
Sensory Properties and Main Differential Metabolites Influencing the Taste Quality of Dry-Cured Beef during Processing

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Processing of Dry-Cured Beef
2.3. Physicochemical Properties and Protein Hydrolysis and Oxidation
2.4. Sensory Assessment
2.5. Analysis of Dry-Cured Beef Metabolomics Using UPLC-MS/MS
2.5.1. Sample Preparation
2.5.2. High-Performance Liquid Chromatography-Mass Spectrometry and Qualitative and Quantitative Analysis of Metabolites
2.5.3. Analysis of Metabolite Differences and Metabolic Pathways
2.6. Statistical Analysis
3. Results and Discussion
3.1. Physicochemical Properties and Protein Hydrolysis and Oxidation Analyses
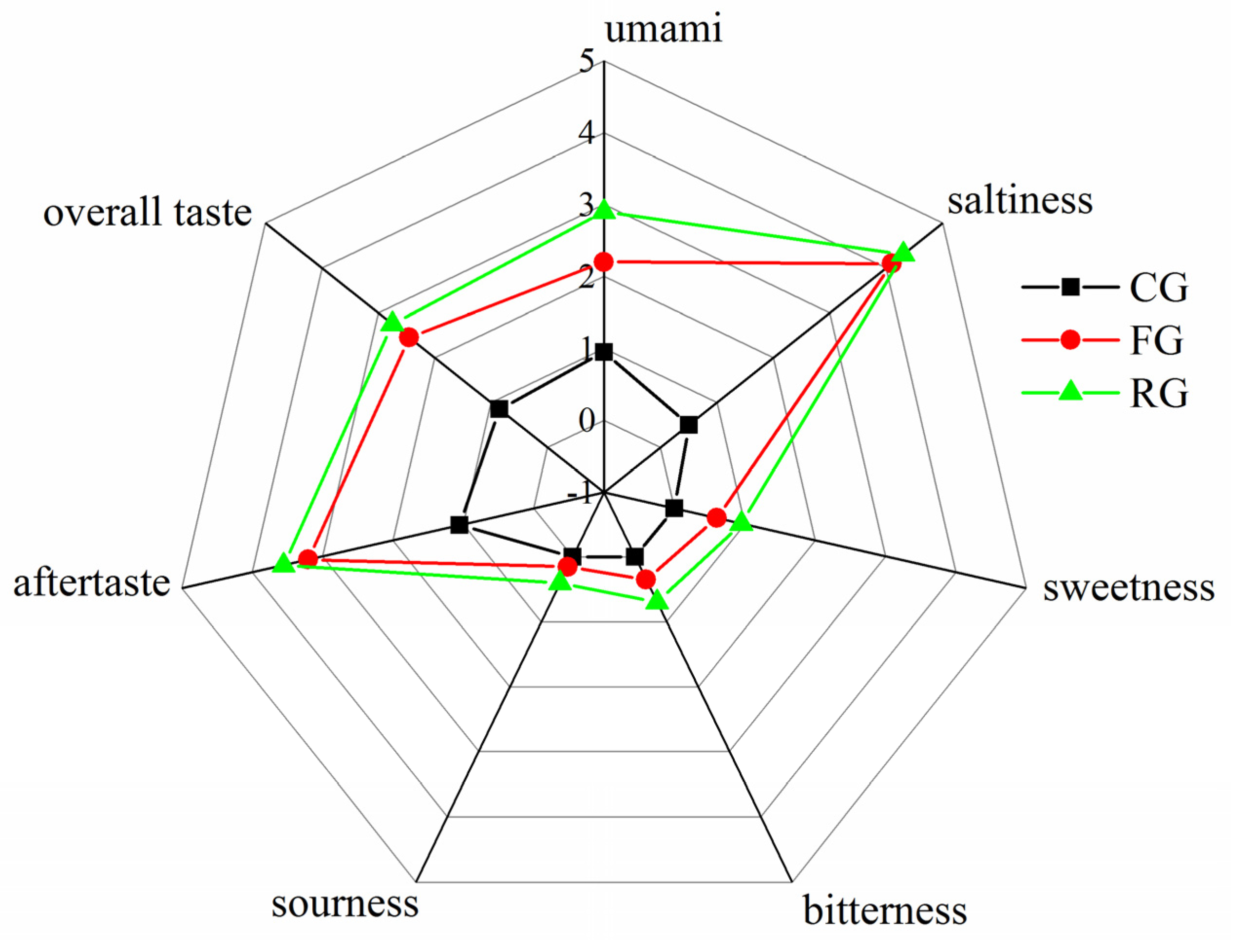
3.2. Sensory Assessment
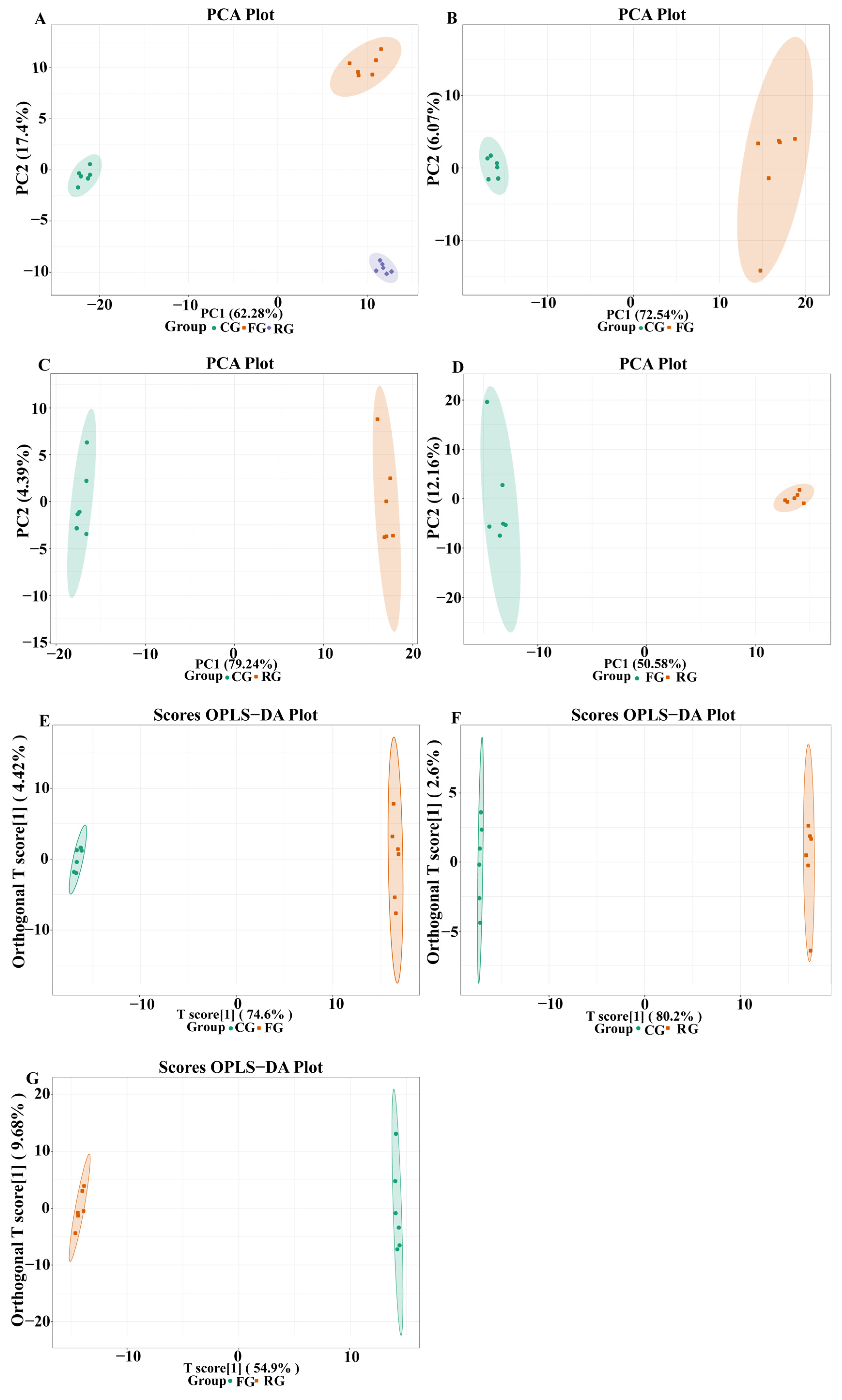
3.3. Metabolite Profiles of Dry-Cured Beef
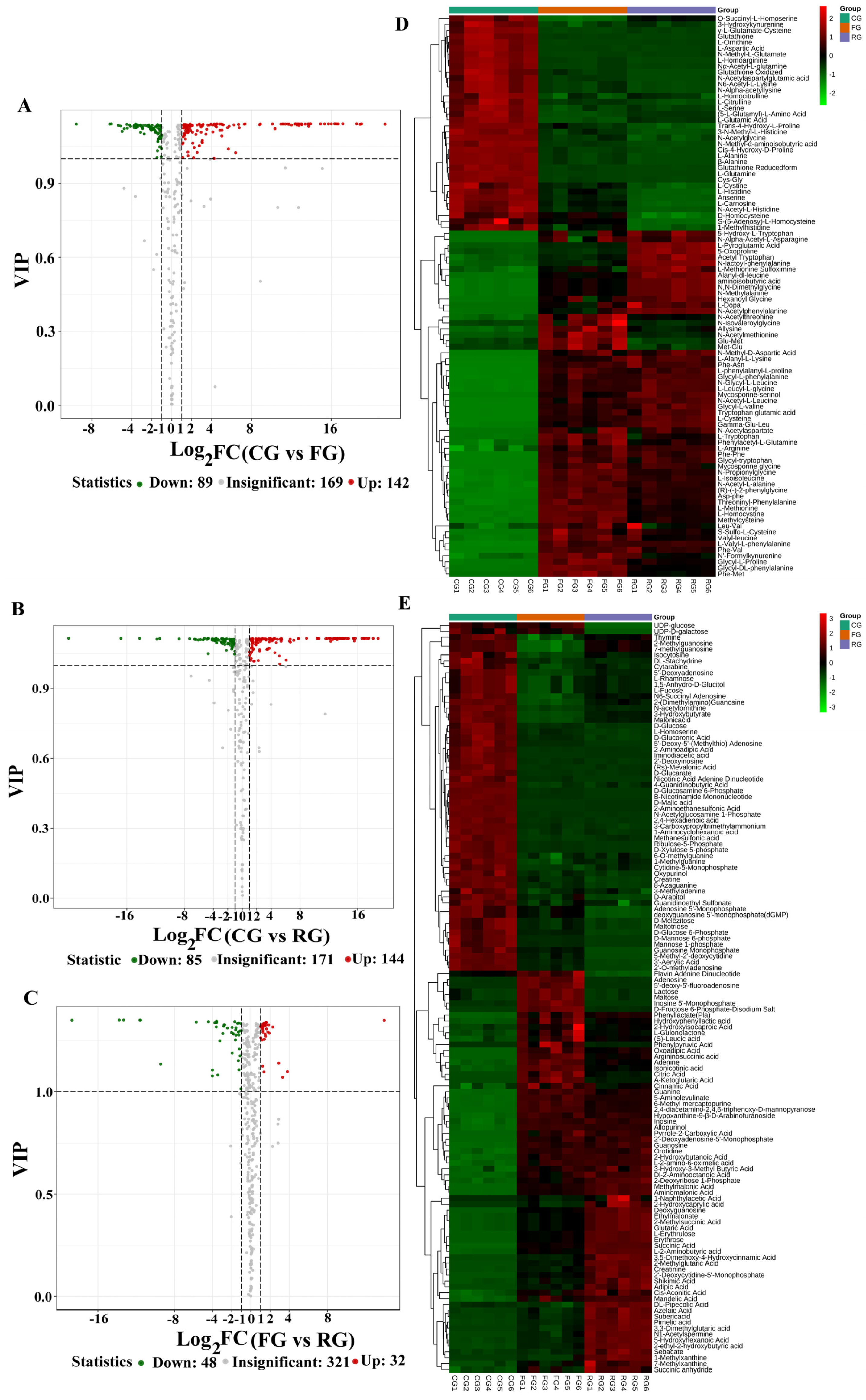
3.4. Identification of Differential Metabolites
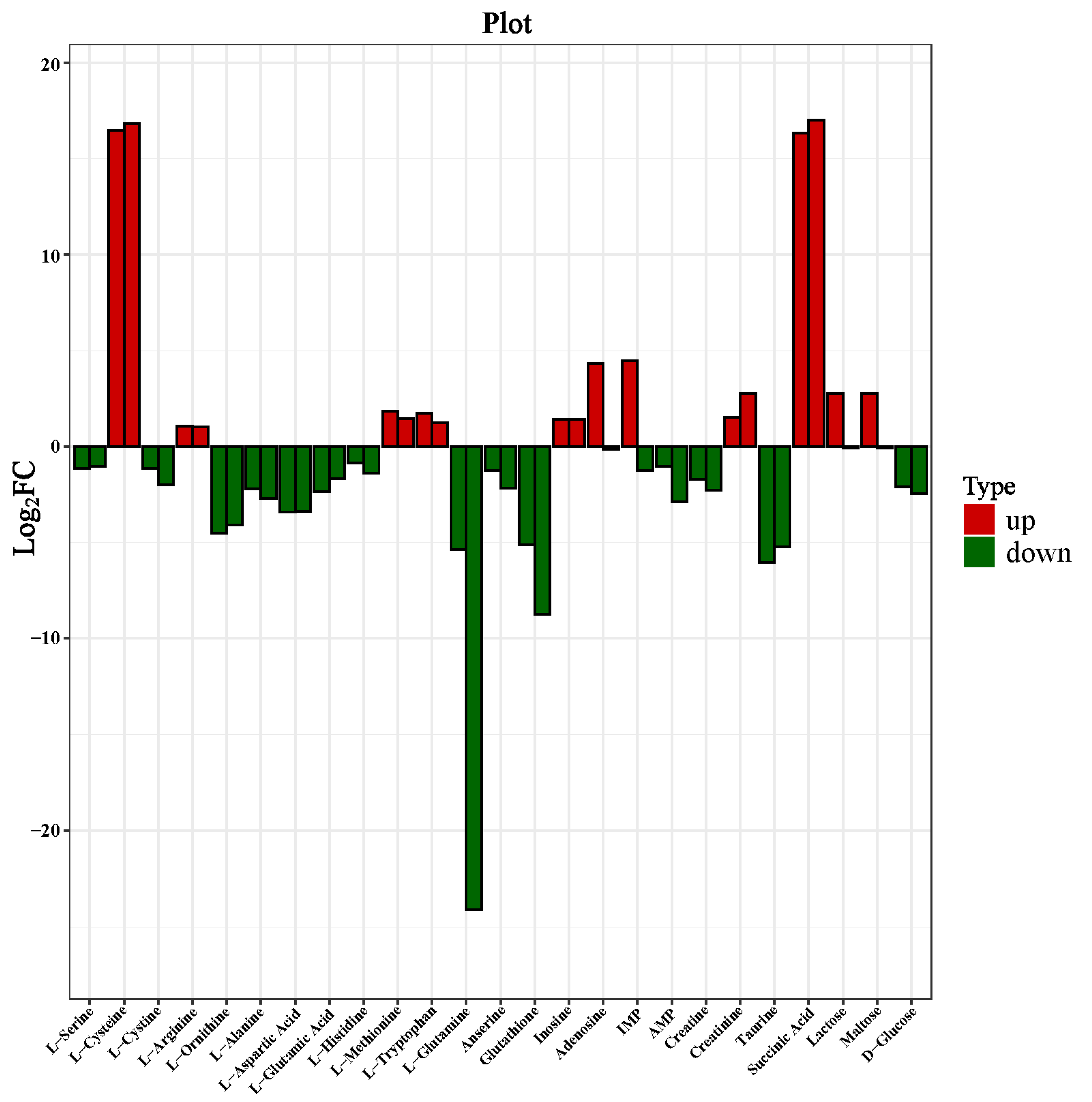
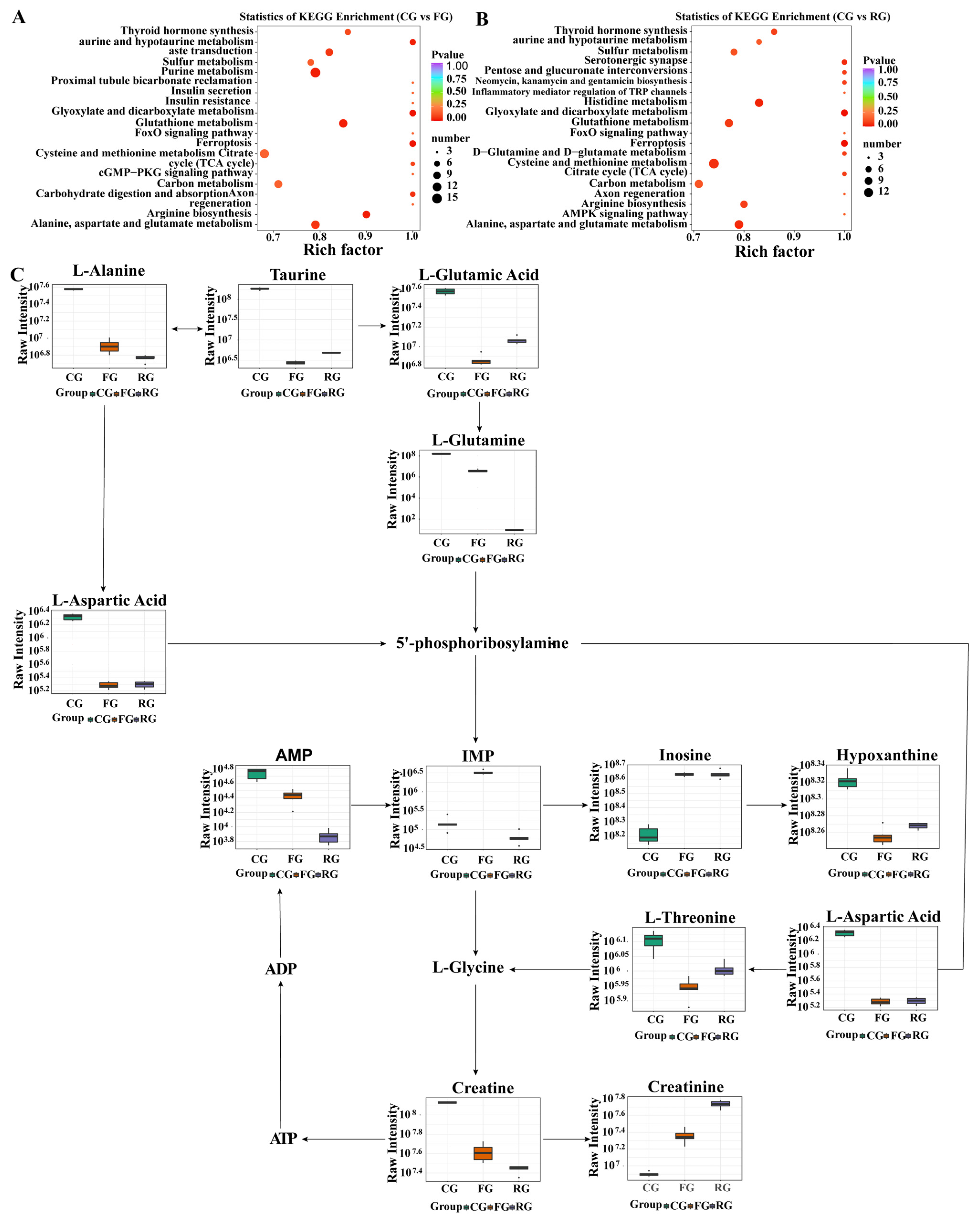
3.5. Changes in Metabolites Composition of Dry-Cured Beef
3.6. Pathway Analysis
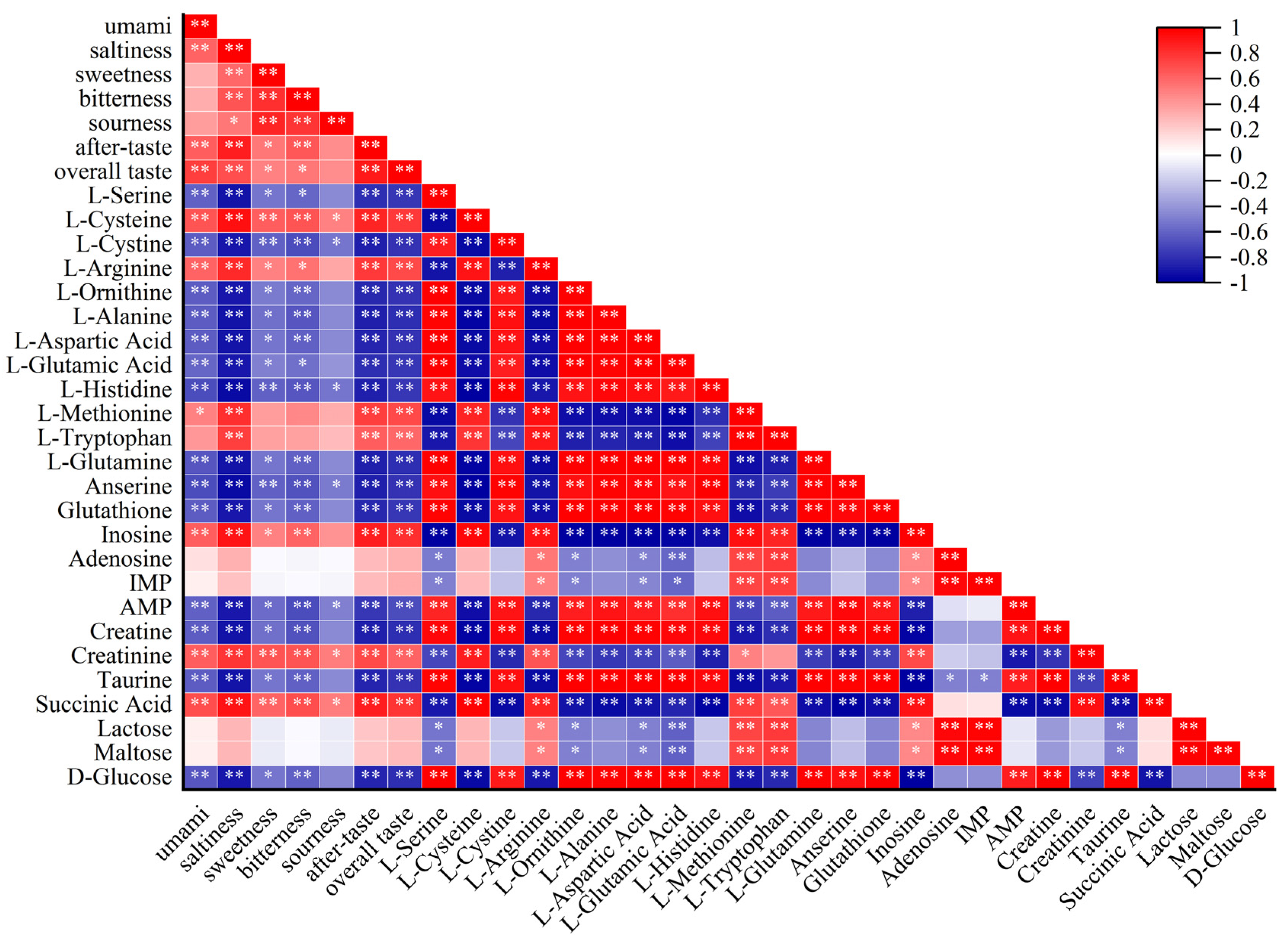
3.7. Analysis of the Correlation between Sensory Assessment and Taste Metabolites
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sha, K.; Lang, Y.M.; Sun, B.Z.; Su, H.W.; Li, H.P.; Zhang, L.; Lei, Y.H.; Li, H.B.; Zhang, Y. Changes in lipid oxidation, fatty acid profile and volatile compounds of traditional kazakh dry-cured beef during processing and storage. J. Food Process. Preserv. 2017, 41, e13059. [Google Scholar] [CrossRef]
- Zhou, C.Y.; Bai, Y.; Wang, C.; Li, C.B.; Xu, X.L.; Pan, D.D.; Cao, J.X.; Zhou, G.H. 1H NMR-based metabolomics and sensory evaluation characterize taste substances of Jinhua ham with traditional and modern processing procedures. Food Control 2021, 126, 107873. [Google Scholar] [CrossRef]
- Suwandy, V.; Carne, A.; Remy, V.D.V.; Bekhit, E.D.A.; Hopkins, D.L. Effect of pulsed electric field on the proteolysis of cold boned beef M. Longissimus lumborum and M. Semimembranosus. Meat Sci. 2015, 100, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Qiao, Y.; Zou, Y.F.; Huang, M.; Kang, Z.L.; Zhou, G.G. Effect of Flavourzyme on proteolysis, antioxidant capacity and sensory attributes of Chinese sausage. Meat Sci. 2014, 98, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.C.; Ge, C.G.; Zhou, G.H.; Zhang, W.G.; Liao, G.Z. H NMR-based metabolic characterization of Chinese Wuding chicken meat. Food Chem. 2019, 274, 574–582. [Google Scholar] [CrossRef]
- Tian, X.; Li, Z.J.; Chao, Y.Z.; Wu, Z.Q.; Zhou, M.X.; Xiao, S.T.; Zeng, J.; Zhe, J. Evaluation by electronic tongue and headspace-GC-IMS analyses of the flavor compounds in dry-cured pork with different salt content. Food Res. Int. 2020, 137, 109456. [Google Scholar] [CrossRef]
- Martín-Gómez, A.; Segura-Borrego, M.P.; Ríos-Reina, R.; Cardador, M.J.; Callejón, R.M.; Morales, M.L.; Rodríguez-Estévezd, V.; Arce, L. Discrimination of defective dry-cured Iberian ham determining volatile compounds by non-destructive sampling and gas chromatography. LWT-Food Sci. Technol. 2022, 154, 112785. [Google Scholar] [CrossRef]
- Karpiński, P.; Kruszewski, B.; Stachelska, M.A.; Szabowska, E. Development of volatiles profile of Kumpiak podlaski dry-cured ham during traditional ripening. Int. J. Food Sci. Technol. 2020, 55, 3630–3638. [Google Scholar] [CrossRef]
- Koutsidis, G.; Elmore, J.S.; Oruna-Concha, M.J.; Campo, M.M.; Wood, J.D.; Mottram, D.S. Water-soluble precursors of beef flavour: I. Effect of diet and breed. Meat Sci. 2008, 79, 124–130. [Google Scholar] [CrossRef]
- Graham, S.F.; Kennedy, T.; Chevallier, O.; Gordon, A.; Farmer, L.; Elliott, C.; Moss, B. The application of NMR to study changes in polar metabolite concentrations in beef longissimus dorsi stored for different periods post mortem. Metabolomics 2010, 6, 395–404. [Google Scholar] [CrossRef]
- Sugimoto, M.; Sugawara, T.; Obiya, S.; Enomoto, A.; Kaneko, M.; Ota, S.; Soga, T.; Tomita, M. Sensory properties and metabolomic profiles of dry-cured ham during the ripening process. Food Res. Int. 2020, 129, 108850. [Google Scholar] [CrossRef] [PubMed]
- Koutsidis, G.; Elmore, J.S.; Oruna-Concha, M.J.; Campo, M.M.; Wood, J.D.; Mottram, D.S. Water-soluble precursors of beef flavour. Part II: Effect of post-mortem conditioning. Meat Sci. 2008, 79, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Lou, X.W.; Ye, Y.F.; Wang, Y.; Sun, Y.Y.; Pan, D.D.; Cao, J.X. Effect of high-pressure treatment on taste and metabolite profiles of ducks with two different vinasse-curing processes. Food Res. Int. 2018, 105, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.Y.; Wang, Y.; Cao, J.X.; Chen, Y.J.; Liu, Y.; Sun, Y.Y.; Pan, D.D.; Ou, C.R. The effect of dry-cured salt contents on accumulation of non-volatile compounds during dry-cured goose processing. Poult. Sci. 2016, 95, 2160–2166. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.Z.; Gonzalez, F.J. LC–MS-based metabolomics: An update. Arch. Toxicol. 2014, 88, 1491–1502. [Google Scholar] [CrossRef]
- Dunn, W.B.; Broadhurst, D.I.; Helen, J.; Atherton, H.J.; Goodacre, R.; Griffin, J.L. Systems level studies of mammalian metabolomes: The roles of mass spectrometry and nuclear magnetic resonance spectroscopy. Chem. Soc. Rev. 2011, 40, 387–426. [Google Scholar] [CrossRef]
- Liao, R.y.; Xia, Q.; Zhou, C.Y.; Geng, F.; Wang, Y.; Sun, Y.Y.; He, J.; Pan, D.D.; Cao, J.X. LC-MS/MS-based metabolomics and sensory evaluation characterize metabolites and texture of normal and spoiled dry-cured hams. Food Chem. 2021, 371, 131156. [Google Scholar] [CrossRef]
- Mabuchi, R.; Ishimaru, A.; Tanaka, M.; Kawaguchi, O.; Tanimoto, S. Metabolic profiling of fish meat by GC-MS analysis, and correlations with taste attributes obtained using an electronic tongue. Metabolites 2019, 9, 1. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Yi, Y.; Pan, D.D.; Zhou, G.H.; Wang, Y.; Dang, Y.L.; He, J.; Li, G.; Cao, J.X. H1-NMR-based metabolomics profiling and taste of boneless dry-cured hams during processing. Food Res. Int. 2019, 122, 114–122. [Google Scholar] [CrossRef]
- Zhang, J.; Ye, Y.F.; Sun, Y.Y.; Pan, D.D.; Ou, C.G.; Dang, Y.L.; Wang, Y.; Cao, J.X.; Wang, D.Y. H1-NMR and multivariate data analysis of the differences of metabolites in five types of dry-cured hams. Food Res. Int. 2018, 113, 140–148. [Google Scholar] [CrossRef]
- Liu, S.X.; Zhang, Y.W.; Zhou, G.H.; Bao, Y.J.; Ren, X.P.; Zhu, Y.X.; Peng, Z.Q. Protein degradation, color and textural properties of low sodium dry cured beef. Int. J. Food Prop. 2019, 22, 487–498. [Google Scholar] [CrossRef]
- Shi, S.; Kong, B.H.; Wang, Y.; Liu, Q.; Xia, X.F. Comparison of the quality of beef jerky processed by traditional and modern drying methods from different districts in Inner Mongolia. Meat Sci. 2020, 163, 108080. [Google Scholar] [CrossRef] [PubMed]
- AOAC (Ed.) Official Methods of Analysis of AOAC International, 16th ed.; Association of Analytical Communities: Arlington, VA, USA, 1995; Volume I–II. [Google Scholar]
- Careri, M.; Mangia, A.; Barbieri, G.; Bouoni, L.; Virgili, R.; Parolari, G. Sensory property relationships to chemical data of italian-type dry-cured ham. J. Food Sci. 1993, 58, 968–972. [Google Scholar] [CrossRef]
- Ran, L.D.; Li, W.H.; Zhao, C.; Zhong, Y.Y.; Yuan, H.C.; Yan, Q.Q.; Zhu, W.C.; Dong, J. Effect of Tea Polyphenol/Hydroxypropyl-β-Cyclodextrin Inclusion Complex on Myofibrillar Protein Oxidation in Ovine Tripe during Refrigerated Storage. Food Sci. 2021, 42, 227–235. [Google Scholar]
- Zhang, B.; Fang, C.D.; Hao, G.J.; Zhang, Y.Y. Effect of kappa-carrageenan oligosaccharides on myofibrillar protein oxidation in peeled shrimp (Litopenaeus vannamei) during long-term frozen storage. Food Chem. 2018, 245, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Jia, D.; You, J.; Hu, Y.; Liu, R.; Xiong, S. Effect of CaCl2 on denaturation and aggregation of silver carp myosin during setting. Food Chem. 2015, 185, 212–218. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, Y.; Pan, D.D.; Zhang, Y.; He, J.; Xia, Q.; Cao, J.X. The application of H NMR to explore the taste difference caused by taste-active metabolites of different Chinese sauce-stewed beef. Food Sci. Nutr. 2020, 8, 4868–4876. [Google Scholar] [CrossRef]
- Xia, J.G.; Sinelnikov, I.V.; Han, B.; Wishart, D.S. MetaboAnalyst 3.0—Making metabolomics more meaningful. Nucleic Acids Res. 2015, 43, 251–257. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.M.; Lu, S.L.; Wang, Q.L. Study on the variation of physicochemical properties during dry-cured mutton ham processing. Food Ind. 2016, 37, 80–88. [Google Scholar]
- Zhou, C.Y.; Pan, D.D.; Bai, Y.; Li, C.B.; Xu, X.L.; Zhou, G.H.; Cao, J.X. Evaluating endogenous protease of salting exudates during the salting process of Jinhua ham. LWT-Food Sci. Technol. 2019, 101, 76–82. [Google Scholar] [CrossRef]
- Martuscelli, M.; Lupieri, L.; Chaves-Lopez, C.; Mastrocola, D.; Pittia, P. Technological approach to reduce NaCl content of traditional smoked dry-cured hams: Effect on quality properties and stability. J. Food Sci. Technol. 2015, 52, 7771–7782. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.M.; Lu, S.L.; Li, K.X.; Jiang, C.H.; Peng, X.L.; Pei, L.Y.; Wang, Y.H.; Chu, X.H. Study on proteolysis during processing of dry-cured mutton ham. Food Ferment. Ind. 2014, 40, 238–242. [Google Scholar]
- Zhou, G.H.; Zhao, G.M. Biochemical changes during processing of traditional Jinhua ham. Meat Sci. 2007, 77, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Xiong, Y. Comparative time-course of lipid and myofibrillar protein oxidation in different biphasic systems under hydroxyl radical stress. Food Chem. 2018, 243, 231–238. [Google Scholar] [CrossRef]
- Saiga, A.; Tanabe, S.; Nishimura, T. Antioxidant activity of peptides obtained from porcine myofibrillar proteins by protease treatment. J. Agric. Food Chem. 2003, 51, 3661–3667. [Google Scholar] [CrossRef]
- Sforza, S.; Galaverna, G.; Schivazappa, C.; Marchelli, R.; Dossena, A.; Virgili, R. Effect of extended aging of Parma dry-cured ham on the content of oligopeptides and free amino acids. J. Agric. Food Chem. 2006, 54, 9422–9429. [Google Scholar] [CrossRef]
- Zhao, G.M.; Zhou, G.H.; Tian, W.; Xu, X.L.; Wang, Y.L.; Luo, X. Changes of alanyl aminopeptidase activity and free amino acid contents in biceps femoris during processing of Jinhua ham. Meat Sci. 2005, 71, 612–619. [Google Scholar] [CrossRef]
- Dang, Y.L.; Wang, Z.; Xu, S.Y. Methods for extracting the taste compounds from water soluble extract of Jinhua ham. Eur. Food Res. Technol. 2008, 228, 93–102. [Google Scholar] [CrossRef]
- Feng, X.; Hang, S.S.; Zhou, Y.G.; Liu, Q.; Yang, H.S. Bromelain Kinetics and Mechanism on Myofibril from Golden Pomfret (Trachinotus blochii). J. Food Sci. 2018, 83, 2148–2158. [Google Scholar] [CrossRef]
- Zhao, X.; Wu, J.; Chen, L.; Yang, H.S. Effect of vacuum impregnated fish gelatin and grape seed extract on metabolite profiles of tilapia (Oreochromis niloticus) fillets during storage. Food Chem. 2019, 293, 418–428. [Google Scholar] [CrossRef]
- Liu, S.Y.; Wang, G.Y.; Xiao, Z.C.; Pu, Y.H.; Ge, C.R.; Liao, G.Z. 1H-NMR-based water-soluble low molecular weight compound characterization and free fatty acid composition of five kinds of Yunnan dry-cured hams. LWT-Food Sci. Technol. 2019, 108, 174–182. [Google Scholar] [CrossRef]
- Ramanathan, R.; Mancini, R.A.; Dady, G.A.; Van, B.C.B. Effects of succinate and pH on cooked beef color. Meat Sci. 2013, 93, 888–892. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Y.; Li, C.S.; Li, L.H.; Yang, X.Q.; Chen, S.J.; Wu, Y.Y.; Zhao, Y.Q.; Wang, J.X.; Wei, Y.; Yang, D.Q. Application of UHPLC-Q/TOF-MS-based metabolomics in the evaluation of metabolites and taste quality of Chinese fish sauce (yu-lu) during fermentation. Food Chem. 2019, 296, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Sundekilde, U.K.; Rasmussen, M.K.; Young, J.F.; Bertram, H.C. High resolution magic angle spinning NMR spectroscopy reveals that pectoralis muscle dystrophy in chicken is associated with reduced muscle content of anserine and carnosine. Food Chem. 2017, 217, 151–154. [Google Scholar] [CrossRef]
- Park, J.N.; Watanabe, T.; Endoh, K.I.; Watanabe, K.; Abe, H. Taste-active components in a Vietnamese fish sauce. Fish. Sci. 2002, 68, 913–920. [Google Scholar] [CrossRef]
- Hong, H.; Regenstein, J.M.; Luo, Y.K. The importance of ATP-related compounds for the freshness and flavor of post-mortem fish and shellfish muscle: A review. Crit. Rev. Food Sci. Nutr. 2017, 57, 1787–1798. [Google Scholar] [CrossRef]
- Yang, Y.; Ye, Y.F.; Pan, D.D.; Sun, Y.Y.; Wang, Y.; Cao, J.X. Metabonomics profiling of marinated meat in soy sauce during processing. J. Sci. Food Agric. 2018, 98, 1325–1331. [Google Scholar] [CrossRef]
- Guo, Y.R.; Gu, S.Q.; Wang, X.C.; Zhuang, K.J.; Wang, S.; Shi, J. Nutrients and non-volatile taste compounds in Chinese mitten crab by-products. Fish. Sci. 2015, 81, 193–203. [Google Scholar] [CrossRef]
- Zou, Y.H.; Kang, D.C.; Liu, R.; Qi, J.; Zhou, G.H.; Zhang, W.G. Effects of ultrasonic assisted cooking on the chemical profiles of taste and flavor of spiced beef. Ultrason. Sonochem. 2018, 46, 36–45. [Google Scholar] [CrossRef]
- Chen, D.A.; Ye, Y.F.; Chen, J.J.; Zhan, P.P.; Lou, Y.J. Molecular nutritional characteristics of vinasse pike eel (Muraenesox cinereus) during pickling. Food Chem. 2017, 224, 359–364. [Google Scholar] [CrossRef]
- Kang, S.M.; Kang, G.H.; Seong, P.; Kim, Y.C.; Park, B.Y.; Cho, S.H. Effect of Cooking Condition on the Water-Soluble Flavor Precursors in Various Beef Muscles from Hanwoo (Korean Cattle). Korean J. Food Sci. Anim. Resour. 2013, 33, 752–756. [Google Scholar] [CrossRef] [Green Version]
- Klamt, S.; Stelling, J.B. Two approaches for metabolic pathway analysis? Trends Biotechnol. 2003, 21, 64–69. [Google Scholar] [CrossRef]
- Cônsolo, N.R.B.; Rosa, A.F.; Barbosa, L.C.G.S.; Maclean, P.H.; Higuera, P.A.; Colnago, L.A.; Titto, E.A.L. Preliminary study on the characterization of Longissimus lumborum dark cutting meat in Angus × Nellore crossbreed cattle using NMR-based metabolomics. Meat Sci. 2021, 172, 108350. [Google Scholar] [CrossRef]
- Franciosa, I.; Ferrocino, I.; Giordano, M.; Mounier, J.; Rantsiou, K.; Cocolin, L. Specific metagenomic asset drives the spontaneous fermentation of Italian sausages. Food Res. Int. 2021, 144, 110379. [Google Scholar] [CrossRef] [PubMed]
- Huang, S. Integrating Transcriptomics and Metabolomics to Reveal Metabolism of Amino Acid of Streptococcus thermophilus TF96; Northeast Agricultural University: Harbin City, China, 2017. [Google Scholar]
- Sun, H.W.; Wang, J.Z.; Zhang, C.H.; Li, X.; Xu, X.; Dong, X.B.; Hu, L.; Li, C.H. Changes of flavor compounds of hydrolyzed chicken bone extracts during Maillard reaction. J. Food Sci. 2014, 79, 2415–2426. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Pan, D.D.; Wang, Y.; He, J.; Yue, Y.; Xia, Q.; Zhou, G.H.; Cao, J.X. Effect of reconstituted broth on the taste-active metabolites and sensory quality of stewed and roasted pork-hock. Foods 2020, 9, 513. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CG | FG | RG | |
|---|---|---|---|
| Moisture (%) | 74.71 ± 0.68 aA | 34.93 ± 0.42 bB | 25.43 ± 0.22 cC |
| pH value | 5.83 ± 0.03 cC | 5.91 ± 0.01 bB | 6.1 ± 0.02 aA |
| NaCl (%) | 0.05 ± 0.01 cC | 10.36 ± 0.28 bB | 13.56 ± 0.34 aA |
| Total nitrogen (%) | 8.98 ± 0.40 cC | 9.23 ± 0.52 bB | 10.42 ± 0.49 aA |
| Non-protein nitrogen (%) | 0.67 ± 0.64 cC | 1.18 ± 0.35 bB | 1.81 ± 0.18 aA |
| Proteolysis index (%) | 7.46 ± 0.48 cC | 12.82 ± 0.32 bB | 17.34 ± 0.15 aA |
| SP Carbonyl content (nmol/mg) | 5.36 ± 0.01 cC | 8.52 ± 0.11 bB | 11.53 ± 0.18 aA |
| MP Carbonyl content (nmol/mg) | 11.56 ± 0.07 cC | 14.97 ± 0.16 bB | 18.3 ± 0.13 aA |
| SP Sulfhydryl content (nmol/mg) | 41.05 ± 0.51 aA | 28.43 ± 0.26 bB | 24.34 ± 0.37 cC |
| MP Sulfhydryl content (nmol/mg) | 32.13 ± 0.45 aA | 19.71 ± 0.22 bB | 16.3 ± 0.31 cC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, H.; Pan, L.; Wang, J.; Zhao, J.; Guo, X.; Chen, J.; Lu, S.; Dong, J.; Wang, Q. Sensory Properties and Main Differential Metabolites Influencing the Taste Quality of Dry-Cured Beef during Processing. Foods 2022, 11, 531. https://doi.org/10.3390/foods11040531
Fu H, Pan L, Wang J, Zhao J, Guo X, Chen J, Lu S, Dong J, Wang Q. Sensory Properties and Main Differential Metabolites Influencing the Taste Quality of Dry-Cured Beef during Processing. Foods. 2022; 11(4):531. https://doi.org/10.3390/foods11040531
Chicago/Turabian StyleFu, Huihui, Li Pan, Jingyun Wang, Jixing Zhao, Xin Guo, Jingya Chen, Shiling Lu, Juan Dong, and Qingling Wang. 2022. "Sensory Properties and Main Differential Metabolites Influencing the Taste Quality of Dry-Cured Beef during Processing" Foods 11, no. 4: 531. https://doi.org/10.3390/foods11040531
APA StyleFu, H., Pan, L., Wang, J., Zhao, J., Guo, X., Chen, J., Lu, S., Dong, J., & Wang, Q. (2022). Sensory Properties and Main Differential Metabolites Influencing the Taste Quality of Dry-Cured Beef during Processing. Foods, 11(4), 531. https://doi.org/10.3390/foods11040531