
Rennet-Induced Casein Micelle Aggregation Models: A Review




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
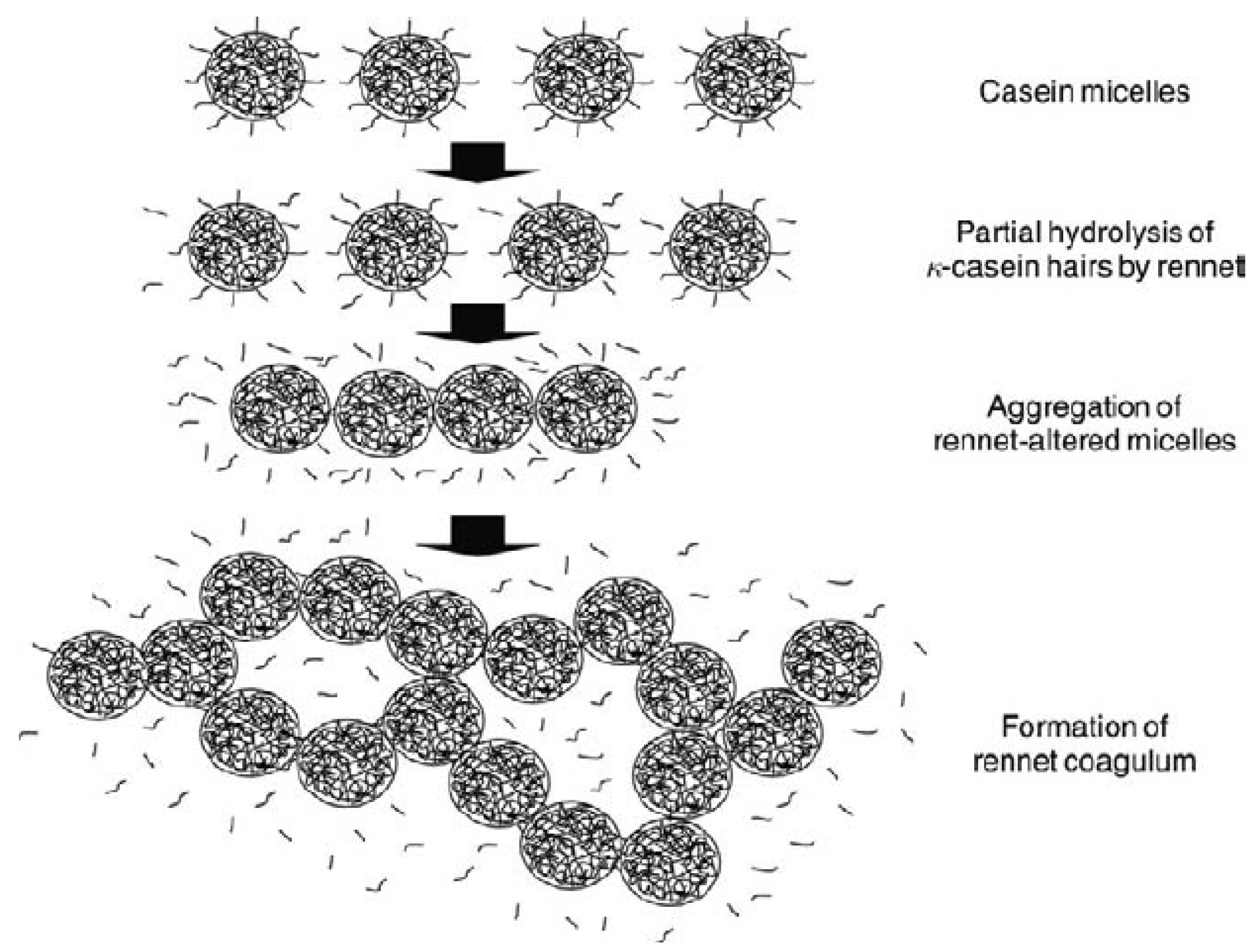
2. Micellar Aggregation
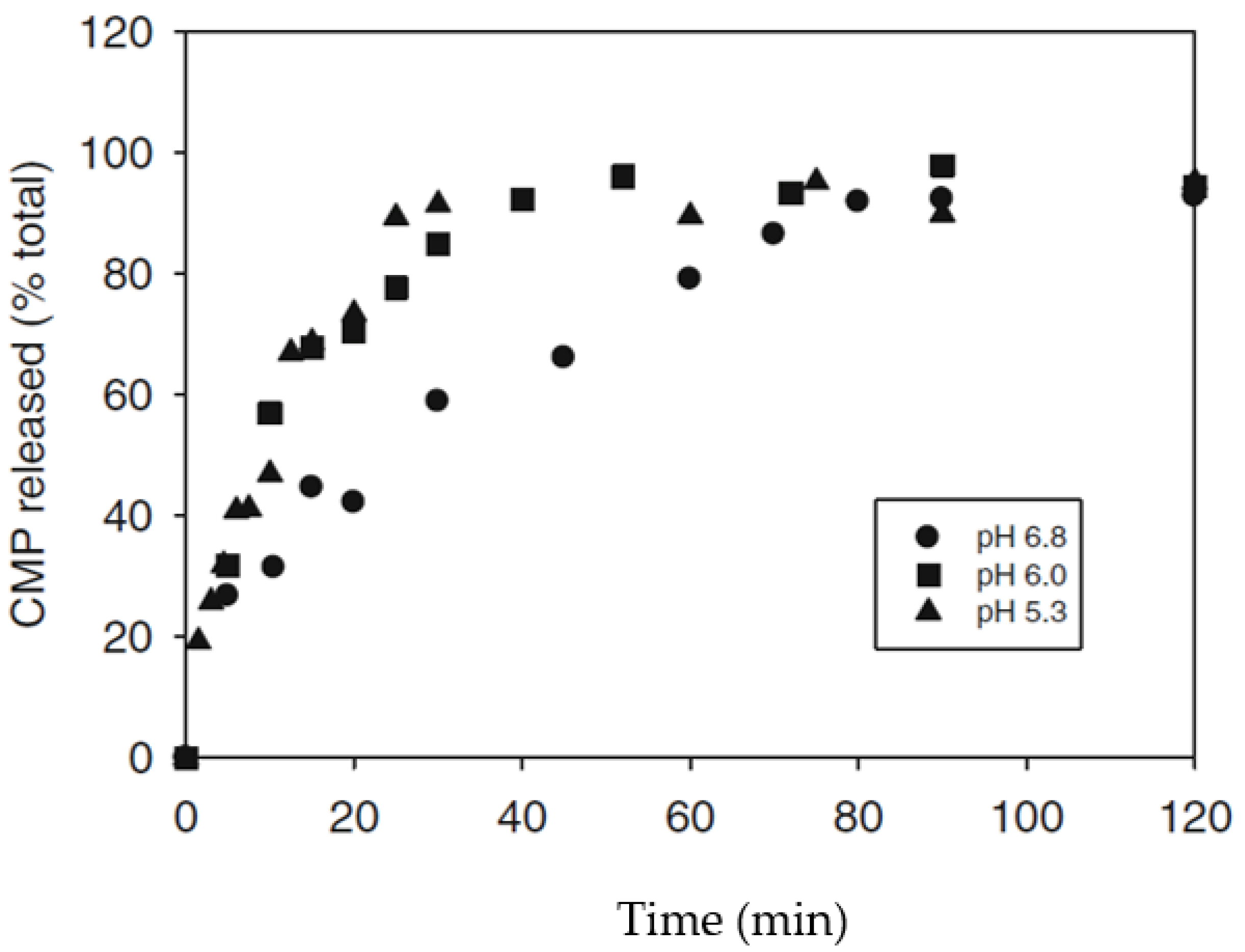
3. Aggregation Models of Hydrolyzed Casein Micelles
3.1. Holter Model
3.2. Payens Model
3.3. Step Function Model
3.4. The Energy Barrier Model
3.5. The Functionality Model
3.6. Adhesive Hard-Sphere Model
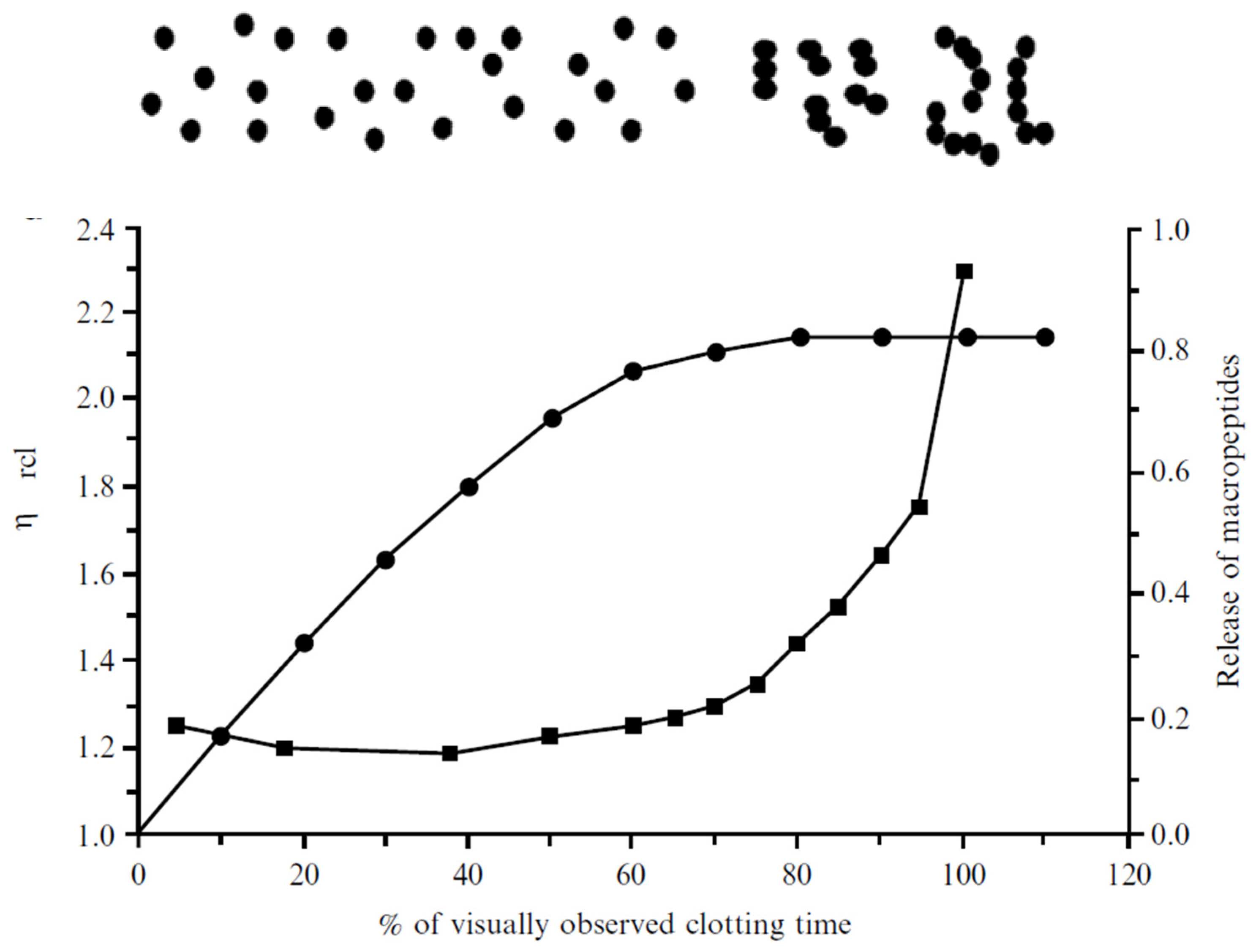
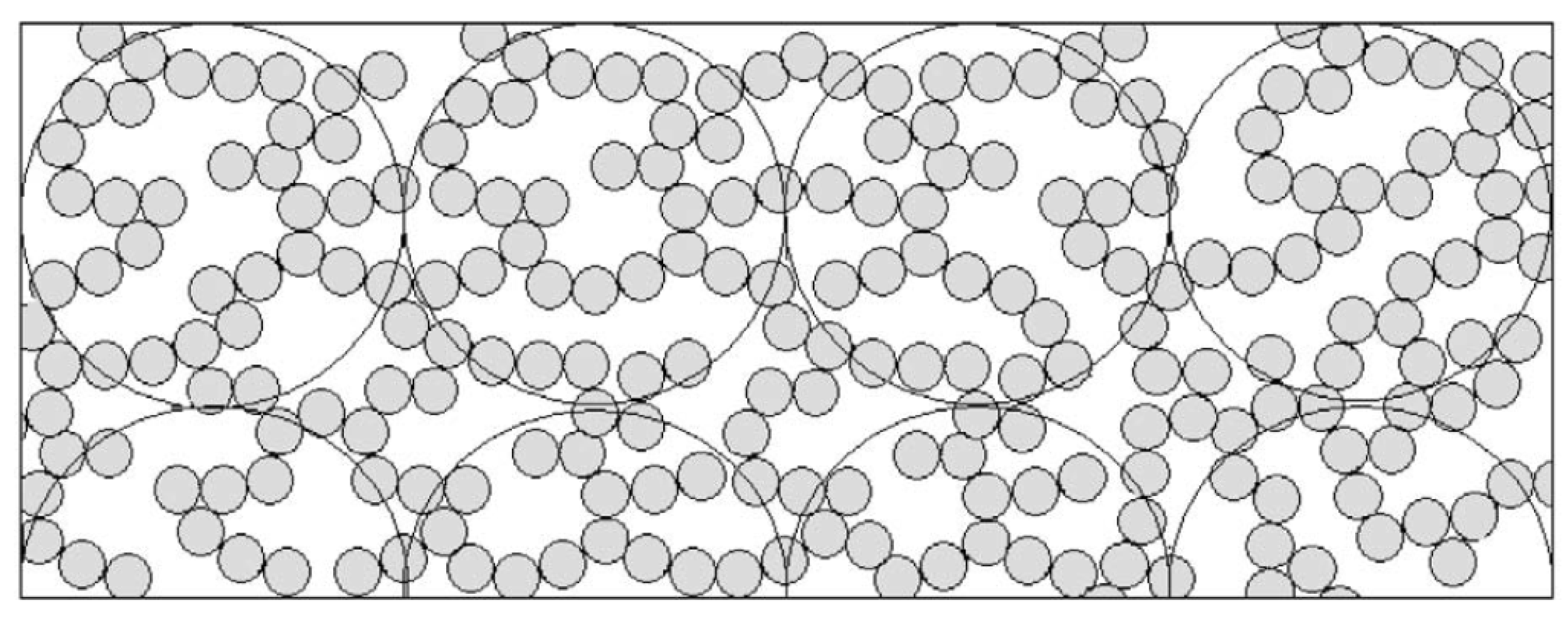
3.7. Fractal Aggregation Model
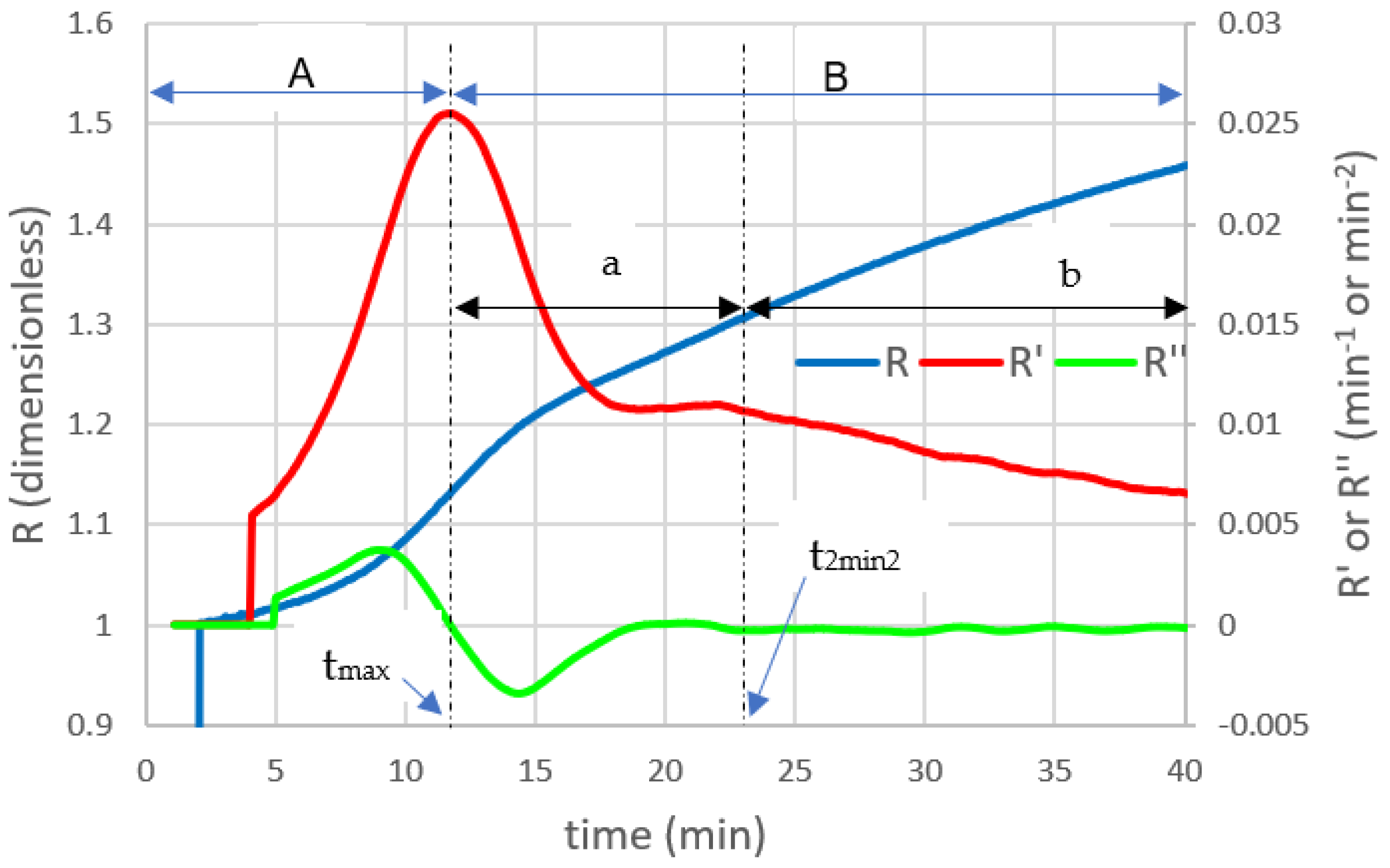
3.8. Light Scattering Model
4. Summary
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gaygadzhiev, Z.; Massel, V.; Alexander, M.; Corredig, M. Addition of Sodium Caseinate to Skim Milk Inhibits Rennet-Induced Aggregation of Casein Micelles. Food Hydrocoll. 2012, 26, 405–411. [Google Scholar] [CrossRef]
- Fox, P.F.; McSweeney, P.L.H. Rennets: Their role in milk coagulation and cheese ripening. In Microbiology and Biochemistry of Cheese and Fermented Milk, 2nd ed.; Law, B.A., Ed.; Chapman & Hall: London, UK, 1997; pp. 1–49. [Google Scholar]
- Hyslop, D. Enzymatic coagulation of milk. In Advanced Dairy Chemistry-L Proteins; Fox, P., McSweeney, P., Eds.; Kluwer Academic: New York, NY, USA; Plenum Publishers: New York, NY, USA, 2003; pp. 839–878. [Google Scholar]
- Horne, D.S.; Banks, J. Rennet-induced Coagulation of Milk. In Cheese Chemistry, Physics and Microbiology; Fox, P., McSweeney, P., Cogan, T., Guinee, T., Eds.; Elsevier Ltd.: London, UK, 2004; pp. 47–70. [Google Scholar]
- Lucey, J. Rennet-Induced Coagulation of Milk. In Encyclopedia of Dairy Sciences; Fuquay, J., Fox, P., McSweeney, P., Eds.; Elsevier: Amsterdam, The Netherlands, 2011; pp. 579–584. [Google Scholar]
- Chen, C.-C.; Chen, L.-Y.; Li, W.-T.; Chang, K.-L.; Kuo, M.-I.; Chen, C.-J.; Hsieh, J.-F. Influence of Chymosin on Physicochemical and Hydrolysis Characteristics of Casein Micelles and Individual Caseins. Nanomaterials 2021, 11, 2594. [Google Scholar] [CrossRef] [PubMed]
- Castillo, M. Cutting time prediction methods in cheese making. In Encyclopedia of Agricultural, Food, and Biological Engineering; Taylor & Francis Group: Boca Raton, FL, USA, 2006; pp. 1–7. [Google Scholar]
- Fox, P.F.; Guinee, T.P.; Cogan, T.M.; McSweeney, P.L.H. Fundamentals of Cheese Science, 2nd ed.; Springer: Boston, MA, USA, 2017; pp. 184–229. [Google Scholar]
- Payne, F.A.; Castillo, M. Light Backscatter Sensor Applications in Milk Coagulation. In Encyclopedia of Agricultural, Food, and Biological Engineering; Heldman, D., Moraru, C., Eds.; Taylor & Francis: New York, NY, USA, 2007; pp. 1530–1534. [Google Scholar]
- Arango, O.; Trujillo, A.J.; Castillo, M. Influence of Fat Replacement by Inulin on Rheological Properties, Kinetics of Rennet Milk Coagulation, and Syneresis of Milk Gels. J. Dairy Sci. 2013, 96, 1984–1996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arango, O.; Trujillo, A.J.; Castillo, M. Predicting Coagulation and Syneresis Parameters of Milk Gels When Inulin Is Added as Fat Substitute Using Infrared Light Backscatter. J. Food Eng. 2015, 157, 63–69. [Google Scholar] [CrossRef]
- Salvador, D.; Arango, O.; Castillo, M. In-Line Estimation of the Elastic Module of Milk Gels with Variation of Temperature Protein Concentration. Int. J. Food Sci. Technol. 2019, 54, 354–360. [Google Scholar] [CrossRef]
- Harboe, M.; Broe, M.; Qvist, K. The Production, Action and Application of Rennet and Coagulants. In Technology of Cheesemaking; Law, B., Tamine, A., Eds.; Blackwell Publishing Ltd.: Oxford, UK, 2010; pp. 98–129. [Google Scholar]
- Horne, D.; Lucey, J. Rennet-Induced Coagulation of Milk. In Cheese: Chemistry, Physics and Microbiology; McSweeney, P., Fox, P., Cotter, P., Everett, D., Eds.; Elsevier: Boston, MA, USA, 2017; pp. 115–143. [Google Scholar]
- Sinaga, H.; Bansal, N.; Bhandari, B. Partial Renneting of Pasteurised Bovine Milk: Casein Micelle Size, Heat and Storage Stability. Food Res. Int. 2016, 84, 52–60. [Google Scholar] [CrossRef] [Green Version]
- Koutina, G.; Knudsen, J.C.; Andersen, U.; Skibsted, L.H. Influence of Colloidal Calcium Phosphate Level on the Microstructure and Rheological Properties of Rennet-Induced Skim Milk Gels. LWT—Food Sci. Technol. 2015, 63, 654–659. [Google Scholar] [CrossRef]
- Dalgleish, D.G.; Holt, C. A Geometrical Model to Describe the Initial Aggregation of Partly Renneted Casein Micelles. J. Colloid Interface Sci. 1988, 123, 80–84. [Google Scholar] [CrossRef]
- Hill, R.D. The Effect of the Modification of Arginine Side Chains in Casein on the Coagulation of Rennin-Altered Casein. J. Dairy Res. 1970, 37, 187–192. [Google Scholar] [CrossRef]
- Peri, C.; Pagliarini, E.; Iametti, S.; Bonomi, F. A Study of Surface Hydrophobicity of Milk Proteins during Enzymic Coagulation and Curd Hardening. J. Dairy Res. 1990, 57, 101–108. [Google Scholar] [CrossRef]
- Iametti, S.; Giangiacomo, R.; Messina, G.; Bonomi, F. Influence of Processing on the Molecular Modifications of Milk Proteins in the Course of Enzymic Coagulation. J. Dairy Res. 1993, 60, 151–159. [Google Scholar] [CrossRef]
- Bauland, J.; Famelart, M.H.; Bouhallab, S.; Jeantet, R.; Roustel, S.; Faiveley, M.; Croguennec, T. Addition of Calcium and Magnesium Chlorides as Simple Means of Varying Bound and Precipitated Minerals in Casein Micelle: Effect on Enzymatic Coagulation. J. Dairy Sci. 2020, 103, 9923–9935. [Google Scholar] [CrossRef] [PubMed]
- Walstra, P.; Wouters, J.; Geurts, T. Dairy Science and Technology, 2nd ed.; Taylor & Francis Group: Boca Raton, FL, USA, 2006; pp. 34–37. [Google Scholar]
- Corredig, M.; Salvatore, E. Enzymatic Coagulation of Milk. In Advanced Dairy Chemistry: Volume 1B: Proteins: Applied Aspects; McSweeney, P.L.H., O’Mahony, J.A., Eds.; Springer: New York, NY, USA, 2016; pp. 287–307. [Google Scholar]
- Lauzin, A.; Bérubé, A.; Britten, M.; Pouliot, Y. Effect of PH Adjustment on the Composition and Rennet-Gelation Properties of Milk Concentrates Made from Ultrafiltration and Reverse Osmosis. J. Dairy Sci. 2019, 102, 3939–3946. [Google Scholar] [CrossRef] [PubMed]
- Berridge, N.J. An Improved Method of Observing the Clotting of Milk Containing Rennin. J. Dairy Res. 1952, 19, 328–329. [Google Scholar] [CrossRef]
- Lazouskaya, M.; Stulova, I.; Sõrmus, A.; Scheler, O.; Tiisma, K.; Vinter, T.; Loov, R.; Tamm, M. Front-Face Fluorimeter for the Determination of Cutting Time of Cheese Curd. Foods 2021, 10, 576. [Google Scholar] [CrossRef] [PubMed]
- Castillo, M.; Payne, F.A.; Hicks, C.L.; Lopez, M.B. Predicting Cutting and Clotting Time of Coagulating Goat’s Milk Using Difuse Reflectance: Efect of PH, Temperature and Enzyme Concentration. Int. Dairy J. 2000, 10, 12. [Google Scholar] [CrossRef]
- Tabayehnejad, N.; Castillo, M.; Payne, F.A. Comparison of Total Milk-Clotting Activity Measurement Precision Using the Berridge Clotting Time Method and a Proposed Optical Method. J. Food Eng. 2012, 108, 549–556. [Google Scholar] [CrossRef]
- Castillo, M.Z.; Payne, F.A.; Hicks, C.L.; Laencina, J.S.; López, M.-B.M. Modelling Casein Aggregation and Curd Firming in Goats’ Milk from Backscatter of Infrared Light. J. Dairy Res. 2003, 70, 335–348. [Google Scholar] [CrossRef] [Green Version]
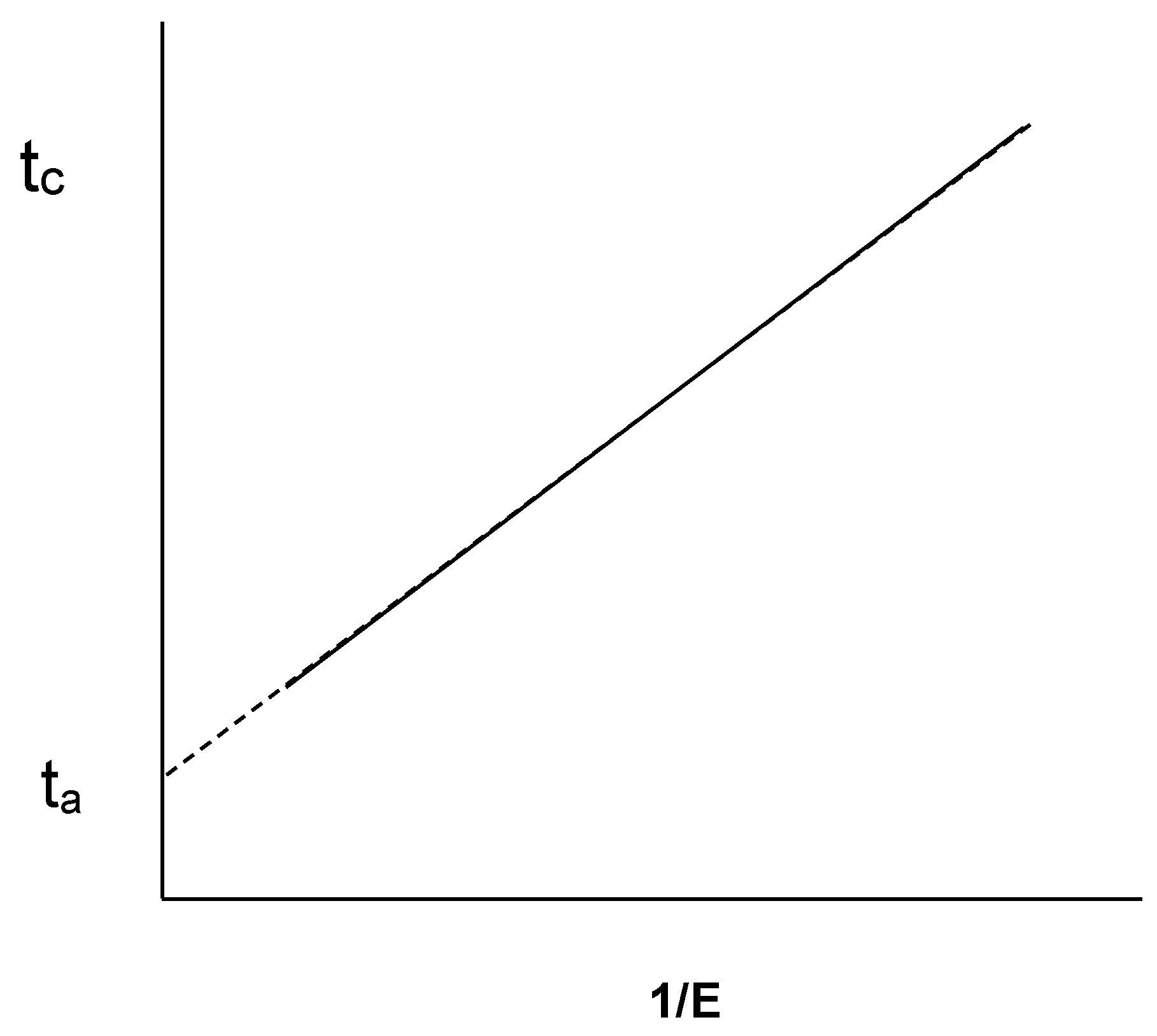
- McMahon, D.J.; Brown, R.J. Milk Coagulation Time: Linear Relationship with Inverse of Rennet Activity. J. Dairy Sci. 1983, 66, 341–344. [Google Scholar] [CrossRef]
- Holter, H. Über die Labwirkung. Biochem. Zeitschr. 1932, 255, 160–168. [Google Scholar]
- Foltmann, B. On the enzymatic and the coagulation stages of the renneting process. In Proceedings of the 15th International Dairy Congress, London, UK, 29 June–3 July 1959. [Google Scholar]
- Foltmann, B. The biochemistry of prorennin (prochymosin) and rennin (chymosin). In Milk Proteins: Chemistry and Molecular Biology II; McKenzie, H.A., Ed.; Academic Press: New York, NY, USA, 1971; pp. 217–254. [Google Scholar]
- Dalgleish, D.G. A New Calculation of the Kinetics of the Renneting Reaction. J. Dairy Res. 1988, 55, 521–528. [Google Scholar] [CrossRef]
- Payens, T. On the enzyme—Triggered clotting of casein; A preliminary account. Neth. Milk Dairy J. 1976, 30, 55–59. [Google Scholar]
- Payens, T.A.J.; Wiersma, A.K.; Brinkhuis, J. On enzymatic clotting processes I. kinetics of enzyme-triggered coagulation reactions. Biophys. Chem. 1977, 6, 253–261. [Google Scholar] [CrossRef]
- Payens, T.A.J.; Wiersma, A.K. On enzymic clotting processes v. Rate equations for the case of armtrary rate of production of the clotting specxes. Biophys. Chem. 1980, 11, 137–146. [Google Scholar] [CrossRef]
- McMahon, D.J.; Brown, R.J. Enzymic Coagulation of Casein Micelles: A Review. J. Dairy Sci. 1984, 67, 919–929. [Google Scholar] [CrossRef]
- Dalgleish, D. The Enzymatic Coagulation of Milk. In Cheese: Chemistry, Physics and Microbiology; Fox, P.F., Ed.; Springer: Boston, FL, USA, 1993; pp. 69–100. [Google Scholar]
- Dalgleish, D.G.; Brinkhuis, J.; Payens, T.A.J. The Coagulation of Differently Sized Casein Micelles by Rennet. Eur. J. Biochem. 1981, 119, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Dalgleish, D.G. Coagulation of Renneted Bovine Casein Micelles: Dependence on Temperature, Calcium Ion Concentration and Ionic Strength. J. Dairy Res. 1983, 50, 331–340. [Google Scholar] [CrossRef]
- Brinkhuis, J.; Payens, T.A. The influence of temperature on the flocculation rate of renneted casein micelles. Biophys. Chem. 1984, 19, 75–81. [Google Scholar] [CrossRef]
- Dalgleish, D.G. Effect of Milk Concentration on the Rennet Coagulation Time. J. Dairy Res. 1980, 47, 231–235. [Google Scholar] [CrossRef]
- Dalgleish, D.G. A Mechanism for the Chymosin-Induced Flocculation of Casein Micelles. Biophys. Chem. 1980, 11, 147–155. [Google Scholar] [CrossRef]
- Darling, D.F.; Van Hooydonk, A.C.M. Derivation of a Mathematical Model for the Mechanism of Casein Micelle Coagulation by Rennet. J. Dairy Res. 1981, 48, 189–200. [Google Scholar] [CrossRef]
- Hyslop, D.B. Enzyme-Induced Coagulation of Casein Micelles: A Number of Different Kinetic Models. J. Dairy Res. 1993, 60, 517–533. [Google Scholar] [CrossRef]
- Hyslop, D.B.; Qvist, K.B. Application of Numerical Analysis to a Number of Models for Chymosin-Induced Coagulation of Casein Micelles. J. Dairy Res. 1996, 63, 223–232. [Google Scholar] [CrossRef]
- Stockmayer, W.H. Theory of Molecular Size Distribution and Gel Formation in Branched-Chain Polymers. J. Chem. Phys. 1943, 11, 45–55. [Google Scholar] [CrossRef]
- De Kruif, C.G. Casein Micelles: Diffusivity as a Function of Renneting Time. Langmuir 1992, 8, 2932–2937. [Google Scholar] [CrossRef]
- De Kruif, C.G. Casein Micelle Interactions. Int. Dairy J. 1999, 9, 183–188. [Google Scholar] [CrossRef]
- López, M.B.; Jordán, M.J.; Granados, M.V.; Fernández, J.C.; Castillo, M.; Laencina, J. Viscosity Changes during Rennet Coagulation of Murciano-Granadina Goat Milk. Int. J. Dairy Technol. 1999, 52, 102–106. [Google Scholar] [CrossRef]
- Walstra, P.; Bloomfield, V.A.; Wei, G.J.; Jenness, R. Effect of Chymosin Action on the Hydrodynamic Diameter of Casein Micelle. Biochim. Biophys. Acta 1981, 669, 258–259. [Google Scholar] [CrossRef]
- Horne, D.S.; Davidson, C.M. Direct Observation of Decrease in Size of Casein Micelles during the Initial Stages of Renneting of Skim Milk. Int. Dairy J. 1993, 3, 61–71. [Google Scholar] [CrossRef]
- Horne, D.S. Casein Micelles as Hard Spheres: Limitations of the Model in Acidified Gel Formation. Colloids Surf. A Physicochem. Eng. Asp. 2003, 213, 255–263. [Google Scholar] [CrossRef]
- Bremer, L.G.B.; van Vliet, T.; Walstra, P. Theoretical and Experimental Study of the Fractal Nature of the Structure of Casein Gels. J. Chem. Soc. Faraday Trans. 1 Phys. Chem. Condens. Phases 1989, 85, 3359. [Google Scholar] [CrossRef]
- Bremer, L.G.B.; Bijsterbosch, B.H.; Schrijvers, R.; van Vliet, T.; Walstra, P. On the Fractal Nature of the Structure of Acid Casein Gels. Colloids Surf. 1990, 51, 159–170. [Google Scholar] [CrossRef]
- Bremer, L.G.B.; Bijsterbosch, B.H.; Walstra, P.; van Vliet, T. Formation, Properties and Fractal Structure of Particle Gels. Adv. Colloid Interface Sci. 1993, 46, 117–128. [Google Scholar] [CrossRef]
- Horne, D.S. Determination of the Fractal Dimension Using Turbidimetric Techniques. Application to Aggregating Protein Systems. Faraday Discuss. Chem. Soc. 1987, 83, 259. [Google Scholar] [CrossRef]
- Worning, P.; Bauer, R.; Øgendal, L.; Lomholt, S. A Novel Approach to Turbidimetry of Dense Systems: An Investigation of the Enzymatic Gelation of Casein Micelles. J. Colloid Interface Sci. 1998, 203, 265–277. [Google Scholar] [CrossRef]
- Lehner, D.; Worning, P.; Fritz, G.; Øgendal, L.; Bauer, R.; Glatter, O. Characterization of Enzymatically Induced Aggregation of Casein Micelles in Natural Concentration Byin SituStatic Light Scattering and Ultra Low Shear Viscosimetry. J. Colloid Interface Sci. 1999, 213, 445–456. [Google Scholar] [CrossRef]
- Vétier, N.; Banon, S.; Ramet, J.-P.; Hardy, J. Hydratation des micelles de caséine et structure fractale des agrégats et des gels de lait. Lait 2000, 80, 237–246. [Google Scholar] [CrossRef]
- Vétier, N.; Banon, S.; Chardot, V.; Hardy, J. Effect of Temperature and Aggregation Rate on the Fractal Dimension of Renneted Casein Aggregates. J. Dairy Sci. 2003, 86, 2504–2507. [Google Scholar] [CrossRef]
- Walstra, P. Physical Chemistry of Foods; Marcel Dekker: New York, NY, USA, 2003; pp. 425–486. [Google Scholar]
- Horne, D.S. Formation and Structure of Acidified Milk Gels. Int. Dairy J. 1999, 8, 261–268. [Google Scholar] [CrossRef]
- Mellema, M.; Heesakkers, J.W.M.; van Opheusden, J.H.J.; van Vliet, T. Structure and Scaling Behavior of Aging Rennet-Induced Casein Gels Examined by Confocal Microscopy and Permeametry. Langmuir 2000, 16, 6847–6854. [Google Scholar] [CrossRef]
- Mellema, M.; Walstra, P.; van Opheusden, J.H.J.; van Vliet, T. Effects of Structural Rearrangements on the Rheology of Rennet-Induced Casein Particle Gels. Adv. Colloid Interface Sci. 2002, 98, 25–50. [Google Scholar] [CrossRef]
- Bauer, R.; Hansen, M.; Hansen, S.; Øgendal, L.; Lomholt, S.; Qvist; Horne, D. The structure of casein aggregates during renneting studied by indirect Fourier transformation and inverse Laplace transformation of static and dynamic light scattering data, respectively. J. Chem. Phys. 1995, 103, 2725–2737. [Google Scholar] [CrossRef]
- Lomholt, S.B.; Worning, P.; Øgendal, L.; Qvist, K.B.; Hyslop, D.B.; Bauer, R. Kinetics of the Renneting Reaction Followed by Measurement of Turbidity as a Function of Wavelength. J. Dairy Res. 1998, 65, 545–554. [Google Scholar] [CrossRef]
- Smykov, I.T. Kinetic of milk gelation. Part I Coagulation mechanism. In Rheology: Principles, Applications and Enviromental Impacts, 1st ed.; Karpushkin, E., Ed.; Nova Science Piblisher, Inc.: New York, NY, USA, 2015; pp. 65–82. [Google Scholar]
- Hass, R.; Munzke, D.; Vargas Ruiz, S.; Tippmann, J.; Reich, O. Optical Monitoring of Chemical Processes in Turbid Biogenic Liquid Dispersions by Photon Density Wave Spectroscopy. Anal. Bioanal. Chem. 2015, 407, 2791–2802. [Google Scholar] [CrossRef] [PubMed]
- Shayeh, J.S.; Sefidbakht, Y.; Siadat, S.O.R.; Niknam, K. Continuous Fast Fourier Transforms Cyclic Voltammetry as a New Approach for Investigation of Skim Milk K-Casein Proteolysis, a Comparative Study. Int. J. Biol. Macromol. 2017, 103, 972–977. [Google Scholar] [CrossRef] [PubMed]
- Panikuttira, B.; Payne, F.A.; O’Shea, N.; Tobin, J.T.; O’Callaghan, D.J.; O’Donnell, C.P. Investigation of an in-line prototype fluorescence and infrared backscatter sensor to monitor rennet-induced coagulation of skim milk at different protein concentrations. Int. J. Food Sci. Technol. 2020, 55, 175–182. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salvador, D.; Acosta, Y.; Zamora, A.; Castillo, M. Rennet-Induced Casein Micelle Aggregation Models: A Review. Foods 2022, 11, 1243. https://doi.org/10.3390/foods11091243
Salvador D, Acosta Y, Zamora A, Castillo M. Rennet-Induced Casein Micelle Aggregation Models: A Review. Foods. 2022; 11(9):1243. https://doi.org/10.3390/foods11091243
Chicago/Turabian StyleSalvador, Daniel, Yoseli Acosta, Anna Zamora, and Manuel Castillo. 2022. "Rennet-Induced Casein Micelle Aggregation Models: A Review" Foods 11, no. 9: 1243. https://doi.org/10.3390/foods11091243
APA StyleSalvador, D., Acosta, Y., Zamora, A., & Castillo, M. (2022). Rennet-Induced Casein Micelle Aggregation Models: A Review. Foods, 11(9), 1243. https://doi.org/10.3390/foods11091243