
Antibacterial Effect of Oregano Essential Oil against Vibrio vulnificus and Its Mechanism

Abstract
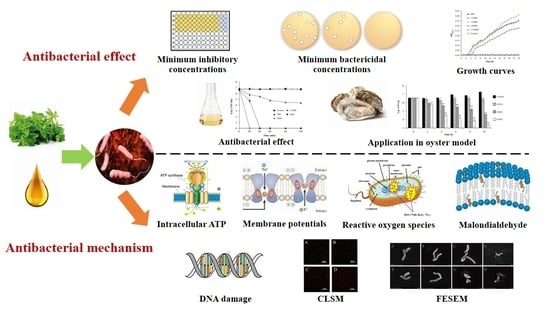
:
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Bacterial Strains and Culture Conditions
2.3. Determination of MICs and MBCs
2.4. Growth Curves
2.5. Antibacterial Effect of OEO on V. vulnificus in APW
2.6. Intracellular ATP Concentrations
2.7. Membrane Potentials
2.8. Detection of Reactive Oxygen Species (ROS)
2.9. Malondialdehyde (MDA) Content Assay
2.10. DNA Comet Assay
2.11. Confocal Laser Scanning Microscopy (CLSM) Observations
2.12. Field-Emission Scanning Electron Microscopy (FESEM) Observations
2.13. Antibacterial Effect of OEO against V. vulnificus in Fresh Artificially Contaminated Oysters Model
2.14. Statistical Analysis
3. Results
3.1. MIC and MBC of OEO against V. vulnificus
3.2. Effect of OEO on V. vulnificus Growth
3.3. Antibacterial Activity of OEO toward V. vulnificus in APW
3.4. Effect of OEO on Intracellular ATP Concentration
3.5. Effect of OEO on Membrane Potential
3.6. Effect of OEO on Intracellular ROS Generation
3.7. Effect of OEO on Intracellular MDA Content of V. vulnificus
3.8. Effect of OEO on DNA Damage
3.9. CLSM-Based Observations of Cell Membrane Injury
3.10. FESEM-Based Observations of Cell Morphology
3.11. Antibacterial Effect of OEO on V. vulnificus in Fresh Oysters
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviation
| OEO | Oregano essential oil |
| V. vulnificus | Vibrio vulnificus |
| MIC | Minimum inhibitory concentration |
| MBC | Minimum bactericidal concentration |
| ROS | Reactive oxygen species |
| MDA | Malondialdehyde |
| DMSO | Dimethyl sulfoxide |
| APW | Alkaline peptone water |
| TSA | Tryptone soya agar |
| PBS | Phosphate-buffered saline |
| PBS | Phosphate-buffered saline |
| PI | Propidium iodide |
| CLSM | Confocal laser scanning microscopy |
| FESEM | Field-emission scanning electron microscopy |
| OD | Optical density |
| SD | Standard deviation |
References
- Jones, M.K.; Oliver, J.D. Vibrio vulnificus: Disease and Pathogenesis. Infect. Immun. 2009, 77, 1723–1733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Chung, H.Y.; Kwon, J.G.; Choi, S.H.; Lee, J.H. Fresh crab plays an important role as a nutrient reservoir for the rapid propagation of Vibrio vulnificus. Front Microbiol. 2021, 12, 645860. [Google Scholar] [CrossRef] [PubMed]
- Hubert, C.L.; Michell, S.L. A universal oyster infection model demonstrates that Vibrio vulnificus Type 6 secretion systems have antibacterial activity in vivo. Environ. Microbiol. 2020, 22, 4318–4393. [Google Scholar] [CrossRef] [PubMed]
- Baker-Austin, C.; Oliver, J.D. Vibrio vulnificus: New insights into a deadly opportunistic pathogen. Trends Microbiol. 2020, 28, 81–82. [Google Scholar] [CrossRef]
- Phillips, K.E.; Satchell, K.J.F. Vibrio vulnificus: From Oyster Colonist to Human Pathogen. PLoS Pathog. 2017, 13, e1006053. [Google Scholar] [CrossRef] [Green Version]
- Carla, H.-C.; Chung-Te, L.; Verónica, T.-E.; Eva, S.; David, P.; Felipe, R.-L.; Lluis, T.; Carmen, A. Adaptation to host in Vibrio vulnificus, a zoonotic pathogen that causes septicemia in fish and humans. Environ. Microbiol. 2019, 21, 3118–3139. [Google Scholar] [CrossRef]
- Lopez-Perez, M.; Jayakumar, J.M.; Haro-Moreno, J.M.; Zaragoza-Solas, A.; Reddi, G.; Rodriguez-Valera, F.; Shapiro, O.H.; Alam, M.; Almagro-Moreno, S. Evolutionary model of cluster divergence of the emergent marine pathogen Vibrio vulnificus: From genotype to ecotype. Mbio 2019, 10, e02852-18. [Google Scholar] [CrossRef] [Green Version]
- Saxena, S.; Gautam, S.; Sharma, A. Microbial Decontamination of Honey of Indian Origin Using Gamma Radiation and Its Biochemical and Organoleptic Properties. J. Food Sci. 2010, 75, M19–M27. [Google Scholar] [CrossRef]
- Bouletis, A.D.; Arvanitoyannis, I.S.; Hadjichristodoulou, C. Application of modified atmosphere packaging on aquacultured fish and fish products: A review. Crit. Rev. Food Sci. Nutr. 2017, 57, 2263–2285. [Google Scholar] [CrossRef]
- Inanli, A.G.; Tumerkan, E.T.A.; El Abed, N.; Regenstein, J.M.; Ozogul, F. The impact of chitosan on seafood quality and human health: A review. Trends Food Sci. Technol. 2020, 97, 404–416. [Google Scholar] [CrossRef]
- Chao, G.; Jiao, X.; Zhou, X.; Yang, Z.; Pan, Z.; Huang, J.; Zhou, L.; Qian, X. Systematic functional pandemic strain-specific genes, three genomic islands, two T3SSs in foodborne, and clinical Vibrio parahaemolyticus isolates in China. Foodborne Pathog. Dis. 2009, 6, 689–698. [Google Scholar] [CrossRef] [PubMed]
- Erikson, U.; Standal, I.B.; Aursand, I.G.; Veliyulin, E.; Aursand, M. Use of NMR in fish processing optimization: A review of recent progress. Magn. Reson. Chem. 2012, 50, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Habibullah-Al-Mamun, M.; Ahmed, M.K.; Islam, M.S.; Tokumura, M.; Masunaga, S. Distribution of polycyclic aromatic hydrocarbons (PAHs) in commonly consumed seafood from coastal areas of bangladesh and associated human health implications. Environ. Geochem. Health 2019, 41, 1105–1121. [Google Scholar] [CrossRef] [PubMed]
- Falleh, H.; Ben Jemaa, M.; Saada, M.; Ksouri, R. Essential oils: A promising eco-friendly food preservative. Food Chem. 2020, 330, 127268. [Google Scholar] [CrossRef]
- Wei, H.K.; Chen, G.; Wang, R.J.; Peng, J. Oregano essential oil decreased susceptibility to oxidative stress-induced dysfunction of intestinal epithelial barrier in rats. J. Funct. Foods 2015, 18, 1191–1199. [Google Scholar] [CrossRef]
- Leyva-Lopez, N.; Gutierrez-Grijalva, E.P.; Vazquez-Olivo, G.; Heredia, J.B. Essential oils of oregano: Biological activity beyond their antimicrobial properties. Molecules 2017, 22, 989. [Google Scholar] [CrossRef] [Green Version]
- Manso, S.; Becerril, R.; Nerin, C.; Gomez-Lus, R. Influence of pH and temperature variations on vapor phase action of an antifungal food packaging against five mold strains. Food Control 2015, 47, 20–26. [Google Scholar] [CrossRef]
- Muriel-Galet, V.; Cran, M.J.; Bigger, S.W.; Hernandez-Munoz, P.; Gavara, R. Antioxidant and antimicrobial properties of ethylene vinyl alcohol copolymer films based on the release of oregano essential oil and green tea extract components. J. Food Eng. 2015, 149, 9–16. [Google Scholar] [CrossRef]
- Elshafie, H.S.; Armentano, M.F.; Carmosino, M.; Bufo, S.A.; De Feo, V.; Camele, I. Cytotoxic activity of Origanum Vulgare L. on hepatocellular carcinoma cell line HepG2 and evaluation of its biological activity. Molecules 2017, 22, 1435. [Google Scholar] [CrossRef] [Green Version]
- Llana-Ruiz-Cabello, M.; Maisanaba, S.; Puerto, M.; Pichardo, S.; Jos, A.; Moyano, R.; Camean, A.M. A subchronic 90-day oral toxicity study of Origanum vulgare essential oil in rats. Food Chem. Toxic. 2017, 101, 36–47. [Google Scholar] [CrossRef]
- Approved Standard M7-A10; Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically. 8th ed. LSI: Wayne, PA, USA, 2018.
- Shi, C.; Sun, Y.; Liu, Z.Y.; Guo, D.; Sun, H.H.; Sun, Z.; Chen, S.; Zhang, W.T.; Wen, Q.W.; Peng, X.L.; et al. Inhibition of Cronobacter sakazakii virulence factors by citral. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashrafudoulla, M.; Mizan, M.F.R.; Ha, A.J.W.; Park, S.H.; Ha, S.D. Antibacterial and antibiofilm mechanism of eugenol against antibiotic resistance Vibrio parahaemolyticus. Food Microbiol. 2020, 91, 103500. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, E.; Garcia, S.; Heredia, N. Extracts of edible and medicinal plants damage membranes of Vibrio cholerae. Appl. Environ. Microb. 2010, 76, 6888–6894. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.K.; Ma, X.Y.; Li, Y.; Xu, H.D.; Han, X.Y.; Wang, R.X.; Zhao, P.Y.; Li, Z.Y.; Shi, C. Antimicrobial activity and antibiofilm potential of coenzyme Q0 against Salmonella typhimurium. Foods 2021, 10, 1211. [Google Scholar] [CrossRef] [PubMed]
- Shivaprasad, D.P.; Taneja, N.K.; Lakra, A.; Sachdev, D. In vitro and in situ abrogation of biofilm formation in E. coli by vitamin C through ROS generation, disruption of quorum sensing and exopolysaccharide production. Food Chem. 2021, 341, 128171. [Google Scholar] [CrossRef]
- Cao, J.J.; Liu, H.; Wang, Y.; He, X.X.; Jiang, H.Y.; Yao, J.J.; Xia, F.; Zhao, Y.N.; Chen, X.F. Antimicrobial and antivirulence efficacies of citral against foodborne pathogen Vibrio parahaemolyticus RIMD 2210633. Food Control 2021, 120, 107507. [Google Scholar] [CrossRef]
- Kim, M.J.; Miks-Krajnik, M.; Kumar, A.; Yuk, H.G. Inactivation by 405 +/- 5 nm light emitting diode on Escherichia coli O157:H7, Salmonella Typhimurium, and Shigella sonnei under refrigerated condition might be due to the loss of membrane integrity. Food Control 2016, 59, 99–107. [Google Scholar] [CrossRef]
- Qi, J.Y.; Gong, M.; Zhang, R.; Song, Y.M.; Liu, Q.; Zhou, H.Y.; Wang, J.; Mei, Y.F. Evaluation of the antibacterial effect of tea tree oil on Enterococcus faecalis and biofilm in vitro. J. Ethnopharmacol. 2021, 281, 114566. [Google Scholar] [CrossRef]
- Tian, L.; Wang, X.Y.; Liu, R.J.; Zhang, D.; Wang, X.; Sun, R.C.; Guo, W.Y.; Yang, S.Q.; Li, H.; Gong, G.L. Antibacterial mechanism of thymol against Enterobacter sakazakii. Food Control 2021, 123, 107716. [Google Scholar] [CrossRef]
- Navarro-Navarro, M.; Ruiz-Bustos, P.; Valencia, D.; Robles-Zepeda, R.; Ruiz-Bustos, E.; Virues, C.; Hernandez, J.; Dominguez, Z.; Velazquez, C. Antibacterial activity of sonoran propolis and some of Its constituents against clinically significant Vibrio species. Foodborne Pathog. Dis. 2013, 10, 150–158. [Google Scholar] [CrossRef]
- Ozogul, Y.; Boga, E.K.; Akyol, I.; Durmus, M.; Ucar, Y.; Regenstein, J.M.; Kosker, A.R. Antimicrobial activity of thyme essential oil nanoemulsions on spoilage bacteria of fish and food-borne pathogens. Food Biosci. 2020, 36, 100635. [Google Scholar] [CrossRef]
- Ozogul, Y.; Ozogul, F.; Kulawik, P. The antimicrobial effect of grapefruit peel essential oil and its nanoemulsion on fish spoilage bacteria and food-borne pathogens. LWT-Food Sci. Technol. 2021, 136, 110362. [Google Scholar] [CrossRef]
- Dominguez-Borbor, C.; Sanchez-Rodriguez, A.; Sonnenholzner, S.; Rodriguez, J. Essential oils mediated antivirulence therapy against vibriosis in Penaeus vannamei. Aquaculture 2020, 529, 735639. [Google Scholar] [CrossRef]
- Kim, N.H.; Kim, H.W.; Moon, H.; Rhee, M.S. Sodium chloride significantly enhances the bactericidal actions of carvacrol and thymol against the halotolerant species Escherichia coli O157:H7, Listeria monocytogenes, and Staphylococcus aureus. LWT-Food Sci. Technol. 2020, 122, 109015. [Google Scholar] [CrossRef]
- Spari, D.; Beldi, G. Extracellular ATP as an Inter-Kingdom signaling molecule: Release mechanisms by bacteria and its implication on the host. Int. J. Mol. Sci. 2020, 21, 5590. [Google Scholar] [CrossRef]
- Guo, D.; Wang, S.; Li, J.H.; Bai, F.T.; Yang, Y.P.; Xu, Y.F.; Liang, S.; Xia, X.D.; Wang, X.; Shi, C. The antimicrobial activity of coenzyme Q0 against planktonic and biofilm forms of Cronobacter sakazakii. Food Microbiol. 2019, 86, 103337. [Google Scholar] [CrossRef]
- Shi, C.; Sun, Y.; Zheng, Z.W.; Zhang, X.R.; Song, K.K.; Jia, Z.Y.; Chen, Y.F.; Yang, M.C.; Liu, X.; Dong, R.; et al. Antimicrobial activity of syringic acid against Cronobacter sakazakii and its effect on cell membrane. Food Chem. 2015, 197, 100–106. [Google Scholar] [CrossRef]
- Li, X.Z.; Nikaido, H. Efflux-mediated drug resistance in bacteria: An update. Drugs 2009, 64, 159–204. [Google Scholar] [CrossRef]
- Costa, T.R.D.; Felisberto-Rodrigues, C.; Meir, A.; Prevost, M.S.; Redzej, A.; Trokter, M.; Waksman, G. Secretion systems in Gram-negative bacteria: Structural and mechanistic insights. Nat. Rev. Microbiol. 2015, 13, 343–359. [Google Scholar] [CrossRef]
- Scariot, F.J.; Foresti, L.; Delamare, A.P.L.; Echeverrigaray, A.P.L.S. Activity of monoterpenoids on the in vitro growth of two Colletotrichum species and the mode of action on C. acutatum. Pestic. Biochem. Phys. 2020, 170, 104698. [Google Scholar] [CrossRef]
- Bot, C.; Prodan, C. Probing the membrane potential of living cells by dielectric spectroscopy. Eur. Biophys. J. 2009, 38, 1049–1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, C.; Song, K.K.; Zhang, X.R.; Sun, Y.; Sui, Y.; Chen, Y.F.; Jia, Z.Y.; Sun, H.H.; Sun, Z.; Xia, X.D. Antimicrobial activity and possible mechanism of action of citral against Cronobacter sakazakii. PLoS ONE 2016, 11, e0159006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vatansever, F.; de Melo, W.C.M.A.; Avci, P.; Vecchio, D.; Sadasivam, M.; Gupta, A.; Chandran, R.; Karimi, M.; Parizotto, N.A.; Yin, R.; et al. Antimicrobial strategies centered around reactive oxygen species--bactericidal antibiotics, photodynamic therapy, and beyond. FEMS Microbiol. Rev. 2013, 37, 955–989. [Google Scholar] [CrossRef] [Green Version]
- Bajpai, V.K.; Park, I.; Khan, I.; Alshammari, F.H.; Kumar, P.; Chen, L.; Na, M.; Huh, Y.S.; Han, Y.K.; Shukla, S. (−)-Tetrahydroberberrubine∙acetate accelerates antioxidant potential and inhibits food associated Bacillus cereus in rice. Food Chem. 2020, 339, 127902. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Lu, J.Y.; Duan, H.B.; Yang, J.; Tang, C.B. Biofilm inhibition and mode of action of Epigallocatechin Gallate against Vibrio mimicus. Food Control 2020, 113, 107148. [Google Scholar] [CrossRef]
- Yang, S.K.; Yusoff, K.; Ajat, M.; Yap, W.S.; Lim, S.H.E.; Lai, K.S. Antimicrobial activity and mode of action of terpene linalyl anthranilate against carbapenemase-producing Klebsiella pneumoniae. J. Pharm. Anal. 2020, 11, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Su, L.J.; Zhang, J.H.; Gomez, H.; Murugan, R.; Hong, X.; Xu, D.X.; Jiang, F.; Peng, Z.Y. Reactive oxygen species-induced lipid peroxidation in apoptosis, autophagy, and ferroptosis. Oxid. Med. Cell. Longev. 2019, 2019, 5080843. [Google Scholar] [CrossRef] [Green Version]
- Ajiboye, T.O.; Aliyu, M.; Isiaka, I.; Haliru, F.Z.; Ibitoye, O.B.; Uwazie, J.N.; Muritala, H.F.; Bello, S.A.; Yusuf, I.I.; Mohammed, A.O. Contribution of reactive oxygen species to (+)-catechin-mediated bacterial lethality. Chem. Biol. Interact. 2016, 258, 276–287. [Google Scholar] [CrossRef]
- Prasad, K.S.; Patel, H.; Patel, T.; Patel, K.; Selvaraj, K. Biosynthesis of Se nanoparticles and its effect on UV-induced DNA damage. Colloids Surf. B Biointerfaces 2013, 103, 261–266. [Google Scholar] [CrossRef]
- Liu, G.R.; Ren, G.M.; Zhao, L.; Cheng, L.; Wang, C.T.; Sun, B.G. Antibacterial activity and mechanism of bifidocin A against Listeria monocytogenes. Food Control 2017, 73, 854–861. [Google Scholar] [CrossRef]
- Cui, H.Y.; Zhang, C.H.; Li, C.Z.; Lin, L. Antibacterial mechanism of oregano essential oil. Ind. Crop. Prod. 2019, 139, 111498. [Google Scholar] [CrossRef]
- Dutra, T.V.; Castro, J.C.; Menezes, J.L.; Ramos, T.R.; do Prado, I.N.; Machinski, M.; Mikcha, J.M.G.; de Abreu, B.A. Bioactivity of oregano (Origanum vulgare) essential oil against Alicyclobacillus spp. Ind. Crop. Prod. 2018, 129, 345–349. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Oliver, J.D. Vibrio vulnificus: New insights into a deadly opportunistic pathogen. Environ. Microbiol. 2017, 20, 423–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahmoud, B.S.M. Controlling Vibrio vulnificus and spoilage bacteria in fresh shucked oysters using natural antimicrobials. Lett. Appl. Microbiol. 2013, 58, 1–7. [Google Scholar] [CrossRef]
- Hernandez, H.; Frankova, A.; Sykora, T.; Kloucek, P.; Kourimska, L.; Kucerova, I.; Banout, J. The effect of oregano essential oil on microbial load and sensory attributes of dried meat. J. Sci. Food Agric. 2016, 97, 82–87. [Google Scholar] [CrossRef]
- Zhou, Y.; Sun, S.S.; Bei, W.Y.; Zahi, M.R.; Yuan, Q.P.; Liang, H. Preparation and antimicrobial activity of oregano essential oil Pickering emulsion stabilized by cellulose nanocrystals. Int. J. Biol. Macromol. 2018, 112, 7–13. [Google Scholar] [CrossRef]
- Costa, R.C.; Daitx, T.S.; Mauler, R.S.; Silva, N.M.; Miotto, M.; Crespo, J.S.; Carli, L.N. Poly(hydroxybutyrate-co-hydroxyvalerate)-based nanocomposites for antimicrobial active food packaging containing oregano essential oil. Food Packag. Shelf Life 2020, 26, 100602. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | MIC (μL/mL) | MBC (μL/mL) |
|---|---|---|
| ATCC 27562 | 0.06 | 0.06 |
| VV02 | 0.08 | 0.08 |
| VV15 | 0.15 | 0.15 |
| VV20 | 0.08 | 0.08 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, K.; Zhao, P.; He, Y.; Kang, S.; Shen, C.; Wang, S.; Guo, M.; Wang, L.; Shi, C. Antibacterial Effect of Oregano Essential Oil against Vibrio vulnificus and Its Mechanism. Foods 2022, 11, 403. https://doi.org/10.3390/foods11030403
Luo K, Zhao P, He Y, Kang S, Shen C, Wang S, Guo M, Wang L, Shi C. Antibacterial Effect of Oregano Essential Oil against Vibrio vulnificus and Its Mechanism. Foods. 2022; 11(3):403. https://doi.org/10.3390/foods11030403
Chicago/Turabian StyleLuo, Kunyao, Pengyu Zhao, Yifei He, Shengnan Kang, Chenyu Shen, Shuo Wang, Meixian Guo, Lehui Wang, and Chao Shi. 2022. "Antibacterial Effect of Oregano Essential Oil against Vibrio vulnificus and Its Mechanism" Foods 11, no. 3: 403. https://doi.org/10.3390/foods11030403
APA StyleLuo, K., Zhao, P., He, Y., Kang, S., Shen, C., Wang, S., Guo, M., Wang, L., & Shi, C. (2022). Antibacterial Effect of Oregano Essential Oil against Vibrio vulnificus and Its Mechanism. Foods, 11(3), 403. https://doi.org/10.3390/foods11030403