
Bioconversion of Lactose into Glucose–Galactose Syrup by Two-Stage Enzymatic Hydrolysis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Materials
2.2. Analyses of Whey Permeate
2.3. Production of GG Syrup
2.3.1. Hydrolysis of Permeate by β-Galactosidase
2.3.2. Isomerisation of Glucose
2.4. Chromatographic Determination of Carbohydrates
2.5. Data Analysis
3. Results and Discussion
3.1. Whey Permeate Composition
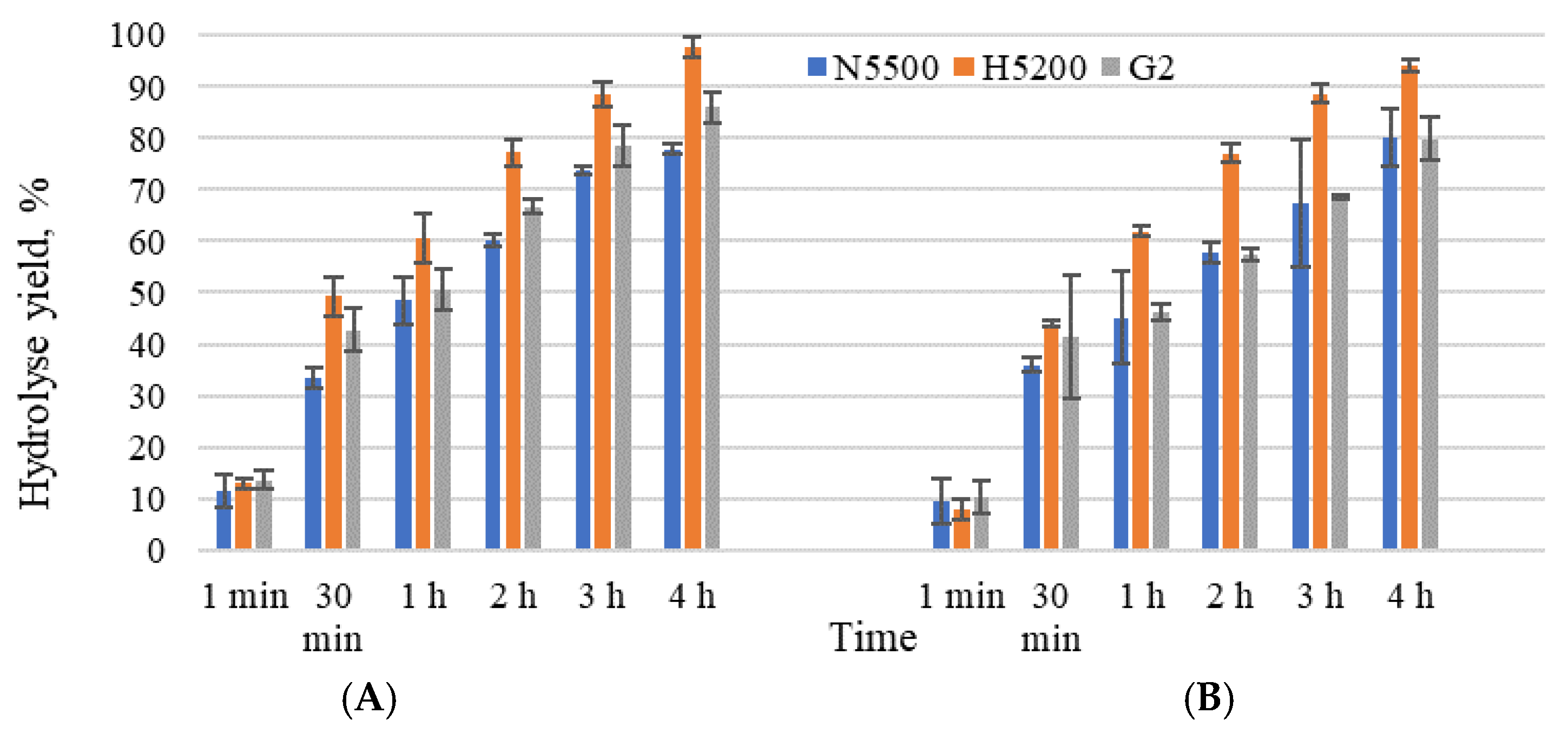
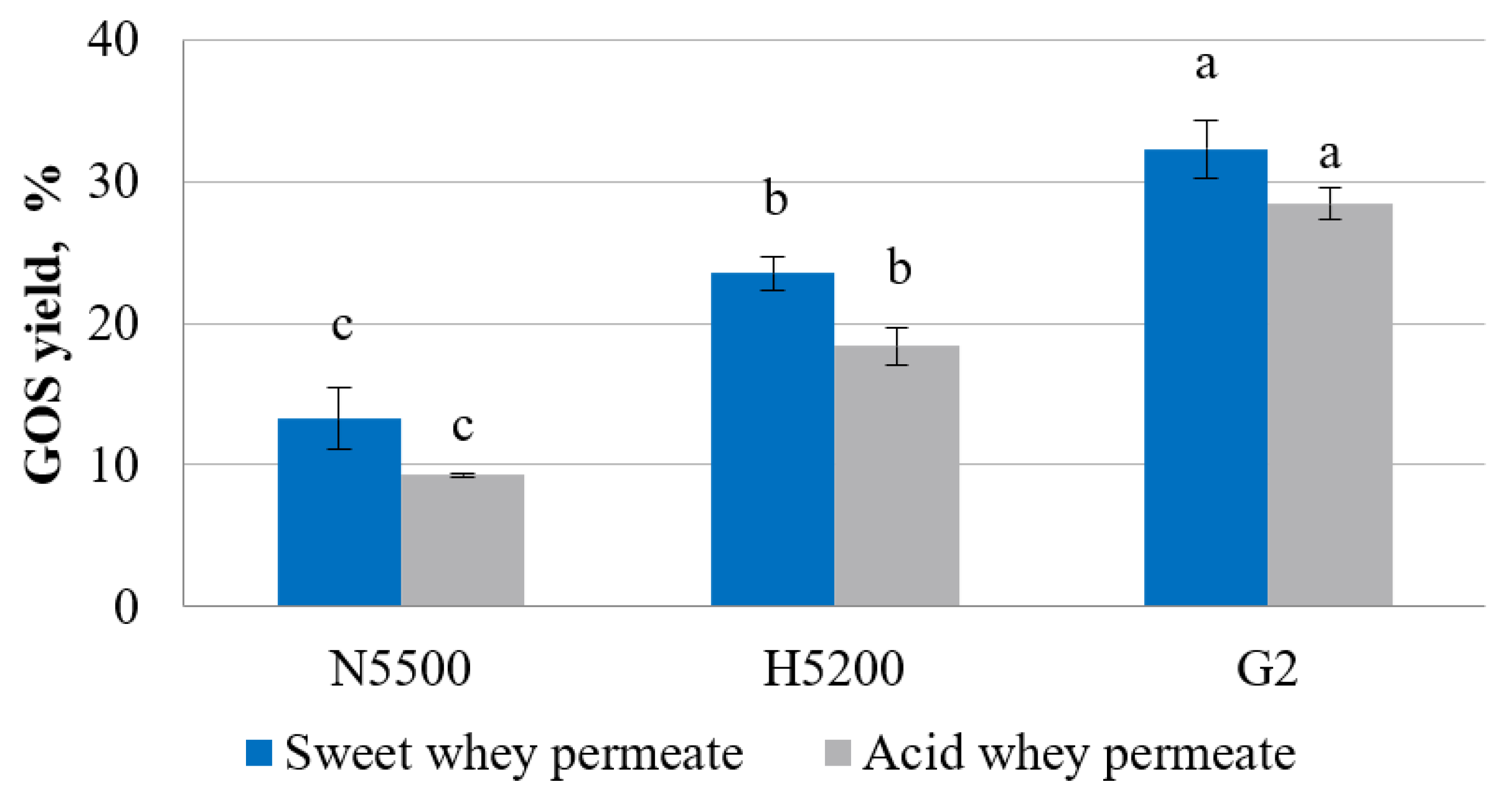
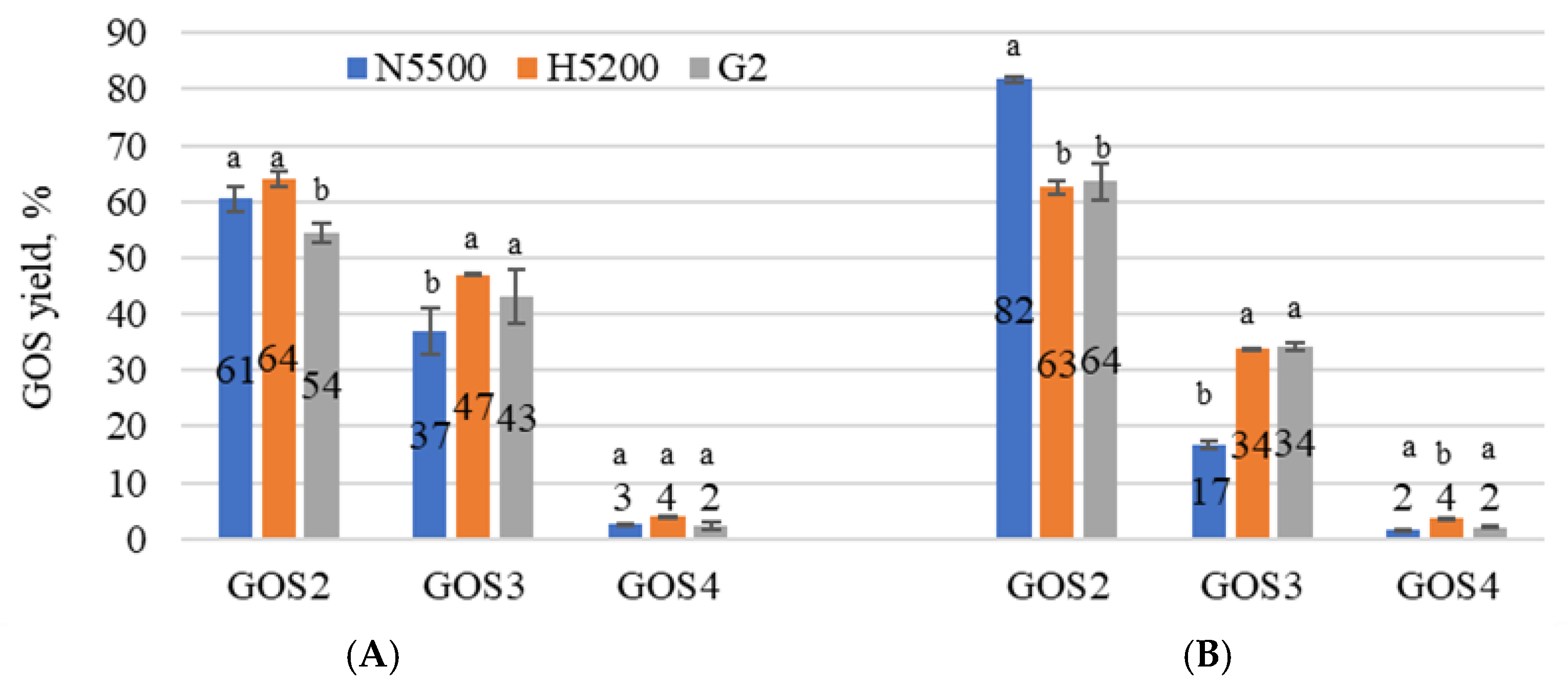
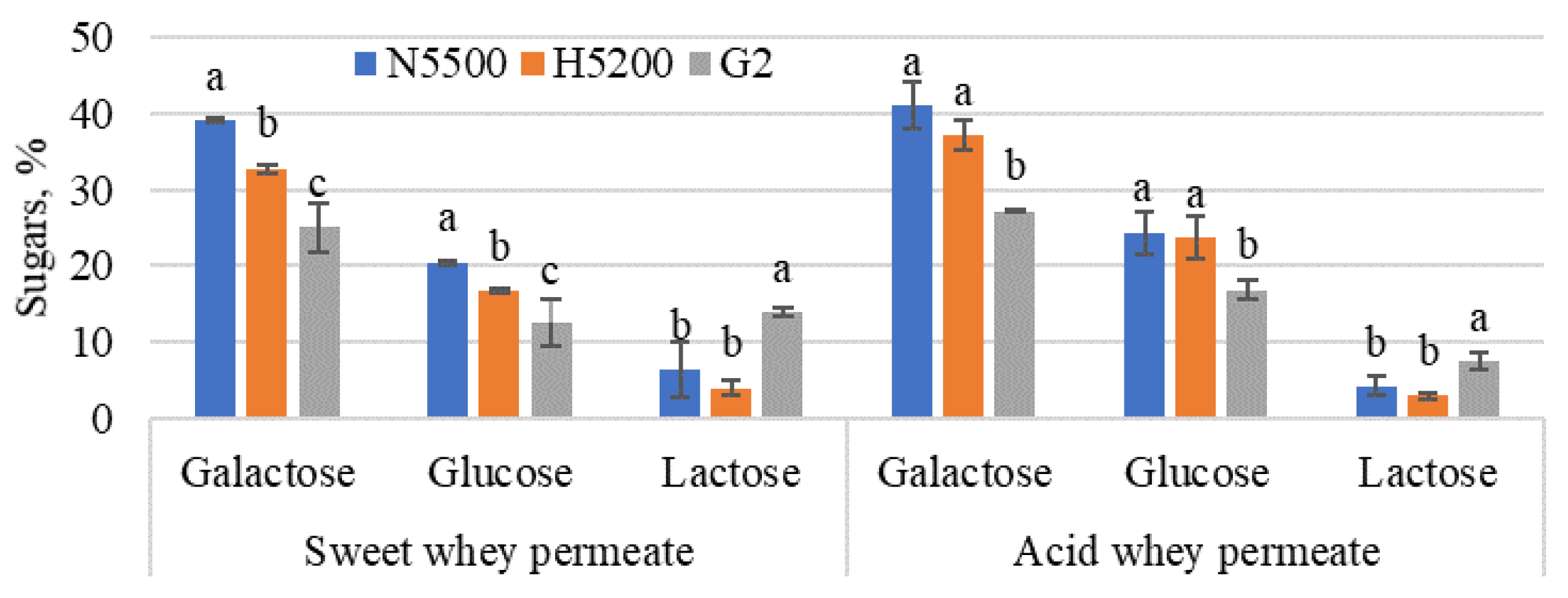
3.2. Enzymatic Hydrolysis and Transgalactosylation in Whey Permeate
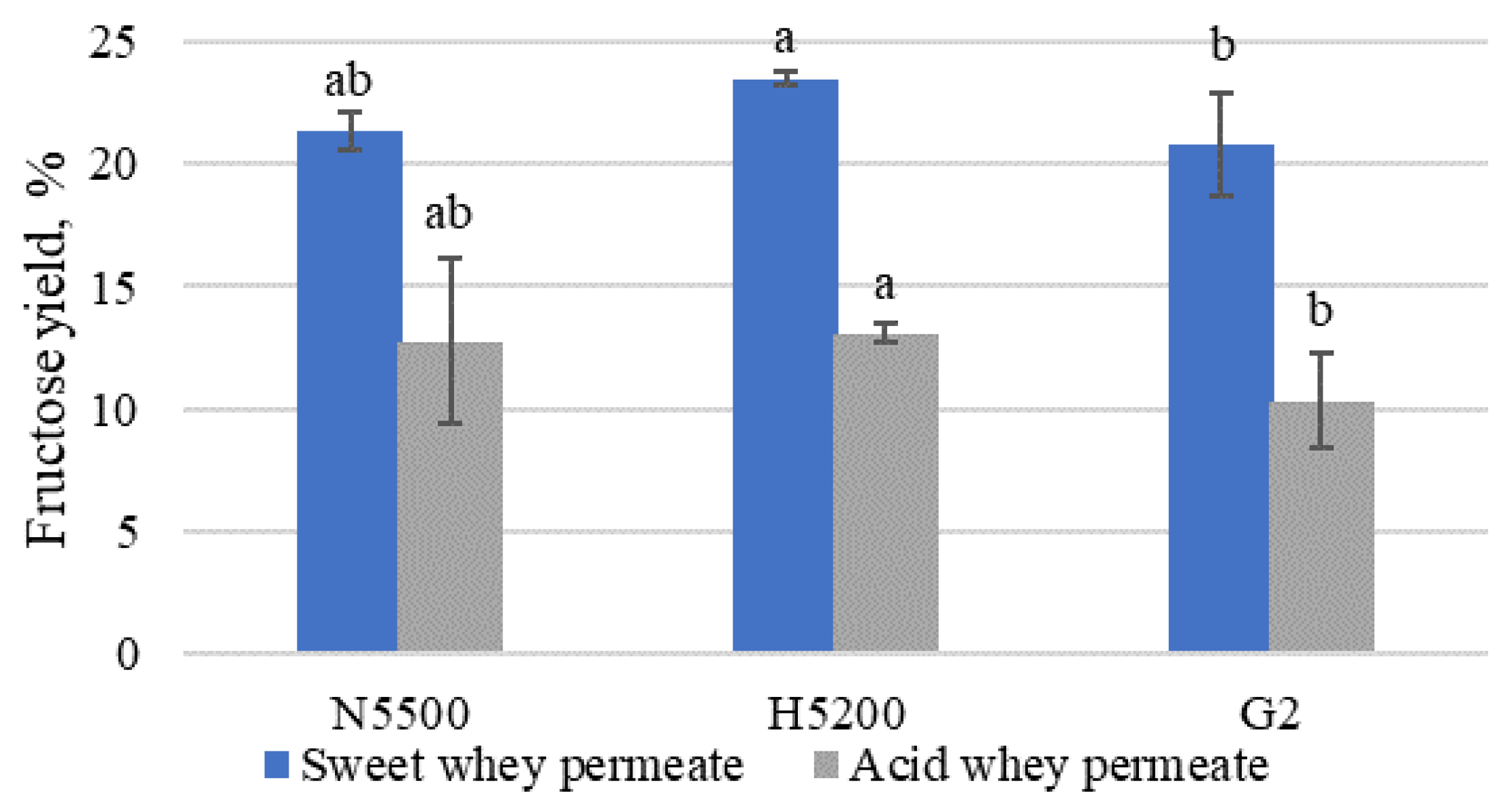
3.3. Effect of Two-Stage Enzymatic Hydrolysis to Increase Syrup Sweetness
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Torres, P.; Batista-Viera, F. Production of d-tagatose and d-fructose from whey by co-immobilized enzymatic system. Mol. Catal. 2019, 463, 99–109. [Google Scholar] [CrossRef]
- Lindsay, M.J.; Huang, K.; Buchinger, B.A.; Maravelias, C.T.; Dumesic, J.A.; Rankin, S.A.; Huber, G.W. Catalytic production of glucose–galactose syrup from greek yogurt acid whey in a continuous-flow reactor. ChemSusChem 2020, 13, 791–802. [Google Scholar] [CrossRef] [PubMed]
- Ureta, M.M.; Martins, G.N.; Figueira, O.; Pires, P.F.; Castilho, P.C.; Gomez-Zavaglia, A. Recent advances in β-galactosidase and fructosyltransferase immobilization technology. Crit. Rev. 2020, 61, 2659–2690. [Google Scholar] [CrossRef] [PubMed]
- Seok, Y.; Uk, H.; Park, C.; Wook, S. Batch and continuous synthesis of lactulose from whey lactose by immobilized b-galactosidase. Food Chem. 2013, 136, 689–694. [Google Scholar] [CrossRef]
- Lindsay, M.J.; Walker, T.W.; Dumesic, J.A.; Rankin, S.A.; Huber, G.W. Production of monosaccharides and whey protein from acid whey waste streams in the dairy industry. Green Chem. 2018, 20, 1824–1834. [Google Scholar] [CrossRef]
- Whintaker, J.R.; Voragen, A.G.J.; Wong, D.W.S. Handbook of Food Enzymology; Marcel Dekker: New York, NY, USA, 2003. [Google Scholar] [CrossRef]
- Mano, M.C.R.; Paulino, B.N.; Pastore, G.M. Whey permeate as the raw material in galacto-oligosaccharide synthesis using commercial enzymes. Food Res. J. 2019, 124, 78–85. [Google Scholar] [CrossRef]
- Kruschitz, A.; Nidetzky, B. Downstream processing technologies in the biocatalytic production of oligosaccharides. Biotechnol. Adv. 2020, 43, 107568. [Google Scholar] [CrossRef]
- Araya, E.; Urrutia, P.; Romero, O.; Illanes, A.; Wilson, L. Design of combined crosslinked enzyme aggregates (combi-CLEAs) of β-galactosidase and glucose isomerase for the one-pot production of fructose syrup from lactose. Food Chem. 2019, 288, 102–107. [Google Scholar] [CrossRef]
- Lee, D.G.; Choi, D.J.; Park, J.K. Ketoisomeric conversion of glucose derived from microalgal biomasses. Process Biochem. 2015, 50, 941–947. [Google Scholar] [CrossRef]
- Dehkordi, A.M.; Tehrany, M.S.; Safari, I. Kinetics of glucose isomerization to fructose by immobilized glucose isomerase (Sweetzyme IT). Ind. Eng. Chem. Res. 2009, 48, 3271–3278. [Google Scholar] [CrossRef]
- Yu, H.; Guo, Y.; Wu, D.; Zhan, W.; Lu, G. Immobilization of glucose isomerase onto GAMM support for isomerization of glucose to fructose. J. Mol. Catal. B Enzym. 2011, 72, 73–76. [Google Scholar] [CrossRef]
- Wang, Y.; Pan, Y.; Zhang, Z.; Sun, R.; Fang, X.; Yu, D. Combination use of ultrasound irradiation and ionic liquid in enzymatic isomerization of glucose to fructose. Process Biochem. 2012, 47, 976–982. [Google Scholar] [CrossRef]
- ISO 8070:2007; Milk and Milk Products—Determination of Calcium, Sodium, Potassium and Magnesium Contents—Atomic Absorption Spectrometric Method. International Organization of Standardization: Geneva, Switzerland, 2007.
- ISO 9874:2006; Milk—Determination of Total Phosphorus Content—Method Using Molecular Absorption Spectrometry. International Organization of Standardization: Geneva, Switzerland, 2006.
- Zolnere, K.; Ciproviča, I. Lactose hydrolysis in different solids content whey and milk permeates. In Proceedings of the 13th Baltic Conference on Food Science and Technology, FOODBALT 2019 Joined with 5th North and East European Congress on Food, NEEFood 2019 “Food. Nutrition. Well-Being”, Jelgava, Latvia, 2–3 May 2019; pp. 35–39. [Google Scholar] [CrossRef]
- Majore, K.; Ciproviča, I. Optimisation of Lactose Hydrolysis by Combining Solids and ß-Galactosidase Concentrations in Whey Permeates. Proc. Latv. Acad. Sciences. Sect. B. Nat. Exact Appl. Sci. 2020, 74, 263–269. [Google Scholar] [CrossRef]
- Gaily, M.H.; Elhassan, B.M.; Abasaeed, A.E.; Al-Shrhan, M. Isomerization and Kinetics of Glucose into Fructose. Int. J. Eng. Technol. 2010, 10, 1–5. [Google Scholar]
- Lee, C.H.; Kim, H.T.; Yun, E.J.; Lee, A.R.; Kim, S.R.; Kim, J.H.; Choi, I.G.; Kim, K.H. A novel agarolytic β-galactosidase acts on agarooligosaccharides for complete hydrolysis of agarose into monomers. Appl. Environ. Microbiol. 2014, 80, 5965–5973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merkel, A.; Voropaeva, D.; Ondrušek, M. The impact of integrated nanofiltration and electrodialytic processes on the chemical composition of sweet and acid whey streams. J. Food Eng. 2021, 298, 110500. [Google Scholar] [CrossRef]
- Bologa, M.K.; Vrabie, E.G.; Stepurina, T.G. Features of mineralization of protein concentrates during the electrophysical treatment of whey. Surf. Eng. Appl. 2013, 49, 504–508. [Google Scholar] [CrossRef]
- Wronkowska, M.; Jadacka, M.; Soral-Śmietana, M.; Zander, L.; Dajnowiec, F.; Banaszczyk, P.; Jeliński, T.; Szmatowicz, B. ACID whey concentrated by ultrafiltration a tool for modeling bread properties. LWT 2015, 61, 172–176. [Google Scholar] [CrossRef]
- Šimun, Z.; Neven, A.; Jasmina, H.D.S.; Zamberlin, Š.; Antunac, N.; Havranek, J.; Samaržija, D. Mineral elements in milk and dairy products. Mljekarstvo 2012, 62, 111–125. [Google Scholar]
- de Albuquerque, T.L.; de Sousa, M.; Gomes, E.; Silva, N.C.; Girão Neto, C.; Gonçalves, L.; Fernandez-Lafuente, R.; Rocha, M. β-Galactosidase from Kluyveromyces lactis: Characterization, production, immobilization and applications—A review. Int. J. Biol. 2021, 191, 881–898. [Google Scholar] [CrossRef]
- Jurado, E.; Camacho, F.; Luzón, G.; Vicaria, J.M. Kinetic models of activity for β-galactosidases: Influence of pH, ionic concentration and temperature. Enzyme Microb. Technol. 2004, 34, 33–40. [Google Scholar] [CrossRef]
- Chandrapala, J.; Duke, M.C.; Gray, S.R.; Zisu, B.; Weeks, M.; Palmer, M.; Vasiljevic, T. Properties of acid whey as a function of pH and temperature. J. Dairy Sci. 2015, 98, 4352–4363. [Google Scholar] [CrossRef] [PubMed]
- Frankowski, K.M.; Miracle, R.E.; Drake, M.A. The role of sodium in the salty taste of permeate. J. Dairy Sci. 2014, 97, 5356–5370. [Google Scholar] [CrossRef]
- Bozanic, R.; Barukcic, I.; Lisak, K.; Jakopovic; Tratnik, L. Possibilities of whey utilisation. Austin J. Nutr. Food Sci. 2014, 2, 1036–1042. [Google Scholar]
- Czyzewska, K.; Trusek, A. Encapsulated NOLATM Fit 5500 Lactase—An economically beneficial way to obtain lactose-free milk at low temperature. Catalysts 2021, 11, 527. [Google Scholar] [CrossRef]
- Mahoney, R.R. Lactose: Enzymatic Modification. In Advanced Dairy Chemistry; Springer: New York, NY, USA, 1997; pp. 77–125. [Google Scholar] [CrossRef]
- Warmerdam, A.; Boom, R.M.; Janssen, A.E. Β-Galactosidase Stability at High Substrate Concentrations. SpringerPlus 2013, 2, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samadov, R.; Ciprovica, I.; Zolnere, K.; Cinkmanis, I. The optimization of acid whey permeate hydrolysis for glucose-galactose syrup production. In Proceedings of the 13th Baltic Conference on Food Science and Technology, FOODBALT 2019 Joined with 5th North and East European Congress on Food, NEEFood 2019 “Food. Nutrition. Well-Being”, Jelgava, Latvia, 2–3 May 2019; pp. 254–257. [Google Scholar] [CrossRef]
- Mariyani, N.; Faridah, D.N.; Khusniati, T.; Lioe, H.N. Hydrolysis of UHT milk lactose by partially purified crude enzyme of β-galactosidase obtained from Lactobacillus plantarum B123 indigenous strain Hydrolysis of UHT milk lactose by partially purified crude enzyme of β-galactosidase obtained from Lactobac. Int. Food Res. J. 2015, 22, 2274–2279. [Google Scholar]
- Luzzi, G.; Steffens, M.; Clawin-Rädecker, I.; Hoffmann, W.; Franz, C.M.A.P.; Fritsche, J.; Lorenzen, P.C. Enhancing the sweetening power of lactose by enzymatic modification in the reformulation of dairy products. Int. J. Dairy Technol. 2020, 73, 502–512. [Google Scholar] [CrossRef] [Green Version]
- Suárez, S.; Guerrero, C.; Vera, C.; Illanes, A. Effect of particle size and enzyme load on the simultaneous reactions of lactose hydrolysis and transgalactosylation with glyoxyl-agarose immobilized β-galactosidase from Aspergillus oryzae. Process Biochem. 2018, 73, 56–64. [Google Scholar] [CrossRef]
- Botvynko, A.; Bednářová, A.; Henke, S.; Shakhno, N.; Čurda, L. Production of galactooligosaccharides using various combinations of the commercial β-galactosidases. Biochem. Biophys. 2019, 517, 762–766. [Google Scholar] [CrossRef]
- Vénica, C.I.; Bergamini, C.V.; Rebechi, S.R.; Perotti, M.C. Galacto-oligosaccharides formation during manufacture of different varieties of yogurt. Stability through storage. LWT 2015, 63, 198–205. [Google Scholar] [CrossRef] [Green Version]
- Gänzle, M.G. Enzymatic synthesis of galacto-oligosaccharides and other lactose derivatives (hetero-oligosaccharides) from lactose. Int. Dairy J. 2012, 22, 116–122. [Google Scholar] [CrossRef]
- Osman, A. Synthesis of Prebiotic Galacto-Oligosaccharides: Science and Technology. In Probiotics, Prebiotics, and Synbiotics: Bioactive Foods in Health Promotion, 1st ed.; Ross, R., Preedy, V., Eds.; Elsevier: London, UK, 2016; pp. 135–154. [Google Scholar] [CrossRef]
- de Vrese, M.; Schrezenmeir, J. Probiotics, Prebiotics, and Synbiotics. In Food Biotechnology. Advances in Biochemical Engineering/Biotechnology; Stahl, U., Donalies, U.E., Nevoigt, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 1–66. [Google Scholar] [CrossRef]
- Fischer, C.; Kleinschmidt, T. Synthesis of galactooligosaccharides using sweet and acid whey as a substrate. Int. Dairy J. 2015, 48, 15–22. [Google Scholar] [CrossRef]
- Cheng, S.; Hummel, M.; Dahal, B.; Gu, Z.; Kharel, P.; Martínez-Monteagudo, S.I. A two-step process for the synthesis of sweetening syrup from aqueous lactose. LWT 2020, 117, 108659. [Google Scholar] [CrossRef]
- Jia, D.-X.; Zhou, L.; Zheng, Y.-G. Properties of a novel thermostable glucose isomerase mined from Thermus oshimai and its application to preparation of high fructose corn syrup. Enzyme Microb. Technol. 2017, 99, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yang, S.; Saravanamurugan, S.; Riisager, A. Glucose isomerization by enzymes and chemo-catalysts: Status and current advances. ACS Catal. 2017, 7, 3010–3029. [Google Scholar] [CrossRef]
- Foda, F.F.; Abd El-Rahman, A.A. Effect of some polluted metals on the activity and kinetics of immobilized glucose isomerase (Sweetzyme type-T) from Streptomyces murinus. Ann. Agric. Sci. 2000, 38, 2217–2228. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Permeates | Fat (%) | Proteins (%) | Lactose (%) | Total Solids (%) | pH |
|---|---|---|---|---|---|
| Sweet whey, 5% | <0.1 | 0.2 ± 0.1 | 3.8 ± 0.1 | 4.6 ± 0.1 | 6.1 ± 0.1 |
| Acid whey, 5% | <0.1 | 0.5 ± 0.1 | 4.2 ± 0.2 | 5.2 ± 0.2 | 4.6 ± 0.1 |
| Sweet whey, 20% | <0.1 | 0.7 ± 0.1 | 18.8 ± 0.1 | 20.7 ± 0.2 | 6.2 ± 0.1 |
| Acid whey, 20% | <0.1 | 1.2 ± 0.2 | 18.2 ± 0.2 | 20.3 ± 0.3 | 4.5 ± 0.1 |
| Permeates | Ca2+ | Na+ | K+ | Mg2+ | PO43− |
|---|---|---|---|---|---|
| Sweet whey, 5% | 349 ± 14 | 429 ± 11 | 1480 ± 100 | 57 ± 9 | 275 ± 61 |
| Acid whey, 5% | 1322 ± 83 | 469 ± 67 | 1636 ± 189 | 123 ± 16 | 736 ± 34 |
| Sweet whey, 20% | 1071 ± 99 | 660 ± 57 | 2710 ± 55 | 260 ± 13 | 1265 ± 50 |
| Acid whey, 20% | 3400 ± 86 | 1100 ± 55 | 5600 ± 140 | 340 ± 18 | 2200 ± 56 |
| Time | |||||||
|---|---|---|---|---|---|---|---|
| Enzyme | Sugar | 1 min | 30 min | 1 h | 2 h | 3 h | 4 h |
| Sweet whey permeate | |||||||
| N5500 | Glucose | 2.6 ± 0.2 c | 8.1 ± 0.4 e | 29.9 ± 0.1 a | 33.0 ± 1.0 c | 41.9 ± 0.2 b | 43.9 ± 1.4 b |
| Galactose | 1.6 ± 0.1 b | 3.8 ± 0.2 e | 12.4 ± 0.5 c | 14.9 ± 0.7 b | 15.5 ± 1.0 c | 16.4 ± 0.9 d | |
| Lactose | 88.6 ± 1.1 bc | 66.6 ± 0.1 a | 51.5 ± 0.6 c | 39.8 ± 1.2 b | 26.3 ± 1.1 b | 22.1 ± 0.5 a | |
| H5200 | Glucose | 3.1 ± 0.1 b | 21.9 ± 0.5 a | 30.0 ± 0.4 a | 43.9 ± 1.5 a | 46.5 ± 2.9 a | 51.0 ± 3.4 a |
| Galactose | 1.6 ± 0.1 b | 8.3 ± 0.6 c | 11.7 ± 0.4 c | 16.8 ± 0.3 a | 17.0 ± 2.5 b | 20.9 ± 1.1 b | |
| Lactose | 87.1 ± 0.7 c | 50.8 ± 0.9 d | 39.5 ± 0.1 e | 22.7 ± 0.9 d | 11.4 ± 1.2 d | 2.1 ± 0.6 e | |
| G2 | Glucose | 1.9 ± 0.1 d | 13.9 ± 1.0 c | 25.0 ± 0.6 b | 29.7 ± 1.6 d | 35.9 ± 0.6 d | 36.0 ± 0.3 c |
| Galactose | 0.9 ± 0.1 c | 6.5 ± 0.1 d | 11.2 ± 1.2 cd | 12.8 ± 1.7 c | 14.9 ± 1.5 c | 15.2 ± 0.1 e | |
| Lactose | 86.5 ± 1.1 c | 57.2 ± 1.2 c | 49.4 ± 0.7 d | 33.2 ± 0.9 c | 21.4 ± 0.1 c | 13.9 ± 0.6 c | |
| Acid whey permeate | |||||||
| N5500 | Glucose | 7.1 ± 0.5 a | 10.8 ± 1.3 d | 15.9 ± 1.9 c | 26.7 ± 2.6 d | 40.8 ± 2.4 b | 43.5 ± 1.6 b |
| Galactose | 7.7 ± 1.6 a | 10.5 ± 0.6 b | 14.0 ± 0.5 b | 15.2 ± 1.2 b | 21.0 ± 0.5 a | 25.3 ± 1.7 a | |
| Lactose | 90.5 ± 0.8 ab | 64.0 ± 1.0 b | 54.9 ± 0.3 a | 42.3 ± 1.3 ab | 32.7 ± 0.8 a | 19.8 ± 0.5 b | |
| H5200 | Glucose | 5.7 ± 1.2 a | 14.7 ± 0.7 c | 23.6 ± 0.5 a | 38.0 ± 1.8 b | 47.8 ± 1.3 a | 50.6 ± 2.1 a |
| Galactose | 6.2 ± 1.8 a | 11.0 ± 0.6 b | 15.4 ± 2.2 c | 16.8 ± 1.3 ab | 17.6 ± 0.6 b | 22.4 ± 1.4 ab | |
| Lactose | 92.2 ± 1.1 a | 56.1 ± 1.3 c | 38.1 ± 0.8 e | 22.8 ± 0.9 d | 11.3 ± 1.0 d | 5.9 ± 0.3 d | |
| G2 | Glucose | 6.0 ± 1.0 a | 16.0 ± 0.3 b | 16.3 ± 1.1 c | 22.5 ± 1.7 e | 26.9 ± 1.3 c | 32.5 ± 1.7 d |
| Galactose | 6.8 ± 1.8 a | 13.0 ± 0.2 a | 10.3 ± 0.1 d | 12.7 ± 0.4 c | 15.4 ± 1.6 b | 17.2 ± 1.6 c | |
| Lactose | 89.8 ± 1.1 b | 58.6 ± 1.3 c | 53.7 ± 0.3 b | 42.4 ± 0.8 a | 31.5 ± 1.3 a | 20.2 ± 1.0 b | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Majore, K.; Ciprovica, I. Bioconversion of Lactose into Glucose–Galactose Syrup by Two-Stage Enzymatic Hydrolysis. Foods 2022, 11, 400. https://doi.org/10.3390/foods11030400
Majore K, Ciprovica I. Bioconversion of Lactose into Glucose–Galactose Syrup by Two-Stage Enzymatic Hydrolysis. Foods. 2022; 11(3):400. https://doi.org/10.3390/foods11030400
Chicago/Turabian StyleMajore, Kristine, and Inga Ciprovica. 2022. "Bioconversion of Lactose into Glucose–Galactose Syrup by Two-Stage Enzymatic Hydrolysis" Foods 11, no. 3: 400. https://doi.org/10.3390/foods11030400
APA StyleMajore, K., & Ciprovica, I. (2022). Bioconversion of Lactose into Glucose–Galactose Syrup by Two-Stage Enzymatic Hydrolysis. Foods, 11(3), 400. https://doi.org/10.3390/foods11030400