
Enzymatic Hydrolysis of Pulse Proteins as a Tool to Improve Techno-Functional Properties



Abstract
:1. Introduction
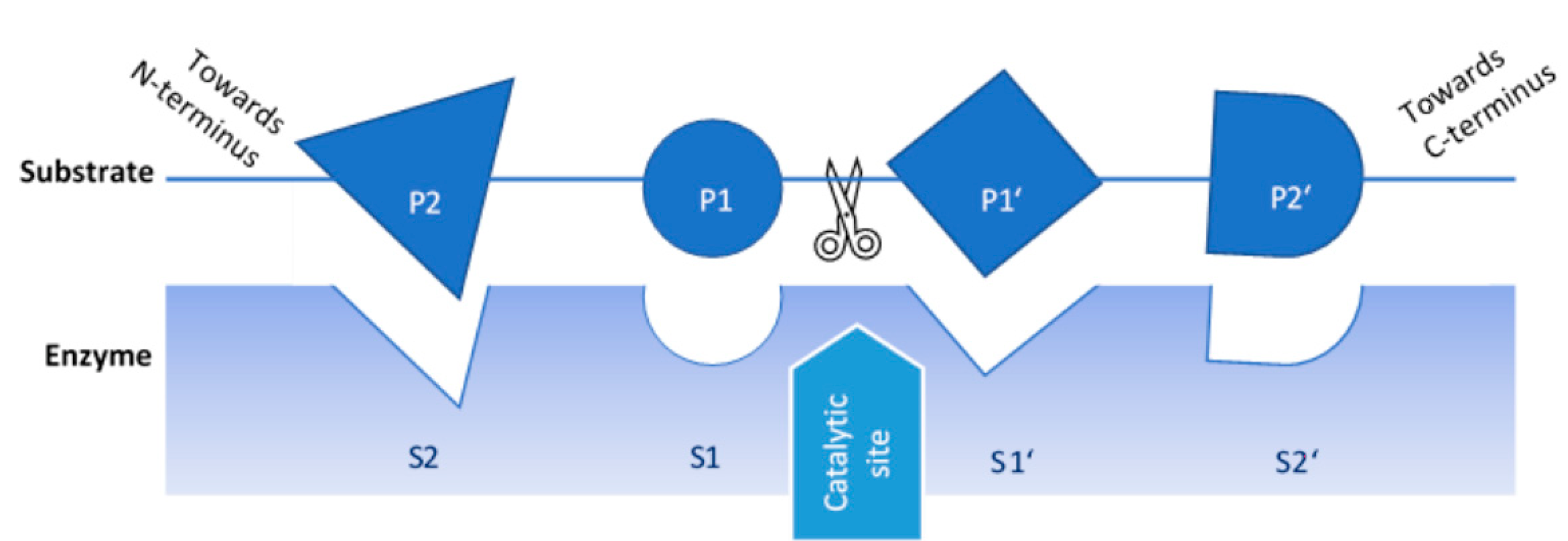
2. Proteases
3. Production of Protein Hydrolysates and Assessment of the Extent of Hydrolysis
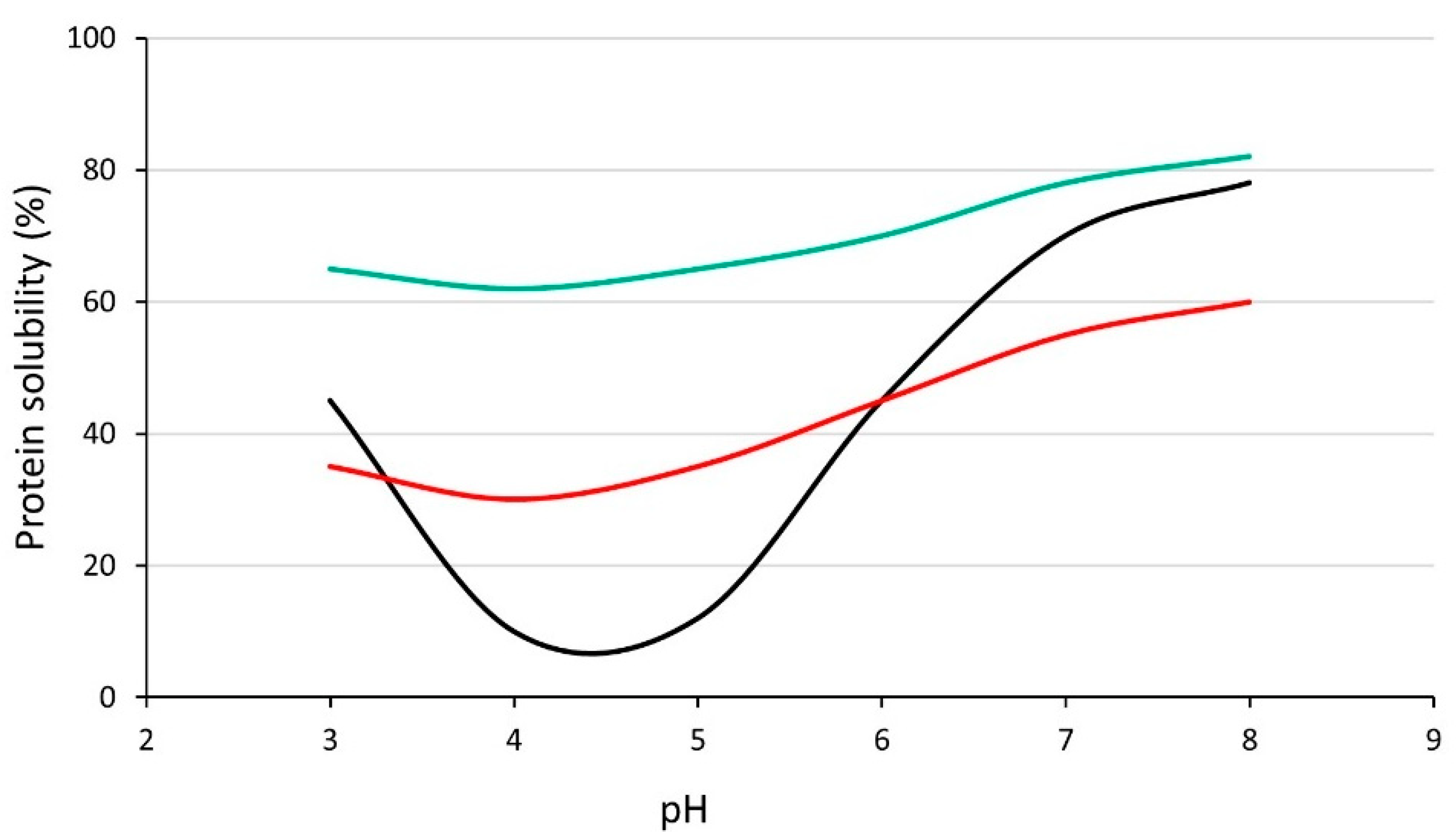
4. Solubility

{kind=link}
{kind=link}
| Reference | Protein Source | Protease | Effect on Solubility |
|---|---|---|---|
| Barać et al. [66] | Pea protein isolate | Chymosin | Increased at pH 3; increased/no difference at pH 5 depending on HT; decreased at pH 7; increased at pH 8 |
| Barac et al. [50] * | Pea protein isolate (L1) | Papain | Increased at pH 3 and 5; increased/decreased at pH 7 depending on HT; increased at pH 8 |
| S. griseus protease | Increased at pH 3 and 5; decreased at pH 7 and 8 | ||
| Pea protein isolate (Maja) | Papain | Increased at pH 3 and 5; increased/decreased at pH 7 depending on HT; decreased at pH 8 | |
| S. griseus protease | Increased at pH 3, 5 and 7; increased/decreased at pH 8 depending on HT | ||
| Betancur-Ancona et al. [63] | P. lunatus protein isolate | Alcalase | Increased at pH 2, 4, 6, 8 and 10 |
| Flavourzyme | Increased/no difference at pH 2 depending on HT; increased at pH 4 and 6; increased /no difference at pH 8 and 10 depending on HT | ||
| Eckert et al. [39] | Faba bean protein isolate | Pepsin | Increased at pH 5 and 7 |
| Trypsin | Increased at pH 5 and 7 | ||
| Flavourzyme | Increased at pH 5 and 7 | ||
| Neutrase | Increased at pH 5 and 7 | ||
| García Arteaga et al. [35] | Pea protein isolate | Alcalase | Increased at pH 4.5; increased/no difference at pH 7 depending on HT |
| Papain | Increased at pH 4.5; no difference at pH 7 | ||
| Esperase | Increased at pH 4.5 and pH 7 | ||
| Bromelain | Increased at pH 4.5; decreased/no difference at pH 7 depending on HT | ||
| Trypsin | Increased at pH 4.5; increased/no difference at pH 7 depending on HT | ||
| Chymotrypsin | Increased/no difference at pH 4.5 depending on HT; decreased at pH 7 | ||
| Klost and Drusch [44] | Pea protein concentrate | Trypsin | Decreased/no difference at pH 3 depending on DH; increased at pH 4, 5, and 6; decreased at pH 7 |
| Konieczny et al. [64] | Pea protein-enriched flour | Trypsin | Decreased at pH 4, 7 and 10 |
| Savinase | Decreased at pH 4, 7 and 10 | ||
| Papain | Decreased at pH 4, 7 and 10 | ||
| Pepsin | Decreased at pH 4, and 7; decreased/no difference at pH 10 depending on DH | ||
| Mokni Ghribi et al. [46] | Chickpea protein isolate | Alcalase | Increased at pH 2, 4, 6, 8, 10 and 12 |
| Schlegel et al. [40] | Lupin protein isolate | Alcalase | Increased at pH 4, 5 and 6; no difference at pH 7, 8 and 9 |
| Papain | Increased at pH 4, 5 and 6; no difference at pH 7, 8 and 9 | ||
| Neutrase | Increased at pH 4, 5 and 6; no difference at pH 7, 8 and 9 | ||
| Protease N-01 | Increased at pH 4 and 5; no difference at pH 6, 7, 8 and 9 | ||
| Flavourzyme | Increased at pH 4 and 5; no difference at pH 6; decreased at pH 7, 8 and 9 | ||
| Protamex | Increased at pH 4, 5 and 6; no difference at pH 7, 8 and 9 | ||
| Corolase 7089 | Increased at pH 4, 5 and 6; no difference at pH 7, 8 and 9 | ||
| Pepsin | Increased at pH 4, 5 and 6; no difference at pH 7, 8 and 9 | ||
| Corolase N | Increased at pH 4, 5 and 6; no difference at pH 7, 8 and 9 | ||
| Segura-Campos et al. [36] | Cowpea protein concentrate | Alcalase | Decreased at pH 2; increased at pH 4 and 6; decreased at pH 8 and 10 |
| Flavourzyme | Increased at pH 2, 4, 6, 8 and 10 | ||
| Xu et al. [67] | Chickpea protein isolate | Alcalase | Increased at pH 2, 4, 7 and 9 |
| Flavourzyme | Increased at pH 2, 4, 7 and 9 | ||
| Yust et al. [42] | Chickpea protein isolate | Alcalase | Increased/no difference at pH < 4 depending on DH; increased at pH ~4–6; increased/decreased at pH 7 depending on DH; increased at pH 8, 9 and 10 |
| Zhang and Motta [45] * | Great Northern bean protein concentrate | Alcalase | Decreased/no difference at pH 3 depending on DH; increased at pH 4, 5 and 6; no difference at pH 7 |
| Papain | Decreased at pH 3; increased at pH 4, 5 and 6; no difference at pH 7 | ||
| Navy bean protein concentrate | Alcalase | Increased/no difference at pH 3 depending on DH; decreased/no difference at pH 4 depending on DH; increased at pH 5 and 6; no difference at pH 7 | |
| Papain | Increased at pH 3; no difference at pH 4; increased at pH 5 and 6; no difference at pH 7 |
5. Emulsifying Properties
| Reference | Protein Source | Protease | Effect on Emulsifying Properties |
|---|---|---|---|
| Ahmed et al. [52] | Lentil protein isolate | Alcalase | EAI: decreased; ESI: decreased |
| Al-Ruwaih et al. [51] | Kidney bean protein isolate | Alcalase | EAI: increased (but decreased for high pressure treated sample) ESI: decreased |
| Avramenko et al. [48] | Lentil protein isolate | Trypsin | EAI: decreased; ESI: decreased |
| Barać et al. [66] | Pea protein isolate | Chymosin | EAI: increased at pH 3; increased/no difference at pH 5 depending on HT; increased/decreased at pH 7 depending on HT; decreased at pH 8 ESI: decreased at pH 3; increased/decreased at pH 5 depending on HT; increased/no difference at pH 7 and 8 depending on HT |
| Barac et al. [50] | Pea protein isolate (L1) | Papain | EAI: increased at pH 3, 5, 7, and 8 ESI: increased at pH 3; decreased/no difference at pH 5 depending on HT; increased at pH 7 and 8 |
| S. griseus protease | EAI: increased at pH 3, 5, 7 and 8 ESI: increased at pH 3; decreased at pH 5; increased at pH 7 and 8 | ||
| Pea protein isolate (Maja) | Papain | EAI: decreased at pH 3; increased/decreased at pH 5 and 7 depending on HT; increased/no difference at pH 8 depending on HT ESI: increased/decreased at pH 3 and 5 depending on HT; decreased at pH 7 and 8 | |
| S. griseus protease | EAI: increased/decreased at pH 3, 5, 7, and 8 depending on HTESI: increased at pH 3 and 5; increased/decreased at pH 7 depending on HT; decreased at pH 8 | ||
| Betancur-Ancona et al. [63] | P. lunatus protein isolate | Alcalase | EC: decreased at pH 2, 4, 6, 8, and 10 ES: decreased at pH 2; increased at pH 4; decreased at pH 6, 8, and 10 |
| Flavourzyme | EC: increased at pH 2; no difference at pH 4; increased/no difference at pH 6 depending on HT; increased at pH 8 and 10ES: No difference at pH 2; increased at pH 4 and 6; decreased/no difference depending on HT at pH 8 and 10 | ||
| Eckert et al. [39] | Faba bean protein isolate | Pepsin | Decreased EAI and ESI |
| Trypsin | Increased/decreased EAI and ESI depending on HT | ||
| Flavourzyme | Decreased EAI; increased ESI | ||
| Neutrase | No difference in EAI, increased ESI | ||
| García Arteaga et al. [35] | Pea protein isolate | Alcalase | EC: no difference |
| Papain | EC: no difference | ||
| Esperase | EC: increased/no difference depending on HT | ||
| Bromelain | EC: no difference | ||
| Trypsin | EC: increased | ||
| Chymotrypsin | EC: increased | ||
| Konieczny et al. [64] | Pea protein-enriched flour | Trypsin | EAI: increased at pH 4; increased/decreased at pH 7 depending on DH; increased at pH 10 ESI: decreased at pH 4, 7, and 10 |
| Savinase | EAI: increased/decreased at pH 4 depending on DH; decreased at pH 7; increased at pH 10 ESI: decreased at pH 4, 7, and 10 | ||
| Papain | EAI: decreased at pH 4, 7, and 10 ESI: increased at pH 4; decreased at pH 7 and 10 | ||
| Pepsin | EAI: decreased at pH 4, 7, and 10 ESI: decreased at pH 4, 7, and 10 | ||
| Mokni Ghribi et al. [46] | Chickpea protein isolate | Alcalase | EAI: increased/decreased depending on DH ESI: decreased/no difference depending on DH |
| Schlegel et al. [40] | Lupin protein isolate | Alcalase | EC: decreased |
| Papain | EC: decreased | ||
| Neutrase | EC: decreased | ||
| Protease N-01 | EC: no difference | ||
| Flavourzyme | EC: decreased | ||
| Protamex | EC: decreased | ||
| Corolase 7089 | EC: no difference | ||
| Pepsin | EC: no difference | ||
| Corolase N | EC: no difference | ||
| Wani et al. [76] | Kidney bean protein isolate (French Yellow) | Papain | EAI: increased/decreased at pH 3 depending on HT; decreased at pH 5; increased at pH 7 ESI: increased/no difference at pH 3 and 5 depending on HT; no difference at pH 7 |
| Kidney bean protein isolate (Contender) | Papain | EAI: increased at pH 3, 5 and 7 ESI: decreased/no difference at pH 3 depending on HT; increased/no difference 5 depending on HT; decreased at pH 7 | |
| Kidney bean protein isolate (Master Bean) | Papain | EAI: increased at pH 3, 5 and 7 ESI: increased/no difference at pH 3 depending on HT; decreased at pH 5 and 7 | |
| Kidney bean protein isolate (Local Red) | Papain | EAI: increased at pH 3, 5 and 7 ESI: no difference at pH 3; increased/no difference at pH 5 and 7 depending on HT | |
| Wani et al. [77] | Black gram protein isolate (Mash 1-1) | Papain | EAI: increased at pH 3, 5 and 7 ESI: increased/no difference at pH 3 depending on HT; increased at pH 5; decreased/no difference at pH 7 depending on HT |
| Black gram protein isolate (PU-19) | Papain | EAI: increased/decreased at pH 3 and 5 depending on HT; increased/no difference at pH 8 depending on HT ESI: increased/decreased at pH 3 and 5 depending on HT; decreased at pH 8 | |
| Black gram protein isolate (T-9) | Papain | EAI: increased/decreased at pH 3 depending on HT; increased at pH 5; increased/decreased at pH 7 depending on HT ESI: increased at pH 3 and 5; increased/no difference at pH 7 depending on HT | |
| Xu et al. [67] | Chickpea protein isolate | Alcalase | EAI: increased; ESI: increased |
| Flavourzyme | EAI: increased; ESI: increased |
6. Foaming Properties
7. Gelation and Rheological Properties
8. Sensory Considerations
9. Future Outlook
10. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aiking, H.; de Boer, J. The next protein transition. Trends Food Sci. Technol. 2020, 105, 515–522. [Google Scholar] [CrossRef]
- Akharume, F.U.; Aluko, R.E.; Adedeji, A.A. Modification of plant proteins for improved functionality: A review. Compr. Rev. Food Sci. Food Saf. 2021, 20, 198–224. [Google Scholar] [CrossRef] [PubMed]
- Poore, J.; Nemecek, T. Reducing food’s environmental impacts through producers and consumers. Science 2018, 360, 987–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arntfield, S.D.; Maskus, H.D. Peas and other legume proteins. In Handbook of Food Proteins; Woodhead Publishing: Sawston, UK, 2011; pp. 233–266. [Google Scholar] [CrossRef]
- Sozer, N.; Holopainen-Mantila, U.; Poutanen, K. Traditional and New Food Uses of Pulses. Cereal Chem. J. 2017, 94, 66–73. [Google Scholar] [CrossRef]
- Karaca, A.C.; Low, N.; Nickerson, M. Emulsifying properties of chickpea, faba bean, lentil and pea proteins produced by isoelectric precipitation and salt extraction. Food Res. Int. 2011, 44, 2742–2750. [Google Scholar] [CrossRef]
- Tamm, F.; Herbst, S.; Brodkorb, A.; Drusch, S. Functional properties of pea protein hydrolysates in emulsions and spray-dried microcapsules. Food Hydrocoll. 2016, 58, 204–214. [Google Scholar] [CrossRef]
- Park, S.J.; Kim, T.W.; Baik, B.-K. Relationship between proportion and composition of albumins, and in vitro protein digestibility of raw and cooked pea seeds (Pisum sativum L.). J. Sci. Food Agric. 2010, 90, 1719–1725. [Google Scholar] [CrossRef]
- Stone, A.K.; Nosworthy, M.G.; Chiremba, C.; House, J.D.; Nickerson, M.T. A comparative study of the functionality and protein quality of a variety of legume and cereal flours. Cereal Chem. 2019, 96, 1159–1169. [Google Scholar] [CrossRef]
- Singhal, A.; Karaca, A.C.; Tyler, R.; Nickerson, M. Pulse Proteins: From Processing to Structure-Function Relationships. In Grain Legumes; Goyal, A., Ed.; IntechOpen: London, UK, 2016. [Google Scholar] [CrossRef] [Green Version]
- Koyoro, H.; Powers, J.R. Functional properties of pea globulin fractions. Cereal Chem. 1987, 64, 97–101. [Google Scholar]
- Lam, A.C.Y.; Warkentin, T.D.; Tyler, R.T.; Nickerson, M.T. Physicochemical and Functional Properties of Protein Isolates Obtained from Several Pea Cultivars. Cereal Chem. 2017, 94, 89–97. [Google Scholar] [CrossRef]
- Hall, C.; Hillen, C.; Garden Robinson, J. Composition, Nutritional Value, and Health Benefits of Pulses. Cereal Chem. 2017, 94, 11–31. [Google Scholar] [CrossRef]
- Boye, J.; Zare, F.; Pletch, A. Pulse proteins: Processing, characterization, functional properties and applications in food and feed. Food Res. Int. 2010, 43, 414–431. [Google Scholar] [CrossRef]
- Schutyser, M.A.I.; Pelgrom, P.J.M.; van der Goot, A.J.; Boom, R.M. Dry fractionation for sustainable production of functional legume protein concentrates. Trends Food Sci. Technol. 2015, 45, 327–335. [Google Scholar] [CrossRef]
- Boye, J.I.; Aksay, S.; Roufik, S.; Ribéreau, S.; Mondor, M.; Farnworth, E.; Rajamohamed, S.H. Comparison of the functional properties of pea, chickpea and lentil protein concentrates processed using ultrafiltration and isoelectric precipitation techniques. Food Res. Int. 2010, 43, 537–546. [Google Scholar] [CrossRef]
- Vogelsang-O’Dwyer, M.; Zannini, E.; Arendt, E.K. Production of pulse protein ingredients and their application in plant-based milk alternatives. Trends Food Sci. Technol. 2021, 110, 364–374. [Google Scholar] [CrossRef]
- Daba, S.D.; Morris, C.F. Pea proteins: Variation, composition, genetics, and functional properties. Cereal Chem. 2021, 99, 8–20. [Google Scholar] [CrossRef]
- Jarpa-Parra, M. Lentil protein: A review of functional properties and food application. An overview of lentil protein functionality. Int. J. Food Sci. Technol. 2018, 53, 892–903. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Pei, R.; Heinonen, M. Faba bean protein: A promising plant-based emulsifier for improving physical and oxidative stabilities of oil-in-water emulsions. Food Chem. 2021, 369, 130879. [Google Scholar] [CrossRef]
- Gaan, K.; Dabir, S.; Ignaszwewski, E.; Manu, N.; Murray, S.; Weston, Z. State of the Industry Report—Plant-Based Meat, Eggs, and Dairy; The Good Food Institute: Washington, DC, USA, 2020. [Google Scholar]
- Kyriakopoulou, K.; Keppler, J.K.; Van Der Goot, A.J. Functionality of Ingredients and Additives in Plant-Based Meat Analogues. Foods 2021, 10, 600. [Google Scholar] [CrossRef]
- Saldanha do Carmo, C.; Knutsen, S.H.; Malizia, G.; Dessev, T.; Geny, A.; Zobel, H.; Myhrer, K.S.; Varela, P.; Sahlstrøm, S. Meat analogues from a faba bean concentrate can be generated by high moisture extrusion. Future Foods 2021, 3, 100014. [Google Scholar] [CrossRef]
- Jeske, S.; Bez, J.; Arendt, E.K.; Zannini, E. Formation, stability, and sensory characteristics of a lentil-based milk substitute as affected by homogenisation and pasteurisation. Eur. Food Res. Technol. 2019, 245, 1519–1531. [Google Scholar] [CrossRef]
- McClements, D.J.; Grossmann, L. The science of plant-based foods: Constructing next-generation meat, fish, milk, and egg analogs. Compr. Rev. Food Sci. Food Saf. 2021, 20, 4049–4100. [Google Scholar] [CrossRef] [PubMed]
- Panyam, D.; Kilara, A. Enhancing the functionality of food proteins by enzymatic modification. Trends Food Sci. Technol. 1996, 7, 120–125. [Google Scholar] [CrossRef]
- Wouters, A.G.B.; Rombouts, I.; Fierens, E.; Brijs, K.; Delcour, J.A. Relevance of the Functional Properties of Enzymatic Plant Protein Hydrolysates in Food Systems. Compr. Rev. Food Sci. Food Saf. 2016, 15, 786–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aryee, A.N.A.; Boye, J.I. Improving the Digestibility of Lentil Flours and Protein Isolate and Characterization of Their Enzymatically Prepared Hydrolysates. Int. J. Food Prop. 2016, 19, 2649–2665. [Google Scholar] [CrossRef]
- Merz, M.; Eisele, T.; Berends, P.; Appel, D.; Rabe, S.; Blank, I.; Stressler, T.; Fischer, L. Flavourzyme, an Enzyme Preparation with Industrial Relevance: Automated Nine-Step Purification and Partial Characterization of Eight Enzymes. J. Agric. Food Chem. 2015, 63, 5682–5693. [Google Scholar] [CrossRef]
- Gurumallesh, P.; Alagu, K.; Ramakrishnan, B.; Muthusamy, S. A systematic reconsideration on proteases. Int. J. Biol. Macromol. 2019, 128, 254–267. [Google Scholar] [CrossRef]
- Rawlings, N.D.; Barrett, A.J. Peptidases. In eLS; John Wiley & Sons: New York, NY, USA, 2014. [Google Scholar] [CrossRef]
- Großmann, K.K.; Merz, M.; Appel, D.; Fischer, L. A fast and novel approach to evaluate technical enzyme preparations for an efficient protein hydrolysis. Eur. Food Res. Technol. 2019, 245, 1695–1708. [Google Scholar] [CrossRef] [Green Version]
- Adler-Nissen, J. Chapter 7—Proteases. In Enzymes in Food Processing, 3rd ed.; Nagodawithana, T., Reed, G., Eds.; Academic Press: London, UK, 1993; pp. 159–203. [Google Scholar] [CrossRef]
- Gupta, R.; Beg, Q.K.; Lorenz, P. Bacterial alkaline proteases: Molecular approaches and industrial applications. Appl. Microbiol. Biotechnol. 2002, 59, 15–32. [Google Scholar] [CrossRef]
- García Arteaga, V.; Apéstegui Guardia, M.; Muranyi, I.; Eisner, P.; Schweiggert-Weisz, U. Effect of enzymatic hydrolysis on molecular weight distribution, techno-functional properties and sensory perception of pea protein isolates. Innov. Food Sci. Emerg. Technol. 2020, 65, 102449. [Google Scholar] [CrossRef]
- Segura-Campos, M.R.; Espinosa-García, L.; Chel-Guerrero, L.A.; Betancur-Ancona, D.A. Effect of Enzymatic Hydrolysis on Solubility, Hydrophobicity, and In Vivo Digestibility in Cowpea (Vigna unguiculata). Int. J. Food Prop. 2012, 15, 770–780. [Google Scholar] [CrossRef] [Green Version]
- Butré, C.I.; Sforza, S.; Wierenga, P.A.; Gruppen, H. Determination of the influence of the pH of hydrolysis on enzyme selectivity of Bacillus licheniformis protease towards whey protein isolate. Int. Dairy J. 2015, 44, 44–53. [Google Scholar] [CrossRef]
- Butré, C.I.; Sforza, S.; Gruppen, H.; Wierenga, P.A. Introducing enzyme selectivity: A quantitative parameter to describe enzymatic protein hydrolysis. Anal. Bioanal. Chem. 2014, 406, 5827–5841. [Google Scholar] [CrossRef] [PubMed]
- Eckert, E.; Han, J.; Swallow, K.; Tian, Z.; Jarpa-Parra, M.; Chen, L. Effects of enzymatic hydrolysis and ultrafiltration on physicochemical and functional properties of faba bean protein. Cereal Chem. 2019, 96, 725–741. [Google Scholar] [CrossRef]
- Schlegel, K.; Sontheimer, K.; Hickisch, A.; Wani, A.A.; Eisner, P.; Schweiggert-Weisz, U. Enzymatic hydrolysis of lupin protein isolates-Changes in the molecular weight distribution, technofunctional characteristics, and sensory attributes. Food Sci. Nutr. 2019, 7, 2747–2759. [Google Scholar] [CrossRef]
- Yust, M.D.M.; Millán-Linares, M.D.C.; Alcaide-Hidalgo, J.M.; Millán, F.; Pedroche, J. Hydrolysis of chickpea proteins with Flavourzyme immobilized on glyoxyl-agarose gels improves functional properties. Food Sci. Technol. Int. 2013, 19, 217–223. [Google Scholar] [CrossRef]
- Yust, M.D.M.; Pedroche, J.; Millán-Linares, M.D.C.; Alcaide-Hidalgo, J.M.; Millán, F. Improvement of functional properties of chickpea proteins by hydrolysis with immobilised Alcalase. Food Chem. 2010, 122, 1212–1217. [Google Scholar] [CrossRef]
- Ewert, J.; Claaßen, W.; Stressler, T.; Fischer, L. An innovative two-step enzymatic membrane bioreactor approach for the continuous production of antioxidative casein hydrolysates with reduced bitterness. Biochem. Eng. J. 2019, 150, 107261. [Google Scholar] [CrossRef]
- Klost, M.; Drusch, S. Functionalisation of pea protein by tryptic hydrolysis—Characterisation of interfacial and functional properties. Food Hydrocoll. 2019, 86, 134–140. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Motta, H. Exploring the structure-function relationship of Great Northern and navy bean (Phaseolus vulgaris L.) protein hydrolysates: A study on the effect of enzymatic hydrolysis. Int. J. Biol. Macromol. 2020, 162, 1516–1525. [Google Scholar] [CrossRef]
- Mokni Ghribi, A.; Maklouf Gafsi, I.; Sila, A.; Blecker, C.; Danthine, S.; Attia, H.; Bougatef, A.; Besbes, S. Effects of enzymatic hydrolysis on conformational and functional properties of chickpea protein isolate. Food Chem. 2015, 187, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Ghribi, A.M.; Gafsi, I.M.; Blecker, C.; Danthine, S.; Attia, H.; Besbes, S. Effect of drying methods on physico-chemical and functional properties of chickpea protein concentrates. J. Food Eng. 2015, 165, 179–188. [Google Scholar] [CrossRef]
- Avramenko, N.A.; Low, N.H.; Nickerson, M.T. The effects of limited enzymatic hydrolysis on the physicochemical and emulsifying properties of a lentil protein isolate. Food Res. Int. 2013, 51, 162–169. [Google Scholar] [CrossRef]
- Paraman, I.; Hettiarachchy, N.S.; Schaefer, C.; Beck, M.I. Hydrophobicity, Solubility, and Emulsifying Properties of Enzyme-Modified Rice Endosperm Protein. Cereal Chem. 2007, 84, 343–349. [Google Scholar] [CrossRef]
- Barac, M.; Cabrilo, S.; Stanojevic, S.; Pesic, M.; Pavlicevic, M.; Zlatkovic, B.; Jankovic, M. Functional properties of protein hydrolysates from pea (Pisum sativum L.) seeds. Int. J. Food Sci. Technol. 2012, 47, 1457–1467. [Google Scholar] [CrossRef]
- Al-Ruwaih, N.; Ahmed, J.; Mulla, M.F.; Arfat, Y.A. High-pressure assisted enzymatic proteolysis of kidney beans protein isolates and characterization of hydrolysates by functional, structural, rheological and antioxidant properties. LWT 2019, 100, 231–236. [Google Scholar] [CrossRef]
- Ahmed, J.; Mulla, M.; Al-Ruwaih, N.; Arfat, Y.A. Effect of high-pressure treatment prior to enzymatic hydrolysis on rheological, thermal, and antioxidant properties of lentil protein isolate. Legume Sci. 2019, 1, e10. [Google Scholar] [CrossRef] [Green Version]
- Spellman, D.; McEvoy, E.; O’Cuinn, G.; FitzGerald, R.J. Proteinase and exopeptidase hydrolysis of whey protein: Comparison of the TNBS, OPA and pH stat methods for quantification of degree of hydrolysis. Int. Dairy J. 2003, 13, 447–453. [Google Scholar] [CrossRef]
- Rutherfurd, S.M. Methodology for Determining Degree of Hydrolysis of Proteins in Hydrolysates: A Review. J. AOAC Int. 2010, 93, 1515–1522. [Google Scholar] [CrossRef] [Green Version]
- Avelar, Z.; Vicente, A.A.; Saraiva, J.A.; Rodrigues, R.M. The role of emergent processing technologies in tailoring plant protein functionality: New insights. Trends Food Sci. Technol. 2021, 113, 219–231. [Google Scholar] [CrossRef]
- Day, L. Proteins from land plants—Potential resources for human nutrition and food security. Trends Food Sci. Technol. 2013, 32, 25–42. [Google Scholar] [CrossRef]
- Burger, T.G.; Zhang, Y. Recent progress in the utilization of pea protein as an emulsifier for food applications. Trends Food Sci. Technol. 2019, 86, 25–33. [Google Scholar] [CrossRef]
- Taherian, A.R.; Mondor, M.; Labranche, J.; Drolet, H.; Ippersiel, D.; Lamarche, F. Comparative study of functional properties of commercial and membrane processed yellow pea protein isolates. Food Res. Int. 2011, 44, 2505–2514. [Google Scholar] [CrossRef]
- Tavano, O.L. Protein hydrolysis using proteases: An important tool for food biotechnology. J. Mol. Catal. B Enzym. 2013, 90, 1–11. [Google Scholar] [CrossRef]
- Sathe, S.K.; Zaffran, V.D.; Gupta, S.; Li, T. Protein Solubilization. J. Am. Oil Chem. Soc. 2018, 95, 883–901. [Google Scholar] [CrossRef]
- Nicolai, T.; Chassenieux, C. Heat-induced gelation of plant globulins. Curr. Opin. Food Sci. 2019, 27, 18–22. [Google Scholar] [CrossRef]
- Saricaoglu, F.T. Application of high-pressure homogenization (HPH) to modify functional, structural and rheological properties of lentil (Lens culinaris) proteins. Int. J. Biol. Macromol. 2020, 144, 760–769. [Google Scholar] [CrossRef] [PubMed]
- Betancur-Ancona, D.; Martínez-Rosado, R.; Corona-Cruz, A.; Castellanos-Ruelas, A.; Jaramillo-Flores, M.E.; Chel-Guerrero, L. Functional properties of hydrolysates fromPhaseolus lunatusseeds. Int. J. Food Sci. Technol. 2009, 44, 128–137. [Google Scholar] [CrossRef]
- Konieczny, D.; Stone, A.K.; Korber, D.R.; Nickerson, M.T.; Tanaka, T. Physicochemical properties of enzymatically modified pea protein-enriched flour treated by different enzymes to varying levels of hydrolysis. Cereal Chem. 2020, 97, 326–338. [Google Scholar] [CrossRef]
- Samaei, S.P.; Ghorbani, M.; Tagliazucchi, D.; Martini, S.; Gotti, R.; Themelis, T.; Tesini, F.; Gianotti, A.; Gallina Toschi, T.; Babini, E. Functional, nutritional, antioxidant, sensory properties and comparative peptidomic profile of faba bean (Vicia faba, L.) seed protein hydrolysates and fortified apple juice. Food Chem. 2020, 330, 127120. [Google Scholar] [CrossRef]
- Barać, M.; Čabrilo, S.; Pešić, M.; Stanojević, S.; Pavlićević, M.; Maćej, O.; Ristić, N. Functional Properties of Pea (Pisum sativum, L.) Protein Isolates Modified with Chymosin. Int. J. Mol. Sci. 2011, 12, 8372–8387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Galanopoulos, M.; Sismour, E.; Ren, S.; Mersha, Z.; Lynch, P.; Almutaimi, A. Effect of enzymatic hydrolysis using endo- and exo-proteases on secondary structure, functional, and antioxidant properties of chickpea protein hydrolysates. J. Food Meas. Charact. 2020, 14, 343–352. [Google Scholar] [CrossRef]
- McClements, D.J.; Bai, L.; Chung, C. Recent Advances in the Utilization of Natural Emulsifiers to Form and Stabilize Emulsions. Annu. Rev. Food Sci. Technol. 2017, 8, 205–236. [Google Scholar] [CrossRef]
- Damodaran, S. Protein Stabilization of Emulsions and Foams. J. Food Sci. 2005, 70, R54–R66. [Google Scholar] [CrossRef]
- Gumus, C.E.; Decker, E.A.; McClements, D.J. Formation and Stability of ω-3 Oil Emulsion-Based Delivery Systems Using Plant Proteins as Emulsifiers: Lentil, Pea, and Faba Bean Proteins. Food Biophys. 2017, 12, 186–197. [Google Scholar] [CrossRef]
- Ma, Z.; Boye, J.I.; Simpson, B.K. Preparation of Salad Dressing Emulsions Using Lentil, Chickpea and Pea Protein Isolates: A Response Surface Methodology Study. J. Food Qual. 2016, 39, 274–291. [Google Scholar] [CrossRef]
- Vogelsang-O’Dwyer, M.; Sahin, A.W.; Zannini, E.; Arendt, E.K. Physicochemical and nutritional properties of high protein emulsion-type lupin-based model milk alternatives: Effect of protein source and homogenization pressure. J. Sci. Food Agric. 2021. [Google Scholar] [CrossRef]
- Pearce, K.N.; Kinsella, J.E. Emulsifying properties of proteins: Evaluation of a turbidimetric technique. J. Agric. Food Chem. 1978, 26, 716–723. [Google Scholar] [CrossRef]
- Tabilo-Munizaga, G.; Villalobos-Carvajal, R.; Herrera-Lavados, C.; Moreno-Osorio, L.; Jarpa-Parra, M.; Pérez-Won, M. Physicochemical properties of high-pressure treated lentil protein-based nanoemulsions. LWT 2019, 101, 590–598. [Google Scholar] [CrossRef]
- Evangelho, J.A.D.; Vanier, N.L.; Pinto, V.Z.; Berrios, J.J.D.; Dias, A.R.G.; Zavareze, E.D.R. Black bean (Phaseolus vulgaris L.) protein hydrolysates: Physicochemical and functional properties. Food Chem. 2017, 214, 460–467. [Google Scholar] [CrossRef]
- Wani, I.A.; Sogi, D.S.; Shivhare, U.S.; Gill, B.S. Physico-chemical and functional properties of native and hydrolyzed kidney bean (Phaseolus vulgaris L.) protein isolates. Food Res. Int. 2015, 76, 11–18. [Google Scholar] [CrossRef]
- Wani, I.A.; Sogi, D.S.; Gill, B.S. Physico-chemical and functional properties of native and hydrolysed protein isolates from Indian black gram (Phaseolus mungo L.) cultivars. LWT—Food Sci. Technol. 2015, 60, 848–854. [Google Scholar] [CrossRef]
- Foegeding, E.A.; Davis, J.P. Food protein functionality: A comprehensive approach. Food Hydrocoll. 2011, 25, 1853–1864. [Google Scholar] [CrossRef]
- Liu, C.; Damodaran, S.; Heinonen, M. Effects of microbial transglutaminase treatment on physiochemical properties and emulsifying functionality of faba bean protein isolate. LWT 2019, 99, 396–403. [Google Scholar] [CrossRef] [Green Version]
- Lqari, H.; Pedroche, J.; Girón-Calle, J.; Vioque, J.; Millán, F. Production of Lupinus angustifolius protein hydrolysates with improved functional properties. Grasas Y Aceites 2005, 56, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Grossmann, L.; McClements, D.J. The science of plant-based foods: Approaches to create nutritious and sustainable plant-based cheese analogs. Trends Food Sci. Technol. 2021, 118, 207–229. [Google Scholar] [CrossRef]
- Gharibzahedi, S.M.T.; Smith, B. Effects of high hydrostatic pressure on the quality and functionality of protein isolates, concentrates, and hydrolysates derived from pulse legumes: A review. Trends Food Sci. Technol. 2021, 107, 466–479. [Google Scholar] [CrossRef]
- Berghout, J.A.M.; Boom, R.M.; van der Goot, A.J. Understanding the differences in gelling properties between lupin protein isolate and soy protein isolate. Food Hydrocoll. 2015, 43, 465–472. [Google Scholar] [CrossRef]
- Guldiken, B.; Stobbs, J.; Nickerson, M. Heat induced gelation of pulse protein networks. Food Chem. 2021, 350, 129158. [Google Scholar] [CrossRef]
- Ju, Z.Y.; Otte, J.; Madsen, J.S.; Qvist, K.B. Effects of Limited Proteolysis on Gelation and Gel Properties of Whey Protein Isolate. J. Dairy Sci. 1995, 78, 2119–2128. [Google Scholar] [CrossRef]
- Lamsal, B.; Jung, S.; Johnson, L. Rheological properties of soy protein hydrolysates obtained from limited enzymatic hydrolysis. LWT—Food Sci. Technol. 2007, 40, 1215–1223. [Google Scholar] [CrossRef]
- Felix, M.; Perez-Puyana, V.; Romero, A.; Guerrero, A. Development of thermally processed bioactive pea protein gels: Evaluation of mechanical and antioxidant properties. Food Bioprod. Process. 2017, 101, 74–83. [Google Scholar] [CrossRef]
- Tanger, C.; Müller, M.; Andlinger, D.; Kulozik, U. Influence of pH and ionic strength on the thermal gelation behaviour of pea protein. Food Hydrocoll. 2021, 123, 106903. [Google Scholar] [CrossRef]
- Klost, M.; Giménez-Ribes, G.; Drusch, S. Enzymatic hydrolysis of pea protein: Interactions and protein fractions involved in fermentation induced gels and their influence on rheological properties. Food Hydrocoll. 2020, 105, 105793. [Google Scholar] [CrossRef]
- Bajaj, P.R.; Bhunia, K.; Kleiner, L.; Joyner, H.S.; Smith, D.; Ganjyal, G.; Sablani, S.S. Improving functional properties of pea protein isolate for microencapsulation of flaxseed oil. J. Microencapsul. 2017, 34, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Cho, M.J.; Unklesbay, N.; Hsieh, F.-H.; Clarke, A.D. Hydrophobicity of Bitter Peptides from Soy Protein Hydrolysates. J. Agric. Food Chem. 2004, 52, 5895–5901. [Google Scholar] [CrossRef]
- Humiski, L.M.; Aluko, R.E. Physicochemical and bitterness properties of enzymatic pea protein hydrolysates. J. Food Sci. 2007, 72, S605–S611. [Google Scholar] [CrossRef] [PubMed]
- Raksakulthai, R.; Haard, N.F. Exopeptidases and Their Application to Reduce Bitterness in Food: A Review. Crit. Rev. Food Sci. Nutr. 2003, 43, 401–445. [Google Scholar] [CrossRef]
- Maehashi, K.; Matano, M.; Wang, H.; Vo, L.A.; Yamamoto, Y.; Huang, L. Bitter peptides activate hTAS2Rs, the human bitter receptors. Biochem. Biophys. Res. Commun. 2008, 365, 851–855. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, T.; Kato, H. Taste of free amino acids and peptides. Food Rev. Int. 1988, 4, 175–194. [Google Scholar] [CrossRef]
- Aluko, R.E. Structural Characteristics of Food Protein-Derived Bitter Peptides. In Bitterness: Perception, Chemistry and Food Processing; John Wiley & Sons: Hoboken, NJ, USA, 2017; pp. 105–129. [Google Scholar] [CrossRef]
- Seo, W.H.; Lee, H.G.; Baek, H.H. Evaluation of Bitterness in Enzymatic Hydrolysates of Soy Protein Isolate by Taste Dilution Analysis. J. Food Sci. 2008, 73, S41–S46. [Google Scholar] [CrossRef] [PubMed]
- Meinlschmidt, P.; Sussmann, D.; Schweiggert-Weisz, U.; Eisner, P. Enzymatic treatment of soy protein isolates: Effects on the potential allergenicity, technofunctionality, and sensory properties. Food Sci. Nutr. 2016, 4, 11–23. [Google Scholar] [CrossRef]
- Schlegel, K.; Sontheimer, K.; Eisner, P.; Schweiggert-Weisz, U. Effect of enzyme-assisted hydrolysis on protein pattern, technofunctional, and sensory properties of lupin protein isolates using enzyme combinations. Food Sci. Nutr. 2020, 8, 3041–3051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meinlschmidt, P.; Schweiggert-Weisz, U.; Brode, V.; Eisner, P. Enzyme assisted degradation of potential soy protein allergens with special emphasis on the technofunctionality and the avoidance of a bitter taste formation. LWT—Food Sci. Technol. 2016, 68, 707–716. [Google Scholar] [CrossRef]
- Rezvankhah, A.; Yarmand, M.S.; Ghanbarzadeh, B.; Mirzaee, H. Characterization of bioactive peptides produced from green lentil (Lens culinaris) seed protein concentrate using Alcalase and Flavourzyme in single and sequential hydrolysis. J. Food Process. Preserv. 2021, 45, e15932. [Google Scholar] [CrossRef]
- Ewert, J.; Schlierenkamp, F.; Nesensohn, L.; Fischer, L.; Stressler, T. Improving the colloidal and sensory properties of a caseinate hydrolysate using particular exopeptidases. Food Funct. 2018, 9, 5989–5998. [Google Scholar] [CrossRef] [PubMed]
- Großmann, K.K.; Merz, M.; Appel, D.; Thaler, T.; Fischer, L. Impact of Peptidase Activities on Plant Protein Hydrolysates Regarding Bitter and Umami Taste. J. Agric. Food Chem. 2021, 69, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Bhaskar, B.; Ananthanarayan, L.; Jamdar, S.N. Effect of enzymatic hydrolysis on the functional, antioxidant, and angiotensin I-converting enzyme (ACE) inhibitory properties of whole horse gram flour. Food Sci. Biotechnol. 2019, 28, 43–52. [Google Scholar] [CrossRef]
- Li, Q.; Yi, L.; Marek, P.; Iverson, B.L. Commercial proteases: Present and future. FEBS Lett. 2013, 587, 1155–1163. [Google Scholar] [CrossRef] [Green Version]


| Enzyme Preparation | Main Activity | Origin |
|---|---|---|
| Alcalase | Serine endoprotease; broad specificity, preferentially hydrolyses peptide bonds at the C-terminal side of hydrophobic residues | Bacillus lichenformis |
| Trypsin | Serine endoprotease; specific for peptide bonds at the C-terminal side of Lys and Arg residues | Bovine/porcine pancreas |
| Chymotrypsin | Serine endoprotease; preferentially hydrolyses peptide bonds at the C-terminal side of Tyr, Phe, Trp and Leu residues | Bovine/porcine pancreas |
| Savinase | Serine endoprotease, broad specificity | Bacillus lentus |
| Protamex | Broad specificity endoprotease | Bacillus sp. |
| Corolase 2TS | Metallo endoprotease | Bacillus thermoproteolyticus, Bacillus stearothermophilus |
| Neutrase | Metallo endoprotease | Bacillus amyloliquefaciens |
| Pepsin | Aspartic endoprotease, broad specificity | Bovine/porcine gastric mucosa |
| Papain | Cysteine endoprotease, broad specificity | Papaya latex |
| Bromelain | Cysteine endoprotease, broad specificity | Pineapple stem |
| Flavourzyme | Exo and endoprotease mixture. Includes aminopeptidases, carboxypeptidases, and endoproteases | Aspergillus oryzae |
| Reference | Protein Source | Protease | Effect on Foaming Properties |
|---|---|---|---|
| Ahmed et al. [52] | Lentil protein isolate | Alcalase | FC: increased; FS: decreased |
| Al-Ruwaih et al. [51] | Kidney bean protein isolate | Alcalase | FC: increased (but decreased for high pressure treated sample) FS: decreased |
| Barać et al. [66] | Pea protein isolate | Chymosin | FC: increased at pH 3, 5, and 7; increased/decreased at pH 8 depending on HT FS: increased/decreased at pH 3 and 5 depending on HT; increased/no difference at pH 7 depending on HT; decreased at pH 8 |
| Barac et al. [50] | Pea protein isolate (L1) | Papain | FC: increased at pH 3, 5, 7, and 8 FS: increased/decreased at pH 3 and 5 depending on HT; increased at pH 7; increased/decreased at pH 8 depending on HT |
| S. griseus protease | FC: increased at pH 3 and 5; increased/decreased at pH 7 depending on HT; decreased at pH 8 FS: decreased at pH 3 and 5; no difference at pH 7; decreased at pH 8 | ||
| Pea protein isolate (Maja) | Papain | FC: increased at pH 3, 5, and 7; decreased/no difference at pH 8 depending on HT FS: increased at pH 3, 5, 7 and 8 | |
| S. griseus protease | FC: increased/decreased at pH 3, 5, and 7 depending on HT; decreased at pH 8 FS: increased/no difference at pH 3 depending on HT; increased/decreased at pH 5 and 7 depending on HT; increased at pH 8 | ||
| Betancur-Ancona et al. [63] | P. lunatus protein isolate | Alcalase | FC: decreased at pH 2, 4, 6, 8, and 10 FS: increased/decreased at pH 2 and 4 depending on HT; increased at pH 6 and 8; increased at pH 10 |
| Flavourzyme | FC: increased at pH 2, 4, 6, 8, and 10 FS: increased at pH 2, 4, 6, 8, and 10 | ||
| Eckert et al. [39] | Faba bean protein isolate | Pepsin | FC: increased at pH 5 and 7 FS: no difference at pH 5; increased/no difference at pH 7 depending on HT |
| Trypsin | FC: increased at pH 5; increased/no difference at pH 7 depending on HT FS: no difference at pH 5; increased at pH 7 | ||
| Flavourzyme | FC: increased at pH 5 and 7 FS: no difference at pH 5; increased at pH 7 | ||
| Neutrase | FC: increased at pH 5; increased/no difference at pH 7 depending on HT FS: increased/decreased at pH 5 and 7 depending on HT | ||
| Konieczny et al. [64] | Pea protein-enriched flour | Trypsin | FC: decreased at pH 4, 7, and 10 FS: decreased/no difference at pH 4 depending on DH; increased/no difference at pH 7 depending on DH; increased at pH 10 |
| Savinase | FC: decreased at pH 4, 7, and 10 FS: decreased/no difference at pH 4 depending on DH; increased at pH 7; increased/no difference at pH 10 depending on DH | ||
| Papain | FC: increased/decreased at pH 4 depending on DH; decreased at pH 7 and 10 FS: no difference at pH 4; increased/no difference at pH 7 depending on DH; increased at pH 10 | ||
| Pepsin | FC: increased no/difference at pH 4 depending on DH; decreased at pH 7; decreased/no difference at pH 10 FS: increased at pH 4, 7, and 10 | ||
| Schlegel et al. [40] | Lupin protein isolate | Alcalase | FC: increased; FS: no difference |
| Papain | FC: increased; FS: no difference | ||
| Neutrase | FC: increased; FS: no difference | ||
| Protease N-01 | FC: increased; FS: no difference | ||
| Flavourzyme | FC: no difference; FS: no difference | ||
| Protamex | FC: increased; FS: no difference | ||
| Corolase 7089 | FC: increased; FS: no difference | ||
| Pepsin | FC: increased; FS: no difference | ||
| Corolase N | FC: increased; FS: no difference | ||
| Yust et al. [42] | Chickpea protein isolate | Alcalase | FC: increased; FS: increased |
| Yust et al. [41] | Chickpea protein isolate | Flavourzyme | FC: increased; FS: increased/no difference depending on DH |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vogelsang-O’Dwyer, M.; Sahin, A.W.; Arendt, E.K.; Zannini, E. Enzymatic Hydrolysis of Pulse Proteins as a Tool to Improve Techno-Functional Properties. Foods 2022, 11, 1307. https://doi.org/10.3390/foods11091307
Vogelsang-O’Dwyer M, Sahin AW, Arendt EK, Zannini E. Enzymatic Hydrolysis of Pulse Proteins as a Tool to Improve Techno-Functional Properties. Foods. 2022; 11(9):1307. https://doi.org/10.3390/foods11091307
Chicago/Turabian StyleVogelsang-O’Dwyer, Martin, Aylin W. Sahin, Elke K. Arendt, and Emanuele Zannini. 2022. "Enzymatic Hydrolysis of Pulse Proteins as a Tool to Improve Techno-Functional Properties" Foods 11, no. 9: 1307. https://doi.org/10.3390/foods11091307
APA StyleVogelsang-O’Dwyer, M., Sahin, A. W., Arendt, E. K., & Zannini, E. (2022). Enzymatic Hydrolysis of Pulse Proteins as a Tool to Improve Techno-Functional Properties. Foods, 11(9), 1307. https://doi.org/10.3390/foods11091307