
Oxidative Stress Contributes to Cytoskeletal Protein Degradation of Esox lucius through Activation of Mitochondrial Apoptosis during Postmortem Storage


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Myofibrillar Protein (MP) Extraction
2.3. Mitochondrial Protein Extraction
2.3.1. Measurement of Mitochondrial ROS
2.3.2. Determination of Mitochondrial Antioxidant Enzyme Activity and MDA Levels
2.3.3. Detection of MPTP Opening and MMP
2.3.4. Measurement of Mitochondrial Swelling
2.4. Ultrastructural Observation of Mitochondria
2.5. SDS-PAGE and Immunoblot Analysis
2.6. Analysis of Caspases and Cytochrome C Redox State of Mitochondria
2.7. Statistical Analysis
3. Results and Discussion
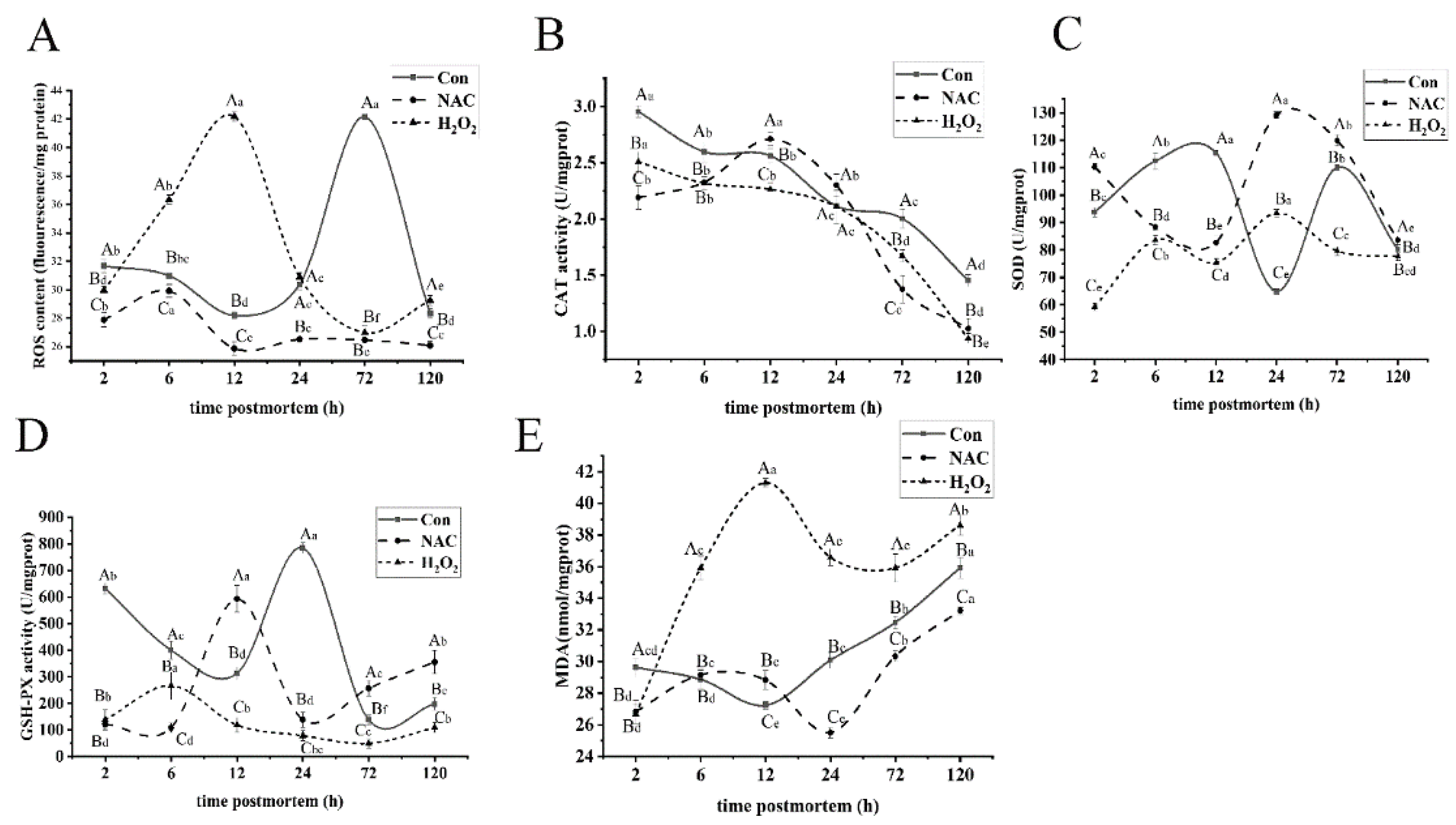
3.1. ROS Induced Mitochondrial Oxidative Stress
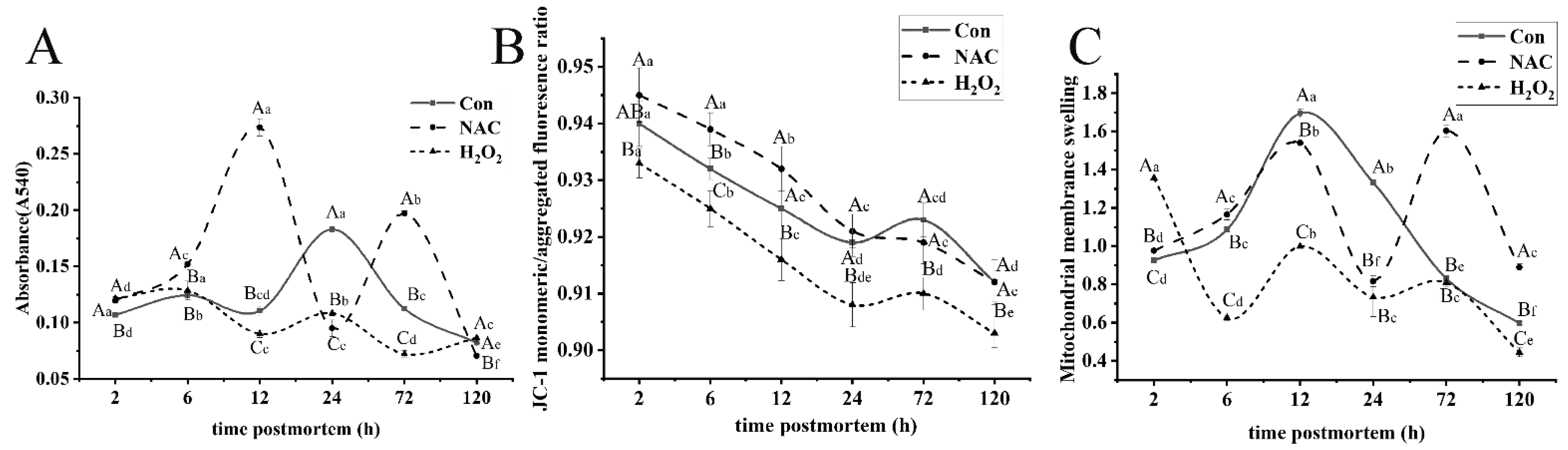
3.2. Changes in Mitochondrial Dysfunction
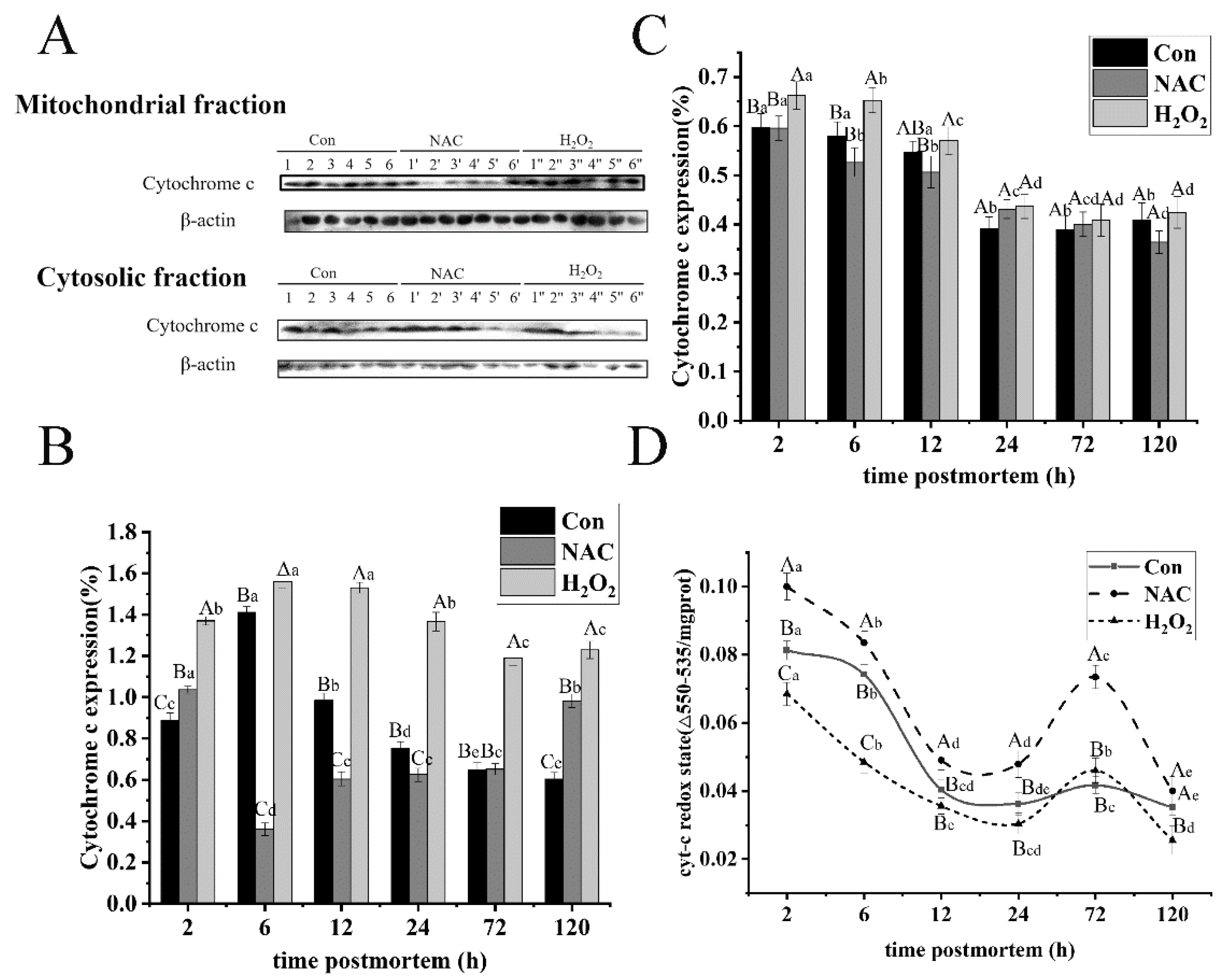
3.3. ROS Promoted Cytochrome C Expression and Oxidative Redox Level
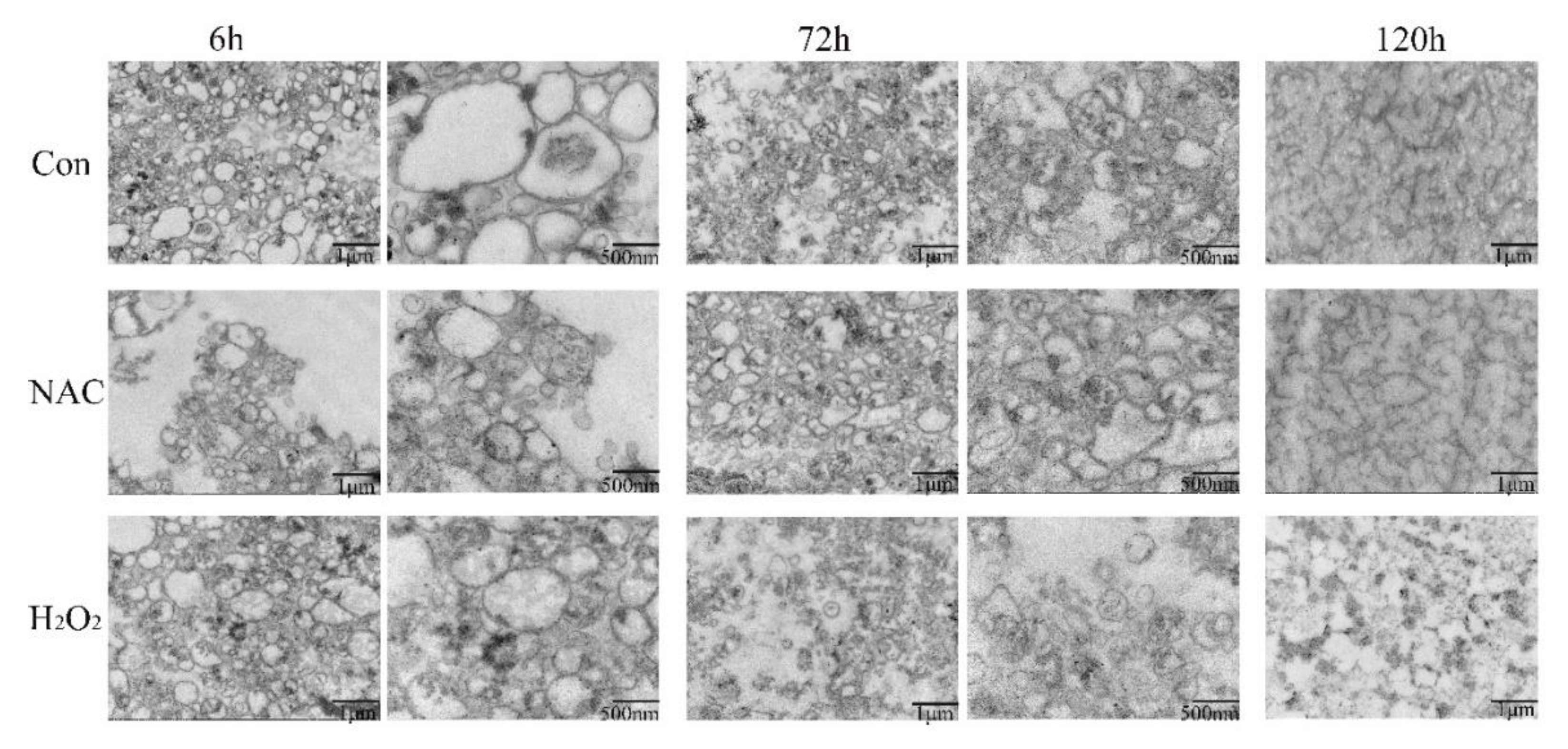
3.4. Mitochondrial Morphology
3.5. Effect of ROS-Mediated Structural Protein Degradation
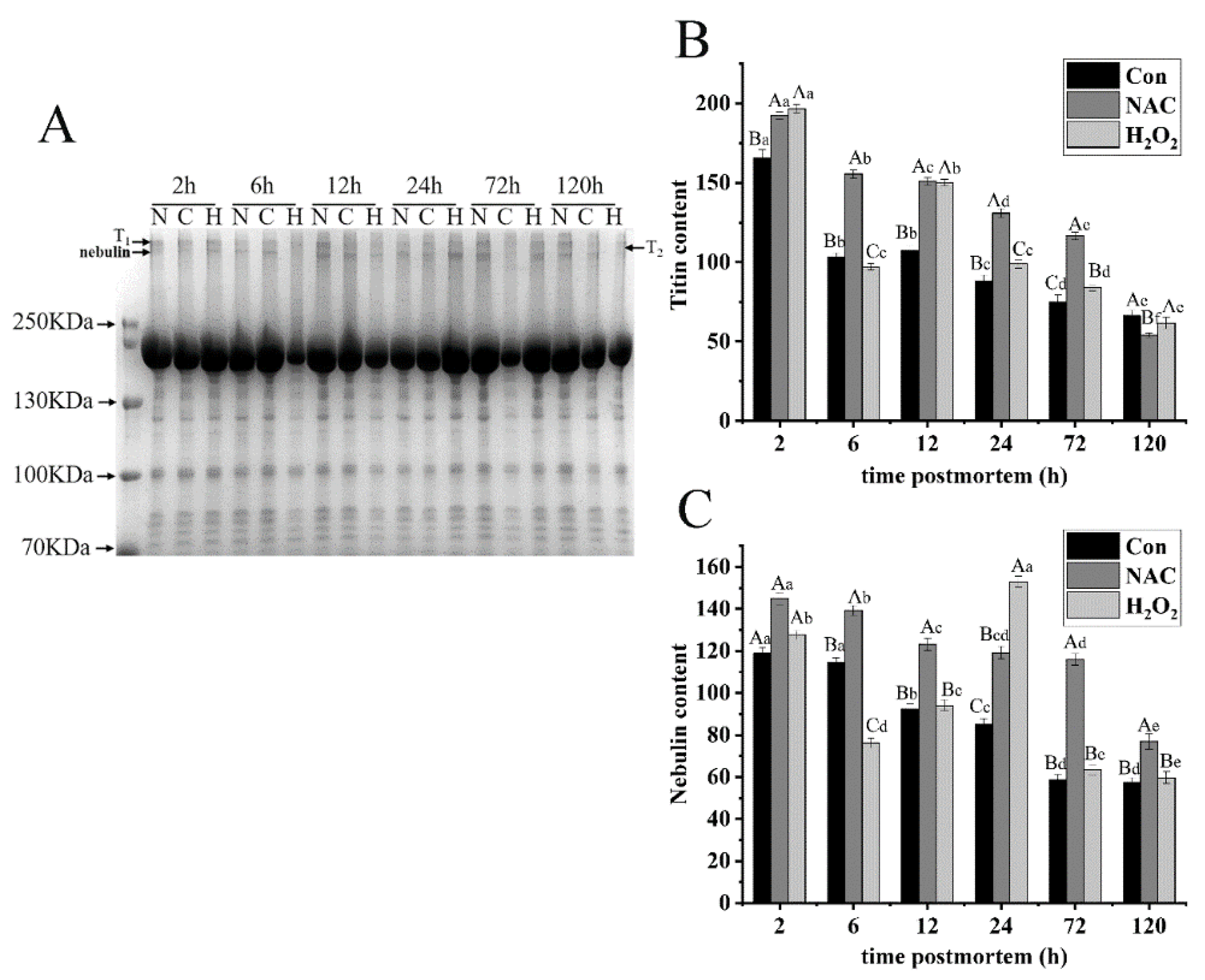
3.5.1. Titin and Nebulin
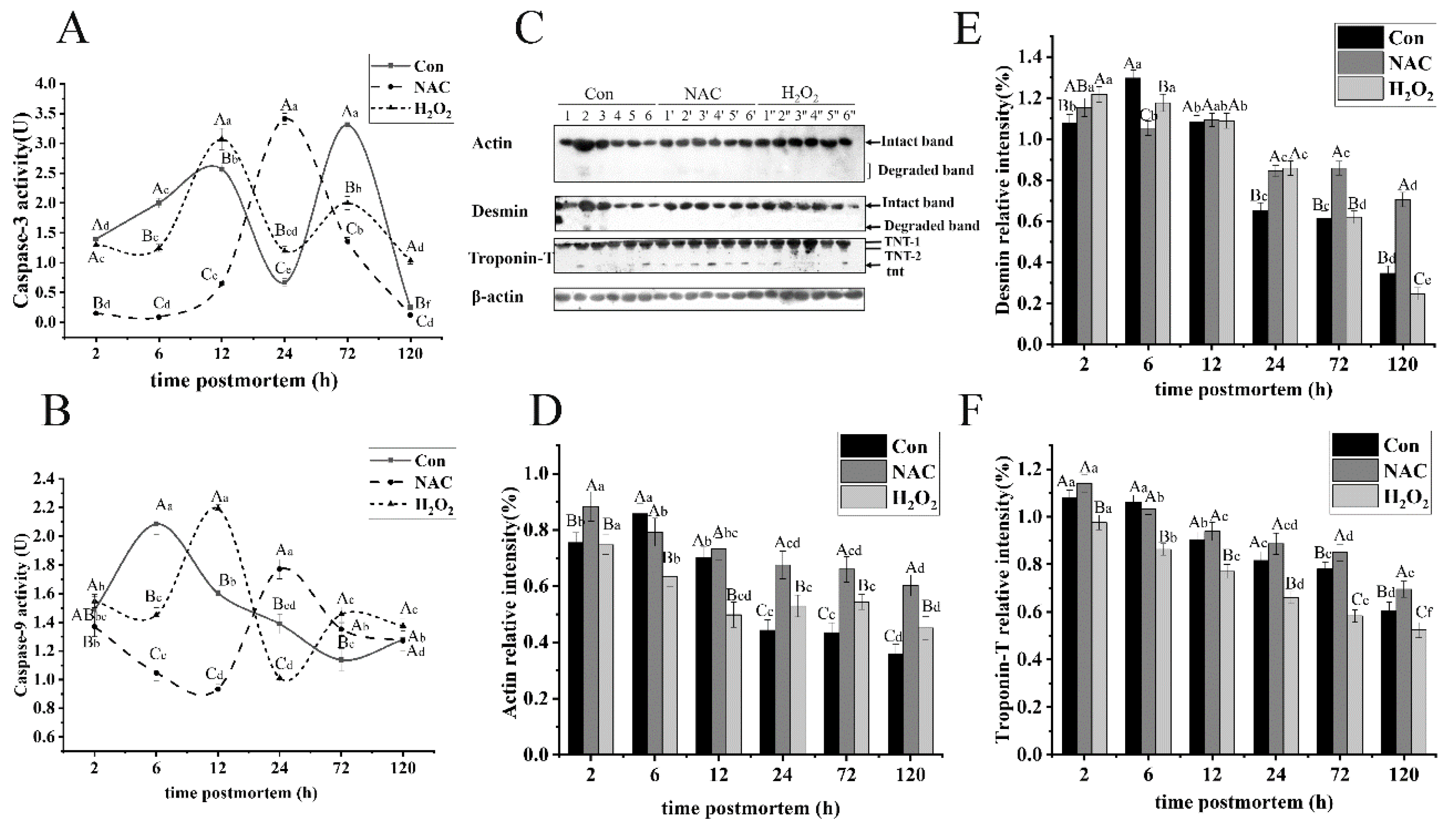
3.5.2. Actin
3.5.3. Desmin
3.5.4. Troponin-T
3.6. Changes in Caspase Activity during Postmortem Storage
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Yu, D.; Regenstein, J.M.; Zang, J.; Xia, W.; Xu, Y.; Jiang, Q.; Yang, F. Inhibitory effects of chitosan-based coatings on endogenous enzyme activities, proteolytic degradation and texture softening of grass carp (Ctenopharyngodon idellus) fillets stored at 4 °C. Food Chem. 2018, 262, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Moslemi-Aqdam, M.; Baker, L.F.; Baltzer, J.L.; Branfireun, B.A.; Evans, M.S.; Laird, B.D.; Low, G.; Low, M.; Swanson, H.K. Understanding among-lake variability of mercury concentrations in Northern Pike (Esox lucius): A whole-ecosystem study in subarctic lakes. Sci. Total Environ. 2022, 822, 153430. [Google Scholar] [CrossRef] [PubMed]
- Cejko, B.I.; Sarosiek, B.; Dryl, K.; Judycka, S.; Szczepkowska, B.; Szczepkowski, M.; Kowalski, R.K. The effect of cryopreservation extender on sperm motility and hatch success in northern pike (Esox lucius). Aquaculture 2020, 514, 734482. [Google Scholar] [CrossRef]
- Pietrzak-Fiećko, R.; Modzelewska-Kapituła, M.; Zakęś, Z.; Szczepkowski, M. The Effect of Thermal Treatment Method on Fatty Acid Composition in Northern Pike (Esox lucius) Fillets. J. Aquat. Food Prod. Technol. 2017, 26, 1303–1311. [Google Scholar] [CrossRef]
- Rigdon, M.; Hung, Y.C.; Stelzleni, A.M. Evaluation of alkaline electrolyzed water to replace traditional phosphate enhancement solutions: Effects on water holding capacity, tenderness, and sensory characteristics. Meat Sci. 2017, 123, 211–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Feng, X.C.; Lu, F.; Xu, X.L.; Zhou, G.H.; Li, Q.Y.; Guo, X.Y. Effects of camptothecin, etoposide and Ca2+ on caspase-3 activity and myofibrillar disruption of chicken during postmortem ageing. Meat Sci. 2011, 87, 165–174. [Google Scholar] [CrossRef]
- Gupta, S.; Gollapudi, S. Susceptibility of naïve and subsets of memory T cells to apoptosis via multiple signaling pathways. Autoimmun. Rev. 2007, 6, 476–481. [Google Scholar] [CrossRef]
- Dorstyn, L.; Read, S.; Cakouros, D.; Huh, J.R.; Hay, B.A.; Kumar, S. The role of cytochrome c in caspase activation in Drosophila melanogaster cells. J. Cell Biol. 2002, 156, 1089–1098. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Guo, Z.; Ma, G.; Ma, J.; Han, L. Lysosomal Fe2+ contributes to myofibrillar protein degradation through mitochondrial-dysfunction-induced apoptosis. LWT—Food Sci. Technol. 2021, 143, 111197. [Google Scholar] [CrossRef]
- Li, X.; Hu, L.; Zhu, X.; Guo, X.; Deng, X.; Zhang, J. The effect of caspase-3 in mitochondrial apoptosis activation on degradation of structure proteins of Esox lucius during postmortem storage. Food Chem. 2022, 367, 130767. [Google Scholar] [CrossRef]
- Kinnally, K.W.; Peixoto, P.M.; Ryu, S.Y.; Dejean, L.M. Is mPTP the gatekeeper for necrosis, apoptosis, or both? Biochim. Biophys. Acta 2011, 1813, 616–622. [Google Scholar] [CrossRef] [Green Version]
- Bou-Teen, D.; Kaludercic, N.; Weissman, D.; Turan, B.; Maack, C.; Di Lisa, F.; Ruiz-Meana, M. Mitochondrial ROS and mitochondria-targeted antioxidants in the aged heart. Free Radic. Biol. Med. 2021, 167, 109–124. [Google Scholar] [CrossRef]
- Pasdois, P.; Parker, J.E.; Griffiths, E.J.; Halestrap, A.P. The role of oxidized cytochrome c in regulating mitochondrial reactive oxygen species production and its perturbation in ischaemia. Biochem. J. 2011, 436, 493–505. [Google Scholar] [CrossRef] [Green Version]
- Kozakowska, M.; Pietraszek-Gremplewicz, K.; Jozkowicz, A.; Dulak, J. The role of oxidative stress in skeletal muscle injury and regeneration: Focus on antioxidant enzymes. J. Muscle Res. Cell Motil. 2015, 36, 377–393. [Google Scholar] [CrossRef] [Green Version]
- Aparicio-Trejo, O.E.; Reyes-Fermín, L.M.; Briones-Herrera, A.; Tapia, E.; León-Contreras, J.C.; Hernández-Pando, R.; Sánchez-Lozada, L.G.; Pedraza-Chaverri, J. Protective effects of N-acetyl-cysteine in mitochondria bioenergetics, oxidative stress, dynamics and S-glutathionylation alterations in acute kidney damage induced by folic acid. Free Radic. Biol. Med. 2019, 130, 379–396. [Google Scholar] [CrossRef]
- Chen, X.; Gu, R.; Zhang, L.; Li, J.; Jiang, Y.; Zhou, G.; Gao, F. Induction of nuclear factor-κB signal-mediated apoptosis and autophagy by reactive oxygen species is associated with hydrogen peroxide-impaired growth performance of broilers. Animal 2018, 12, 2561–2570. [Google Scholar] [CrossRef]
- Kemp, C.M.; Wheeler, T.L. Effects of manipulation of the caspase system on myofibrillar protein degradation in vitro. J. Anim. Sci. 2011, 89, 3262–3271. [Google Scholar] [CrossRef]
- Bao, Y.; Wang, K.; Yang, H.; Regenstein, J.M.; Ertbjerg, P.; Zhou, P. Protein degradation of black carp (Mylopharyngodon piceus) muscle during cold storage. Food Chem. 2020, 308, 125576. [Google Scholar] [CrossRef]
- Wang, L.; Yu, Q.; Han, L.; Ma, X.; Zhang, W. Study on the effect of reactive oxygen species-mediated oxidative stress on the activation of mitochondrial apoptosis and the tenderness of yak meat. Food Chem. 2018, 244, 394–402. [Google Scholar] [CrossRef]
- Chen, C.; Zhang, J.; Guo, Z.; Shi, X.; Zhang, Y.; Zhang, L.; Yu, Q.; Han, L. Effect of oxidative stress on AIF-mediated apoptosis and bovine muscle tenderness during postmortem aging. J. Food Sci. 2020, 85, 77–85. [Google Scholar] [CrossRef]
- Pan, J.; Lian, H.; Jia, H.; Li, S.; Hao, R.; Wang, Y.; Zhang, X.; Dong, X. Ultrasound treatment modified the functional mode of gallic acid on properties of fish myofibrillar protein. Food Chem. 2020, 320, 126637. [Google Scholar] [CrossRef]
- Zhang, J.; Yu, Q.; Han, L.; Chen, C.; Li, H.; Han, G. Study on the apoptosis mediated by cytochrome c and factors that affect the activation of bovine longissimus muscle during postmortem aging. Apoptosis 2017, 22, 777–785. [Google Scholar] [CrossRef]
- Zhang, R.; Humphreys, I.; Sahu, R.P.; Shi, Y.; Srivastava, S.K. In vitro and in vivo induction of apoptosis by capsaicin in pancreatic cancer cells is mediated through ROS generation and mitochondrial death pathway. Apoptosis 2008, 13, 1465–1478. [Google Scholar] [CrossRef]
- Wang, L.; Yu, Q.; Han, L.; Ma, X.; Zhao, S.-N. Effect of mitochondrial apoptotic activation through the mitochondrial membrane permeability transition pore on yak meat tenderness during postmortem aging. Food Chem. 2017, 234, 323–331. [Google Scholar] [CrossRef]
- Huang, J.; Lv, C.; Hu, M.; Zhong, G. The mitochondria-mediate apoptosis of Lepidopteran cells induced by azadirachtin. PLoS ONE 2013, 8, e58499. [Google Scholar] [CrossRef] [Green Version]
- Yu, Q.; Tian, X.; Shao, L.; Li, X.; Dai, R. Mitochondria changes and metabolome differences of bovine longissimus lumborum and psoas major during 24 h postmortem. Meat Sci. 2020, 166, 108112. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, X.; Du, Y.; Ji, H.; Li, S.; Li, L.; Xing, Y.; Zhang, X.; Dong, L.; Wang, C.; et al. Leonurine protects brain injury by increased activities of UCP4, SOD, CAT and Bcl-2, decreased levels of MDA and Bax, and ameliorated ultrastructure of mitochondria in experimental stroke. Brain Res. 2012, 1474, 73–81. [Google Scholar] [CrossRef]
- Hu, Y.-m.; Lu, S.-z.; Li, Y.-s.; Wang, H.; Shi, Y.; Zhang, L.; Tu, Z.-c. Protective effect of antioxidant peptides from grass carp scale gelatin on the H2O2-mediated oxidative injured HepG2 cells. Food Chem. 2021, 373, 131539. [Google Scholar] [CrossRef] [PubMed]
- Shieh, P.; Jan, C.R.; Liang, W.Z. The protective effects of the antioxidant N-acetylcysteine (NAC) against oxidative stress-associated apoptosis evoked by the organophosphorus insecticide malathion in normal human astrocytes. Toxicology 2019, 417, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Seydi, E.; Mehrpouya, L.; Sadeghi, H.; Rahimi, S.; Pourahmad, J. Luteolin attenuates Fipronil-induced neurotoxicity through reduction of the ROS-mediated oxidative stress in rat brain mitochondria. Pestic. Biochem. Physiol. 2021, 173, 104785. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Sun, B.; Luo, X.; Zhao, M.; Zheng, F.; Sun, J.; Li, H.; Sun, X.; Huang, M. Cytoprotective effects of a tripeptide from Chinese Baijiu against AAPH-induced oxidative stress in HepG2 cells via Nrf2 signaling. RSC Adv. 2018, 8, 10898–10906. [Google Scholar] [CrossRef] [Green Version]
- Javadov, S.; Chapa-Dubocq, X.; Makarov, V. Different approaches to modeling analysis of mitochondrial swelling. Mitochondrion 2018, 38, 58–70. [Google Scholar] [CrossRef]
- Sabharwal, S.S.; Schumacker, P.T. Mitochondrial ROS in cancer: Initiators, amplifiers or an Achilles’ heel? Nat. Rev. Cancer 2014, 14, 709–721. [Google Scholar] [CrossRef] [Green Version]
- Huang, F.; Huang, M.; Zhang, H.; Zhang, C.; Zhou, G. Changes in apoptotic factors and caspase activation pathways during the postmortem aging of beef muscle. Food Chem. 2016, 190, 110–114. [Google Scholar] [CrossRef]
- Wang, L.; Ma, G.; Zhang, Y.; Shi, X.; Han, L.; Yu, Q.; Zhao, S.; Ma, J. Effect of mitochondrial cytochrome c release and its redox state on the mitochondrial-dependent apoptotic cascade reaction and tenderization of yak meat during postmortem aging. Food Res. Int. 2018, 111, 488–497. [Google Scholar] [CrossRef]
- Duranova, H.; Valkova, V.; Knazicka, Z.; Olexikova, L.; Vasicek, J. Mitochondria: A worthwhile object for ultrastructural qualitative characterization and quantification of cells at physiological and pathophysiological states using conventional transmission electron microscopy. Acta Histochem. 2020, 122, 151646. [Google Scholar] [CrossRef]
- Huang, M.; Huang, F.; Xue, M.; Xu, X.; Zhou, G. The effect of active caspase-3 on degradation of chicken myofibrillar proteins and structure of myofibrils. Food Chem. 2011, 128, 22–27. [Google Scholar] [CrossRef]
- Ottenheijm, C.A.; Granzier, H.; Labeit, S. The sarcomeric protein nebulin: Another multifunctional giant in charge of muscle strength optimization. Front. Physiol. 2012, 3, 37. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Li, M.; Du, M.; Shen, Q.W.; Zhang, D. Dephosphorylation enhances postmortem degradation of myofibrillar proteins. Food Chem. 2017, 245, 233–239. [Google Scholar] [CrossRef]
- Liu, P.; Zhang, Z.; Guo, X.; Zhu, X.; Mao, X.; Guo, X.; Deng, X.; Zhang, J. μ-Calpain oxidation and proteolytic changes on myofibrillar proteins from Coregonus Peled in vitro. Food Chem. 2021, 361, 130100. [Google Scholar] [CrossRef]
- Malheiros, J.M.; Braga, C.P.; Grove, R.A.; Ribeiro, F.A.; Calkins, C.R.; Adamec, J.; Chardulo, L.A.L. Influence of oxidative damage to proteins on meat tenderness using a proteomics approach. Meat Sci. 2019, 148, 64–71. [Google Scholar] [CrossRef]
- Chen, Q.; Huang, J.; Huang, F.; Huang, M.; Zhou, G. Influence of oxidation on the susceptibility of purified desmin to degradation by μ-calpain, caspase-3 and -6. Food Chem. 2014, 150, 220–226. [Google Scholar] [CrossRef]
- Ding, Z.; Wei, Q.; Zhang, C.; Zhang, H.; Huang, F. Influence of oxidation on heat shock protein 27 translocation, caspase-3 and calpain activities and myofibrils degradation in postmortem beef muscles. Food Chem. 2021, 340, 127914. [Google Scholar] [CrossRef]
- Ding, Z.; Huang, F.; Zhang, C.; Zhang, L.; Sun, H.; Zhang, H. Effect of heat shock protein 27 on the in vitro degradation of myofibrils by caspase-3 and μ-calpain. Int. J. Food Sci. Technol. 2018, 53, 121–128. [Google Scholar] [CrossRef]
- Musumeci, G.; Loreto, C.; Carnazza, M.L.; Martinez, G. Characterization of apoptosis in articular cartilage derived from the knee joints of patients with osteoarthritis. Knee Surg. Sports Traumatol. Arthrosc. 2011, 19, 307–313. [Google Scholar] [CrossRef]
- Zhang, J.; Yu, Q.; Han, L.; Han, M.; Han, G. Effects of lysosomal iron involvement in the mechanism of mitochondrial apoptosis on postmortem muscle protein degradation. Food Chem. 2020, 328, 127174. [Google Scholar] [CrossRef]
- Zhang, X.; Pan, D.; Cao, J.; Wu, Z. Changes in the major caspases involved in cytoskeletal degradation of goose muscle during prolonged aging. Food Res. Int. 2013, 51, 603–610. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Liu, P.; Zhao, Y.; Zhang, L.; Zhang, J. Oxidative Stress Contributes to Cytoskeletal Protein Degradation of Esox lucius through Activation of Mitochondrial Apoptosis during Postmortem Storage. Foods 2022, 11, 1308. https://doi.org/10.3390/foods11091308
Li X, Liu P, Zhao Y, Zhang L, Zhang J. Oxidative Stress Contributes to Cytoskeletal Protein Degradation of Esox lucius through Activation of Mitochondrial Apoptosis during Postmortem Storage. Foods. 2022; 11(9):1308. https://doi.org/10.3390/foods11091308
Chicago/Turabian StyleLi, Xue, Pingping Liu, Yunfeng Zhao, Lianfu Zhang, and Jian Zhang. 2022. "Oxidative Stress Contributes to Cytoskeletal Protein Degradation of Esox lucius through Activation of Mitochondrial Apoptosis during Postmortem Storage" Foods 11, no. 9: 1308. https://doi.org/10.3390/foods11091308
APA StyleLi, X., Liu, P., Zhao, Y., Zhang, L., & Zhang, J. (2022). Oxidative Stress Contributes to Cytoskeletal Protein Degradation of Esox lucius through Activation of Mitochondrial Apoptosis during Postmortem Storage. Foods, 11(9), 1308. https://doi.org/10.3390/foods11091308