
Microbial Communities and Flavor Compounds during the Fermentation of Traditional Hong Qu Glutinous Rice Wine

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation and Collection
2.2. Physiochemical Properties Determination
2.3. Total DNA Extraction and PCR Amplification
2.4. Sequence Analysis
2.5. Analysis of VFCs
2.6. Analysis of Free Amino Acids
2.7. Analysis of Organic Acids
2.8. Statistical Analysis
3. Results and Discussion
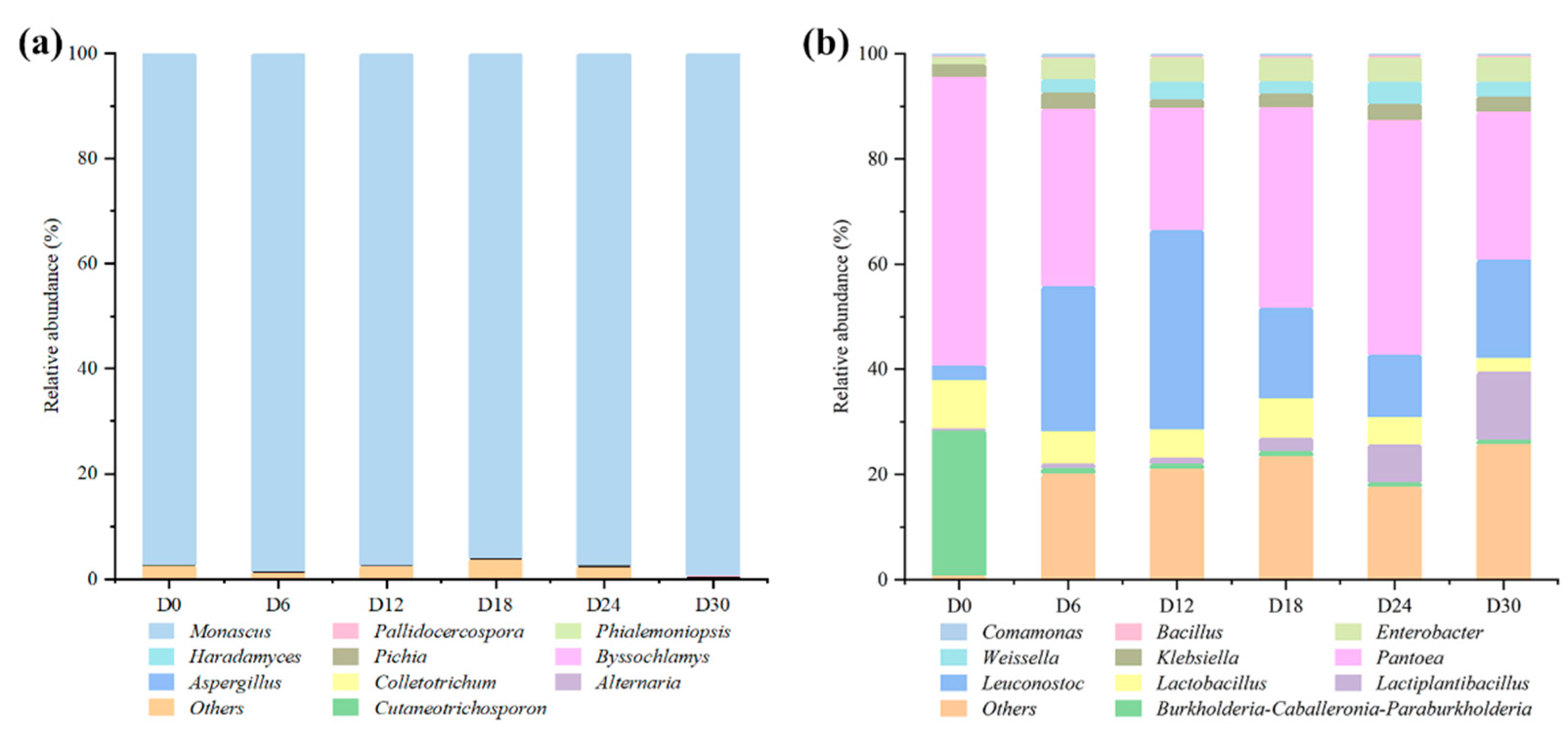
3.1. Analysis of the Composition of Microbial Communities
3.2. Physicochemical Properties and Flavor Compounds
3.2.1. Physicochemical Properties Analysis
3.2.2. Changes in the Free Amino Acid Fraction of HQW during Fermentation
3.2.3. Changes in Organic Acids
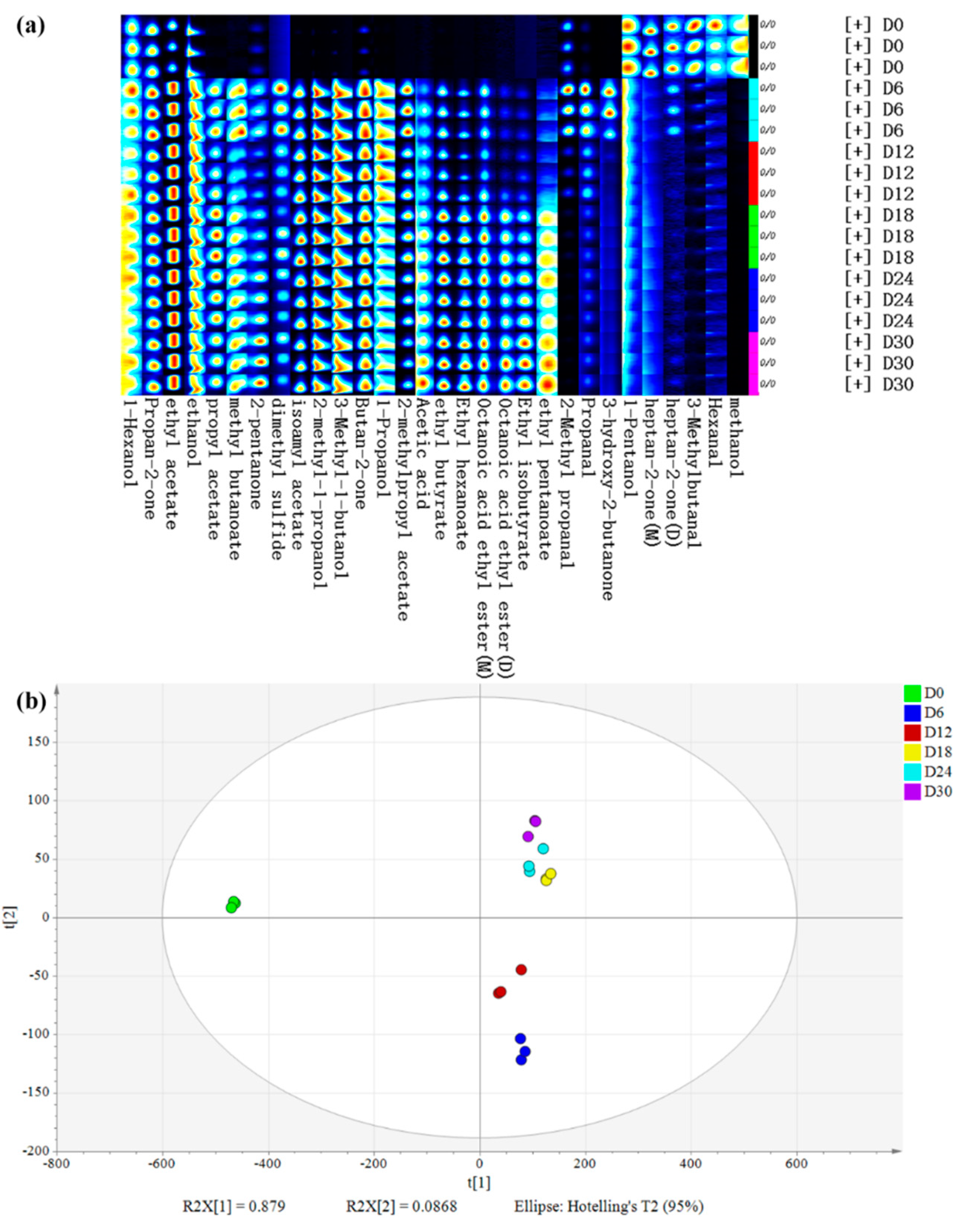
3.2.4. Volatile Flavor Compounds Analysis
3.3. Correlation Analysis of Microbial Communities and Flavor Compounds
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Huang, Y.Y.; Liang, Z.C.; Lin, X.Z.; He, Z.G.; Ren, X.Y.; Li, W.X.; Molnar, I. Fungal community diversity and fermentation characteristics in regional varieties of traditional fermentation starters for Hong Qu glutinous rice wine. Food Res. Int. 2021, 141, 110146. [Google Scholar] [CrossRef]
- Yang, C.L.; Wu, X.P.; Chen, B.; Deng, S.S.; Chen, Z.E.; Huang, Y.Y.; Jin, S.S. Comparative analysis of genetic polymorphisms among Monascus strains by ISSR and RAPD markers. J. Sci. Food Agric. 2017, 97, 636–640. [Google Scholar] [CrossRef] [PubMed]
- Gänzle, M.G. Lactic metabolism revisited: Metabolism of lactic acid bacteria in food fermentations and food spoilage. Curr. Opin. Food Sci. 2015, 2, 106–117. [Google Scholar] [CrossRef]
- Ye, M.Q.; Yue, T.L.; Yuan, Y.H. Evolution of polyphenols and organic acids during the fermentation of apple cider. J. Sci. Food Agric. 2014, 94, 2951–2957. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Huang, J.; Xie, T.; Huang, L.; Zhuang, W.; Zheng, Y.; Zheng, B. Oenological characteristics, amino acids and volatile profiles of Hongqu rice wines during pottery storage: Effects of high hydrostatic pressure processing. Food Chem. 2016, 203, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Luo, T.; Fan, W.L.; Xu, Y. Characterization of volatile and semi-volatile compounds in Chinese rice wines by headspace solid phase microextraction followed by gas chromatography-mass spectrometry. J. Inst. Brew. 2008, 114, 172–179. [Google Scholar] [CrossRef]
- Jiang, L.; Su, W.; Mu, Y.; Mu, Y. Major Metabolites and Microbial Community of Fermented Black Glutinous Rice Wine With Different Starters. Front. Microbiol. 2020, 11, 593. [Google Scholar] [CrossRef]
- Huang, Z.R.; Hong, J.L.; Xu, J.X.; Li, L.; Guo, W.L.; Pan, Y.Y.; Lv, X.C. Exploring core functional microbiota responsible for the production of volatile flavour during the traditional brewing of Wuyi Hong Qu glutinous rice wine. Food Microbiol. 2018, 76, 487–496. [Google Scholar] [CrossRef]
- Mayo, B.; Rachid, C.; Alegria, A.; Leite, A.M.O.; Peixoto, R.S.; Delgado, S. Impact of Next Generation Sequencing Techniques in Food Microbiology. Curr. Genom. 2014, 15, 293–309. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Su, W.; Mu, Y.; Jiang, L.; Mu, Y. Correlations between microbiota with physicochemical properties and volatile flavor components in black glutinous rice wine fermentation. Food Res. Int. 2020, 138, 109800. [Google Scholar] [CrossRef]
- Wang, P.; Mao, J.; Meng, X.; Li, X.; Liu, Y.; Feng, H. Changes in flavour characteristics and bacterial diversity during the traditional fermentation of Chinese rice wines from Shaoxing region. Food Control 2014, 44, 58–63. [Google Scholar] [CrossRef]
- Guo, L.; Luo, Y.; Zhou, Y.; Bianba, C.; Guo, H.; Zhao, Y.; Fu, H. Exploring microbial dynamics associated with flavours production during highland barley wine fermentation. Food Res. Int. 2020, 130, 108971. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Lin, C.; Zhang, W.; Yang, Q.; Meng, J.; He, L.; Zeng, X. Exploring the bacterial community for starters in traditional high-salt fermented Chinese fish (Suanyu). Food Chem. 2021, 358, 129863. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wang, B.; Fu, Y.; Shi, Y.; Chen, F.; Guan, H.; Zhang, N. HS-GC-IMS with PCA to analyze volatile flavor compounds across different production stages of fermented soybean whey tofu. Food Chem. 2021, 346, 128880. [Google Scholar] [CrossRef]
- Chen, W.; He, Y.; Zhou, Y.; Shao, Y.; Feng, Y.; Li, M.; Chen, F. Edible Filamentous Fungi from the Species Monascus: Early Traditional Fermentations, Modern Molecular Biology, and Future Genomics. Compr. Rev. Food Sci. Food Saf. 2015, 14, 555–567. [Google Scholar] [CrossRef] [Green Version]
- Liang, Z.; Lin, X.; He, Z.; Su, H.; Li, W.; Ren, X. Amino acid and microbial community dynamics during the fermentation of Hong Qu glutinous rice wine. Food Microbiol. 2020, 90, 103467. [Google Scholar] [CrossRef]
- Zheng, J.S.; Wittouck, S.; Salvetti, E.; Franz, C.; Harris, H.M.B.; Mattarelli, P.; Lebeer, S. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Liu, X.; Qian, M.; Shen, Y.; Qin, X.; Huang, H.; Yang, H.; Bai, W. An high-throughput sequencing approach to the preliminary analysis of bacterial communities associated with changes in amino acid nitrogen, organic acid and reducing sugar contents during soy sauce fermentation. Food Chem. 2021, 349, 129131. [Google Scholar] [CrossRef]
- Li, W.W.; Fan, G.S.; Fu, Z.L.; Wang, W.H.; Xu, Y.Q.; Teng, C.; Li, X.T. Effects of fortification of Daqu with various yeasts on microbial community structure and flavor metabolism. Food Res. Int. 2020, 129, 10. [Google Scholar] [CrossRef]
- González, Á.; Hierro, N.; Poblet, M.; Mas, A.; Guillamón, J.M. Application of molecular methods to demonstrate species and strain evolution of acetic acid bacteria population during wine production. Int. J. Food Microbiol. 2005, 102, 295–304. [Google Scholar] [CrossRef]
- Sun, L.; Bai, C.; Xu, H.; Na, N.; Jiang, Y.; Yin, G.; Xue, Y. Succession of Bacterial Community During the Initial Aerobic, Intense Fermentation, and Stable Phases of Whole-Plant Corn Silages Treated with Lactic Acid Bacteria Suspensions Prepared From Other Silages. Front Microbiol. 2021, 12, 655095. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Zhang, T.; Zhang, Q.; Luo, J.; Cai, C.; Mao, J. Microbial diversity and chemical analysis of the starters used in traditional Chinese sweet rice wine. Food Microbiol. 2018, 73, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Zhou, Y.; Li, J.; Liu, S.; He, S.; Sun, H.; Xu, S. Effect of enzymes addition on the fermentation of Chinese rice wine using defined fungal starter. LWT 2021, 143, 111101. [Google Scholar] [CrossRef]
- Tian, T.; Sun, J.; Wu, D.; Xiao, J.; Lu, J. Objective measures of greengage wine quality: From taste-active compound and aroma-active compound to sensory profiles. Food Chem. 2021, 340, 128179. [Google Scholar] [CrossRef] [PubMed]
- El-Dalatony, M.M.; Saha, S.; Govindwar, S.P.; Abou-Shanab, R.A.I.; Jeon, B.H. Biological Conversion of Amino Acids to Higher Alcohols. Trends Biotechnol. 2019, 37, 855–869. [Google Scholar] [CrossRef]
- Mato, I.; Suárez-Luque, S.; Huidobro, J.F. A review of the analytical methods to determine organic acids in grape juices and wines. Food Res. Int. 2005, 38, 1175–1188. [Google Scholar] [CrossRef]
- Sumby, K.M.; Grbin, P.R.; Jiranek, V. Microbial modulation of aromatic esters in wine: Current knowledge and future prospects. Food Chem. 2010, 121, 1–16. [Google Scholar] [CrossRef]
- Ye, Y.T.; Wang, L.X.; Zhan, P.; Tian, H.L.; Liu, J.S. Characterization of the aroma compounds of Millet Huangjiu at different fermentation stages. Food Chem. 2022, 366, 10. [Google Scholar] [CrossRef]
- Yang, Y.; Hu, W.; Xia, Y.; Mu, Z.; Tao, L.; Song, X.; Zhang, H.; Ni, B.; Ai, L. Flavor Formation in Chinese Rice Wine (Huangjiu): Impacts of the Flavor-Active Microorganisms, Raw Materials, and Fermentation Technology. Front. Microbiol. 2020, 11, 2541. [Google Scholar] [CrossRef]
- Larralde-Corona, C.P.; De la Torre-Gonzalez, F.J.; Vazquez-Landaverde, P.A.; Hahn, D.; Narvaez-Zapata, J.A. Rational Selection of Mixed Yeasts Starters for Agave Must Fermentation. Front. Sustain. Food Syst. 2021, 5, 13. [Google Scholar] [CrossRef]
- Yang, Y.J.; Xia, Y.J.; Song, X.; Mu, Z.Y.; Qiu, H.Z.; Tao, L.R.; Ai, L.Z. The Potential of Flos sophorae immaturus as a Pigment-Stabilizer to Improve the Monascus Pigments Preservation, Flavor Profiles, and Sensory Characteristic of Hong Qu Huangjiu. Front. Microbiol. 2021, 12, 678903. [Google Scholar] [CrossRef] [PubMed]
- El Hadi, M.A.; Zhang, F.J.; Wu, F.F.; Zhou, C.H.; Tao, J. Advances in fruit aroma volatile research. Molecules 2013, 18, 8200–8229. [Google Scholar] [CrossRef]
- Chen, C.; Liu, Z.; Zhou, W.Y.; Tian, H.X.; Huang, J.; Yuan, H.B.; Yu, H.Y. Comparison of the Fermentation Activities and Volatile Flavor Profiles of Chinese Rice Wine Fermented Using an Artificial Starter, a Traditional JIUYAO and a Commercial Starter. Front. Microbiol. 2021, 12, 716281. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.Y.; Xie, T.; Xie, J.R.; Ai, L.Z.; Tian, H.X. Characterization of key aroma compounds in Chinese rice wine using gas chromatography-mass spectrometry and gas chromatography-olfactometry. Food Chem. 2019, 293, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Contador, R.; Delgado, F.J.; Garcia-Parra, J.; Garrido, M.; Ramirez, R. Volatile profile of breast milk subjected to high-pressure processing or thermal treatment. Food Chem. 2015, 180, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Bueno, M.; Carrascon, V.; Ferreira, V. Release and Formation of Oxidation-Related Aldehydes during Wine Oxidation. J. Agric. Food Chem. 2016, 64, 608–617. [Google Scholar] [CrossRef] [Green Version]
- Yuan, F.; He, F.; Qian, Y.; Zheng, J.; Qian, M.C. Aroma Stability of Lemon-Flavored Hard Iced Tea Assessed by Chirality and Aroma Extract Dilution Analysis. J. Agric. Food Chem. 2016, 64, 5717–5723. [Google Scholar] [CrossRef]
- Gänzle, M.G.; Vermeulen, N.; Vogel, R.F. Carbohydrate, peptide and lipid metabolism of lactic acid bacteria in sourdough. Food Microbiol. 2007, 24, 128–138. [Google Scholar] [CrossRef]
- Cocaign-Bousquet, M.; Garrigues, C.; Loubiere, P.; Lindley, N.D. Physiology of pyruvate metabolism in Lactococcus lactis. Antonie Van Leeuwenhoek 1996, 70, 253–267. [Google Scholar] [CrossRef]
- Ganzle, M.G.; Follador, R. Metabolism of oligosaccharides and starch in lactobacilli: A review. Front. Microbiol. 2012, 3, 340. [Google Scholar] [CrossRef] [Green Version]
- Rabinowitz, J.D.; Enerback, S. Lactate: The ugly duckling of energy metabolism. Nat. Metab. 2020, 2, 566–571. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, J.S.; Seeras, A.; Sanchez-Maldonado, A.F.; Zhang, C.; Su, M.S.-W.; Gänzle, M.G. Glutamine, glutamate, and argi-nine-based acid resistance in Lactobacillus reuteri. Food Microbiol. 2014, 42, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Shang, Z.; Ye, Z.; Li, M.; Ren, H.; Cai, S.; Hu, X.; Yi, J. Dynamics of microbial communities, flavor, and physicochemical properties of pickled chayote during an industrial-scale natural fermentation: Correlation between microorganisms and metabolites. Food Chem. 2022, 377, 132004. [Google Scholar] [CrossRef]
- Hortensia Silla Santos, M. Amino acid decarboxylase capability of microorganisms isolated in Spanish fermented meat products. Int. J. Food Microbiol. 1998, 39, 227–230. [Google Scholar] [CrossRef]
- Pyo, Y.-H. Effect of Monascus-fermentation on the Content of GABA and Free Amino Acids in Soybean. J. Korean Soc. Food Sci. Nutr. 2008, 37, 1208–1213. [Google Scholar] [CrossRef]
- Han, D.M.; Chun, B.H.; Kim, H.M.; Jeon, C.O. Characterization and correlation of microbial communities and metabolite and volatile compounds in doenjang fermentation. Food Res. Int. 2021, 148, 110645. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.; Jeong, D.W.; Heo, G.; Kong, H.; Kim, C.T.; Lee, J.H. Genetic Background Behind the Amino Acid Profiles of Fermented Soybeans Produced by Four Bacillus spp. J. Microbiol. Biotechnol. 2021, 31, 447–455. [Google Scholar] [CrossRef]
- Xiao, Y.; Huang, T.; Huang, C.; Hardie, J.; Peng, Z.; Xie, M.; Xiong, T. The microbial communities and flavour compounds of Jiangxi yancai, Sichuan paocai and Dongbei suancai: Three major types of traditional Chinese fermented vegetables. LWT 2020, 121, 108865. [Google Scholar] [CrossRef]
- Dorokhov, Y.L.; Shindyapina, A.V.; Sheshukova, E.V.; Komarova, T.V. Metabolic Methanol: Molecular Pathways and Physiological Roles. Physiol. Rev. 2015, 95, 603–644. [Google Scholar] [CrossRef]
- Blagden, T.D.; Gilliland, S.E. Reduction of levels of volatile components associated with the “Beany” flavor in soymilk by Lactobacilli and Streptococci. J. Food Sci. 2005, 70, M186–M189. [Google Scholar] [CrossRef]
- Alegría, Á.; Delgado, S.; Flórez, A.B.; Mayo, B. Identification, typing, and functional characterization of Leuconostoc spp. strains from traditional, starter-free cheeses. Dairy Sci. Technol. 2013, 93, 657–673. [Google Scholar] [CrossRef] [Green Version]
- Xiang, W.L.; Zhang, N.D.; Lu, Y.; Zhao, Q.H.; Xu, Q.; Rao, Y.; Zhang, Q. Effect of Weissella cibaria co-inoculation on the quality of Sichuan Pickle fermented by Lactobacillus plantarum. LWT 2020, 121, 7. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Physicochemical Index | D0 | D6 | D12 | D18 | D24 | D30 |
|---|---|---|---|---|---|---|
| pH | 5.83 ± 0.01 a | 3.86 ± 0.02 c | 3.90 ± 0.01 b | 3.85 ± 0.01 c | 3.79 ± 0.02 d | 3.84 ± 0.01 c |
| Total sugar (g/L) | 14.36 ± 0.17 b | 48.70 ± 1.20 a | 3.47 ± 0.17 c | 3.21 ± 0.80 c | 2.29 ± 0.15 cd | 1.56 ± 0.08 d |
| Alcohol (v/v, %) | 0.18 ± 0.02 e | 7.33 ± 1.10 d | 14.00 ± 0.28 c | 17.20 ± 0.30 b | 18.80 ± 0.87 a | 19.77 ± 0.15 a |
| Amino acid nitrogen (g/L) | 0.10 ± 0.004 f | 0.14 ± 0.01 e | 0.33 ± 0.02 d | 0.43 ± 0.004 c | 0.54 ± 0.004 b | 0.65 ± 0.01 a |
| Amino Acid (mg/L) | D0 | D6 | D12 | D18 | D24 | D30 |
|---|---|---|---|---|---|---|
| Umami | ||||||
| Asp | 6.40 ± 0.12 f | 20.31 ± 0.15 e | 79.83 ± 2.06 d | 108.34 ± 2.25 c | 142.99 ± 1.42 b | 161.20 ± 2.11 a |
| Glu | 31.24 ± 0.53 e | 26.41 ± 0.12 e | 92.7 ± 1.92 d | 323.69 ± 9.55 c | 370.19 ± 5.43 b | 392.54 ± 7.64 a |
| TUAA | 37.64 | 46.72 | 172.53 | 432.03 | 513.18 | 553.74 |
| Sweet | ||||||
| Ser | 10.38 ± 0.16 e | 7.39 ± 0.09 f | 36.12 ± 0.93 d | 56.77 ± 1.04 c | 77.35 ± 1.29 b | 94.16 ± 2.64 a |
| Gly | 3.13 ± 0.07 c,d | 1.49 ± 0.01 e | 8.04 ± 0.25 b | 11.91 ± 0.25 a | 3.49 ± 0.64 c | 2.59 ± 0.08 d |
| Thr | 12.43 ± 0.27 f | 44.98 ± 0.25 e | 145.62 ± 3.23 d | 195.54 ± 3.98 c | 226.90 ± 3.98 b | 247.38 ± 5.50 a |
| Met | 7.21 ± 0.74 f | 15.97 ± 0.21 e | 22.71 ± 0.30 d | 53.72 ± 0.88 c | 81.95 ± 1.08 b | 85.64 ± 1.59 a |
| Ala | 1.43 ± 0.08 e | 3.26 ± 0.22 e | 7.49 ± 0.20 d | 26.98 ± 0.94 c | 67.06 ± 1.27 b | 75.28 ± 1.98 a |
| Pro | 31.05 ± 3.57 f | 113.86 ± 2.46 e | 228.6 ± 8.42 c | 247.15 ± 5.14 c | 274.8 ± 14.08 b | 306.4 ± 7.01 a |
| TSAA | 34.58 | 73.00 | 219.98 | 344.91 | 456.70 | 505.05 |
| Bitter | ||||||
| His | 13.08 ± 0.19 f | 35 ± 0.34 e | 95.93 ± 1.57 d | 139.9 ± 4.04 c | 185.57 ± 3.97 b | 218.86 ± 6.10 a |
| Arg | 46 ± 0.76 f | 127.77 ± 0.87 e | 261.12 ± 5.7 d | 377.87 ± 7.37 c | 481.23 ± 8.74 b | 518.08 ± 12.91 a |
| Lys | 1.54 ± 0.70 c | 1.71 ± 0.31 c | 1.29 ± 0.48 c | 2.59 ± 0.26 b | 3.9 ± 0.23 a | 4.63 ± 0.10 a |
| Val | 4.03 ± 0.07 e | 3.1 ± 0.09 e | 26.86 ± 0.64 d | 50.63 ± 0.98 c | 73.92 ± 1.07 b | 91.02 ± 2.43 a |
| Trp | 50.46 ± 1.51 f | 132.71 ± 0.53 e | 475.03 ± 10.21 d | 613.99 ± 13.01 c | 769.46 ± 9.59 b | 871.73 ± 14.36 a |
| Phe | 4.87 ± 0.15 f | 12.81 ± 0.05 e | 45.85 ± 0.99 d | 59.26 ± 1.26 c | 74.26 ± 0.93 b | 84.13 ± 1.39 a |
| Ile | 10.62 ± 0.17 f | 36.65 ± 0.63 e | 154.35 ± 4.67 d | 216.44 ± 4.41 c | 277.72 ± 4.31 b | 318.41 ± 6.75 a |
| Leu | 4.44 ± 1.03 e | 3.3 ± 0.06 e | 27.42 ± 0.23 d | 38.2 ± 1.55 c | 48.74 ± 0.42 b | 57.23 ± 1.05 a |
| TBAA | 135.04 | 353.05 | 1087.85 | 1498.88 | 1914.80 | 2164.09 |
| Astringent | ||||||
| Tyr | 16.46 ± 0.45 f | 38.79 ± 0.17 e | 89.47 ± 2.04 d | 113.22 ± 1.72 c | 130.5 ± 1.9 b | 140.08 ± 2.7 a |
| TAAA | 16.46 | 38.79 | 89.47 | 113.22 | 130.5 | 140.08 |
| TAA | 254.77 | 625.42 | 1798.43 | 2636.19 | 3289.98 | 3669.36 |
| Organic Acid (mg/L) | D0 | D6 | D12 | D18 | D24 | D30 |
|---|---|---|---|---|---|---|
| Oxalic acid | 54.3 ± 0.1 c | 37.1 ± 0.8 d | 59.4 ± 2.6 b | 61.9 ± 0.9 a | 57.4 ± 0.5 b | 53.0 ± 0.1 c |
| Tartaric acid | 785.0 ± 10.5 d | 784.7 ± 9.1 d | 1102.0 ± 4.5 a | 1067.5 ± 18.5 a,b | 1014.4 ± 17.4 b | 953.9 ± 14.1 c |
| Lactic acid | 27.1 ± 0.6 f | 4027.6 ± 6.5 e | 6486.9 ± 12.3 d | 7253.0 ± 11.7 c | 7654.2 ± 79.0 b | 8073.9 ± 46.9 a |
| Citric acid | 106.7 ± 2.2 d | 582.2 ± 17.0 c,d | 650.0 ± 11.5 c,d | 1086.0 ± 22.5b c | 1559.2 ± 23.2 a,b | 1877.4 ± 38.0 a |
| Succinic acid | 58.9 ± 0.6 f | 656.4 ± 18.6 e | 1056.3 ± 3.3 d | 1347.7 ± 6.7 c | 1808.5 ± 63.4 b | 2422.4 ± 13.0 a |
| Malic acid | 128.5 ± 2.8 e | 104.0 ± 5.4 c,d | 196.3 ± 9.7 d | 560.9 ± 13.7 c | 1123.6 ± 35.5 b | 1223.2 ± 38.4 a |
| Total | 1160.5 | 6192.0 | 9550.9 | 11,377.0 | 13,217.3 | 14,603.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, A.; Yang, X.; Guo, Q.; Li, B.; Zheng, Y.; Shi, Y.; Zhu, L. Microbial Communities and Flavor Compounds during the Fermentation of Traditional Hong Qu Glutinous Rice Wine. Foods 2022, 11, 1097. https://doi.org/10.3390/foods11081097
Liu A, Yang X, Guo Q, Li B, Zheng Y, Shi Y, Zhu L. Microbial Communities and Flavor Compounds during the Fermentation of Traditional Hong Qu Glutinous Rice Wine. Foods. 2022; 11(8):1097. https://doi.org/10.3390/foods11081097
Chicago/Turabian StyleLiu, Anqi, Xu Yang, Quanyou Guo, Baoguo Li, Yao Zheng, Yuzhuo Shi, and Lin Zhu. 2022. "Microbial Communities and Flavor Compounds during the Fermentation of Traditional Hong Qu Glutinous Rice Wine" Foods 11, no. 8: 1097. https://doi.org/10.3390/foods11081097
APA StyleLiu, A., Yang, X., Guo, Q., Li, B., Zheng, Y., Shi, Y., & Zhu, L. (2022). Microbial Communities and Flavor Compounds during the Fermentation of Traditional Hong Qu Glutinous Rice Wine. Foods, 11(8), 1097. https://doi.org/10.3390/foods11081097