant properties in protein hydrolysates of mackerel (Scomber austriasicus). Int. Food Res. J. 2003, 36, 949–957. [Google Scholar] [CrossRef]
- Wang, X.; Yu, H.; Xing, R.; Chen, X.; Liu, S.; Li, P. Optimization of the extraction and stability of antioxidative peptides from mackerel (Pneumatophorus japonicus) protein. Biomed Res. Int. 2017, 2017, 6837285. [Google Scholar] [CrossRef] [Green Version]
- Amiza, M.; Nurul Ashikin, S.; Faazaz, A. Optimization of enzymatic protein hydrolysis from silver catfish (Pangasius sp.) frame. Int. Food Res. J. 2011, 18, 775–781. [Google Scholar]
- Wasswa, J.; Tang, J.; GU, X.H. Optimization of the production of hydrolysates from grass carp (Ctenopharyngodon idella) skin using alcalase. J. Food Biochem. 2008, 32, 460–473. [Google Scholar] [CrossRef]
- Vázquez, J.A.; Blanco, M.; Massa, A.E.; Amado, I.R.; Pérez-Martín, R.I. Production of fish protein hydrolysates from Scyliorhinus canicula discards with antihypertensive and antioxidant activities by enzymatic hydrolysis and mathematical optimization using response surface methodology. Mar. Drugs 2017, 15, 306. [Google Scholar] [CrossRef] [PubMed]
- Normah, I.; Jamilah, B.; Saari, N.; Yaakob, B.C.M. Optimization of hydrolysis conditions for the production of threadfin bream (Nemipterus japonicus) hydrolysate by Alcalase. J. Muscle Foods 2005, 16, 87–102. [Google Scholar] [CrossRef]
- Chi, C.-F.; Hu, F.-Y.; Wang, B.; Li, Z.-R.; Luo, H.-Y. Influence of amino acid compositions and peptide profiles on antioxidant capacities of two protein hydrolysates from skipjack tuna (Katsuwonus pelamis) dark muscle. Mar. Drugs 2015, 13, 2580–2601. [Google Scholar] [CrossRef]
- Bhaskar, N.; Benila, T.; Radha, C.; Lalitha, R.G. Optimization of enzymatic hydrolysis of visceral waste proteins of Catla (Catla catla) for preparing protein hydrolysate using a commercial protease. Bioresour. Technol. 2008, 99, 335–343. [Google Scholar] [CrossRef]
- FAO. Protein and Amino acid Requirements in Human Nutrition: Report of a Joint FAO/WHO/UNU Expert Consultation; No. 935; WHO: Geneva, Switzerland, 2007. [Google Scholar]
- Damgaard, T.; Lametsch, R.; Otte, J. Antioxidant capacity of hydrolyzed animal by-products and relation to amino acid composition and peptide size distribution. J. Food Sci. Technol. 2015, 52, 6511–6519. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Water Content | Crude Protein | Lipid Content | Ash Content | ||
|---|---|---|---|---|---|
| Raw material | Dark muscle | 66.5 ± 1.0 A | 14.7 ± 0.2 B | 17.6 ± 1.5 B | 0.8 ± 0.1 B |
| HBB | 54.8 ± 1.0 C | 15.2 ± 0.1 A | 21.9 ± 0.6 A | 7.8 ± 0.4 A | |
| ACO | 60.8 ± 2.0 B | 15.3 ± 0.1 A | 23.5 ± 1.1 A | 1.0 ± 0.1 B | |
| FPIs | DM-FPI | 73.9 ± 0.7 ab | 23.5 ± 0.9 a | 3.2 ± 0.0 a | 0.1 ± 0.0 a |
| HBB-FPI | 77.3 ± 0.8 a | 20.4 ± 0.5 b | 2.8 ± 0.4 a | 0.1 ± 0.0 a | |
| ACO-FPI | 73.1 ± 2.1 b | 24.4 ± 1.4 a | 3.1 ± 0.1 a | 0.0 ± 0.0 a |
| Amino Acids | DM-FPI ** | HBB-FPI ** | ACO-FPI ** | DM-FPH | HBB-FPH | ACO-FPH | FAO/WHO/UNU * |
|---|---|---|---|---|---|---|---|
| 4-Hydroxyproline | ND | ND | 0.6 | ND | ND | ND | |
| Alanine b | 6.3 | 6.5 | 6.4 | 6.6 | 6.5 | 6.6 | |
| Arginie a | 6.6 | 6.9 | 6.3 | 6.4 | 5.9 | 6.6 | |
| Aspartic acid | 11.2 | 12.1 | 10.5 | 10.7 | 10.7 | 10.9 | |
| Cystein | ND | ND | ND | 0.6 | 0.7 | 0.6 | |
| Cystine | 1.1 | 1.1 | 1.0 | 0.7 | 0.6 | 0.7 | |
| Gamma-aminobutyric acid | ND | ND | ND | 0.5 | 0.6 | 0.6 | |
| Glutamic acid | 18.4 | 19.4 | 17.2 | 20.4 | 19.2 | 20.4 | |
| Glycine b | 3.7 | 3.9 | 5.1 | 3.3 | 3.4 | 3.4 | |
| Histidine a | 2.6 | 2.6 | 2.8 | 2.4 | 2.5 | 2.2 | 1.5 |
| Isoleucine ab | 4.7 | 5.0 | 4.6 | 4.0 | 4.2 | 3.9 | 3.0 |
| Leucine ab | 8.8 | 9.1 | 7.9 | 8.5 | 8.8 | 8.5 | 5.9 |
| Lysine a | 9.3 | 10.1 | 8.8 | 10.2 | 9.6 | 10.4 | 4.5 |
| Methionine ab | 3.3 | 3.5 | 2.9 | 2.8 | 2.8 | 3.2 | 2.2 |
| Phenylalanine ab | 3.8 | 4.2 | 3.8 | 3.1 | 3.1 | 2.9 | 3.8 |
| Proline b | 4.6 | 4.3 | 5.0 | 3.6 | 4.0 | 3.4 | |
| Serine | 4.2 | 4.4 | 4.1 | 4.5 | 4.8 | 4.6 | |
| Threonine a | 4.6 | 4.9 | 4.2 | 4.5 | 4.5 | 4.4 | 2.3 |
| Tyrosine | 2.2 | 3.5 | 3.1 | 2.6 | 2.8 | 2.7 | |
| Valine ab | 5.0 | 5.3 | 4.8 | 4.3 | 4.8 | 4.4 | 3.9 |
| Total amino acids | 100.5 | 106.8 | 99.1 | 99.6 | 99.5 | 100.2 | |
| Total essential amino acids | 48.8 | 51.7 | 46.2 | 46.2 | 46.3 | 46.4 | |
| Total hydrophobic amino acid | 40.3 | 41.8 | 40.6 | 36.3 | 37.6 | 36.2 |
| Production | DH (%) | PR (%) | DPPH (%) | TRPC (Equiv. mg Vitamin C/g FPH Protein) |
|---|---|---|---|---|
| DM-FPH | 22.5 ± 1.3 a | 58.4 ± 2.0 b | 75.1 ± 6.8 a | 2.3 ± 0.2 b |
| HBB-FPH | 22.9 ± 1.6 a | 60.4 ± 5.7 b | 57.9 ± 3.6 b | 2.0 ± 0.2 b |
| ACO-FPH | 24.0 ± 3.5 a | 81.5 ± 4.3 a | 86.1 ± 1.6 a | 6.4 ± 0.4 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, H.T.; Bao, H.N.D.; Dang, H.T.T.; Tómasson, T.; Arason, S.; Gudjónsdóttir, M. Protein Characteristics and Bioactivity of Fish Protein Hydrolysates from Tra Catfish (Pangasius hypophthalmus) Side Stream Isolates. Foods 2022, 11, 4102. https://doi.org/10.3390/foods11244102
Nguyen HT, Bao HND, Dang HTT, Tómasson T, Arason S, Gudjónsdóttir M. Protein Characteristics and Bioactivity of Fish Protein Hydrolysates from Tra Catfish (Pangasius hypophthalmus) Side Stream Isolates. Foods. 2022; 11(24):4102. https://doi.org/10.3390/foods11244102
Chicago/Turabian StyleNguyen, Hang Thi, Huynh Nguyen Duy Bao, Huong Thi Thu Dang, Tumi Tómasson, Sigurjón Arason, and María Gudjónsdóttir. 2022. "Protein Characteristics and Bioactivity of Fish Protein Hydrolysates from Tra Catfish (Pangasius hypophthalmus) Side Stream Isolates" Foods 11, no. 24: 4102. https://doi.org/10.3390/foods11244102
APA StyleNguyen, H. T., Bao, H. N. D., Dang, H. T. T., Tómasson, T., Arason, S., & Gudjónsdóttir, M. (2022). Protein Characteristics and Bioactivity of Fish Protein Hydrolysates from Tra Catfish (Pangasius hypophthalmus) Side Stream Isolates. Foods, 11(24), 4102. https://doi.org/10.3390/foods11244102