
In Vitro Study of Probiotic, Antioxidant and Anti-Inflammatory Activities among Indigenous Saccharomyces cerevisiae Strains

 ,
,  ,
, 

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Origin of Yeast Strains
2.2. Preliminary Characterization
2.2.1. Growth at Human Body Temperature
2.2.2. Strain Tolerance to Different pH
- GI < 50%: Yeast Inhibition
- 50% < GI < 75%: Partial Inhibition
- GI > 75%: Growth Similar to the Control
2.2.3. Hydrophobicity of the Cells Surface
2.2.4. Antibiotic Resistance
2.3. Screening for Potentially Probiotic Traits: Simulation of the Transit into Gut
- ▪ Gastric Juice: 0.26 g/L of pepsin (porcine gastric mucosal, Sigma, Milan, Italy) were dissolved in HCl 0.1 N and the final pH of the solution was adjusted to 1.9 with 1 N NaOH;
- ▪ Intestinal fluid: 1.95 g/L of pancreatin (porcine pancreas, Sigma, Milan, Italy) were dissolved in a phosphate buffer solution (NaH2PO4/Na2HPO4 0.02 M, pH 7.5), and added with 3 g/L of bile extract (bile extract porcine, Sigma, Milan, Italy), dissolved in sterile distilled water; and the final pH was adjusted to 7.5 with 1N NaOH.
2.4. Evaluation of Functional Activities on Four Selected Strains
2.4.1. Total Antioxidant Activity
2.4.2. Determination of Glucans
Cell Culture and Treatments
Cytotoxicity Assay
ROS and NO Detection
2.5. Data Analysis
3. Results and Discussion
3.1. Preliminary Characterization
- -
- Growth at human body temperature: The temperature is an important physical parameter to evaluate the probiotic aptitude of the microorganism. In this study, the experiments were carried out at 37 °C, the typical temperature of the human body, and all the strains were able to grow at 37 °C (data not shown). These results agree with previous findings, reporting the ability of yeast strains to grow at human body temperature [22], although yeasts usually show a trend toward surviving to lower temperatures than lactic acid bacteria [61], and confirm that indigenous yeast strains might be an interesting source for the selection of new potential probiotic strains.
- -
- Strain tolerance to different pH: As regards the tolerance to pH, different pH values (2.5, 3.0, 3.5, 4.0, and 7.2) were investigated in order to evaluate the ability of tested yeasts to survive at the pH of the gastro-intestinal environment. In fact, the probiotics must survive harsh conditions during their passage through the intestinal tract [11]. All the strains exhibited a growth similar to the control (GI > 75%) to pH 3.5, 4.2, and 7.2, whereas the lowest pH values (2.5 and 3.0) influenced the yeast growth. In particular, at pH 2.5, 15 strains were inhibited (GI < 50%), 31 strains (59.61%) showed a partial inhibition (50% < GI < 75%), whereas only 6 strains exhibited a good tolerance to low pH (GI > 75%). At pH 3.0, 13 strains exhibited a partial inhibition (50% < GI < 75%) and 39 strains (75%) grew similar to the control (GI > 75%) (Figure 1). The highest tolerance to low pH, with a GI > 90%, were found in five strains, namely FD-3, M5-15 (both isolated from sourdough), TA4-10, CA10-4sc2 (both isolated from grape must) and 9-15G, isolated from honey. Furthermore, the majority of indigenous strains analyzed in this step was more tolerant to low pH level than Sb, the commercial probiotic strain. Resistance to acidic conditions was not surprising since the strains studied in this work were isolated from low pH environments, such as grape must, or substrate in which they coexist with lactic acid bacteria, such as sourdough [62].
- -
- Hydrophobicity of the Cell Surface: Hydrophobicity was defined as an interaction between microbial and host cells, mediated by cell-surface proteins and lipoteichoic acids [63]. As a consequence, this is one of the main criteria for the selection of strains with potential probiotic activity, because the microbial strain included in the probiotic category must have the ability to adhere to the intestinal mucosa for colonization and modulation of the immune system against pathogens [64]. The hydrophobic ability of the yeasts was reported in Figure 1. The results highlighted that only two strains (BP2-33, and SA10-19) showed 0% MATS (no hydrophobic activity), while the majority of the strains (35) exhibited a MATS in a range between 50% to 75%, which were much higher than the level found in S. cerevisiae var. boulardii strain used as control, confirming other results reporting better hydrophobicity level in indigenous strains than in control strains [65,66].
- -
- Antibiotic Resistance: The fifty-two strains were tested for the resistance to five antibiotics, at five different doses (40, 80, 100, 130, and 150 µg/mL) and all the strains survived in the presence of the highest dose of the tested antibiotics (150 µg/mL); in fact, no inhibition zone was observed around the spotted strains (data not shown). The high resistance of the S. cerevisiae strains to these antibiotics provide them advantage over bacteria in patients undergoing antibiotic treatment for therapeutic use [67]. Furthermore, this characteristic is of great interest also for potential application of these strains as probiotic sources in animal feed in consequence of potential presence of antibiotics residues in animal feed.
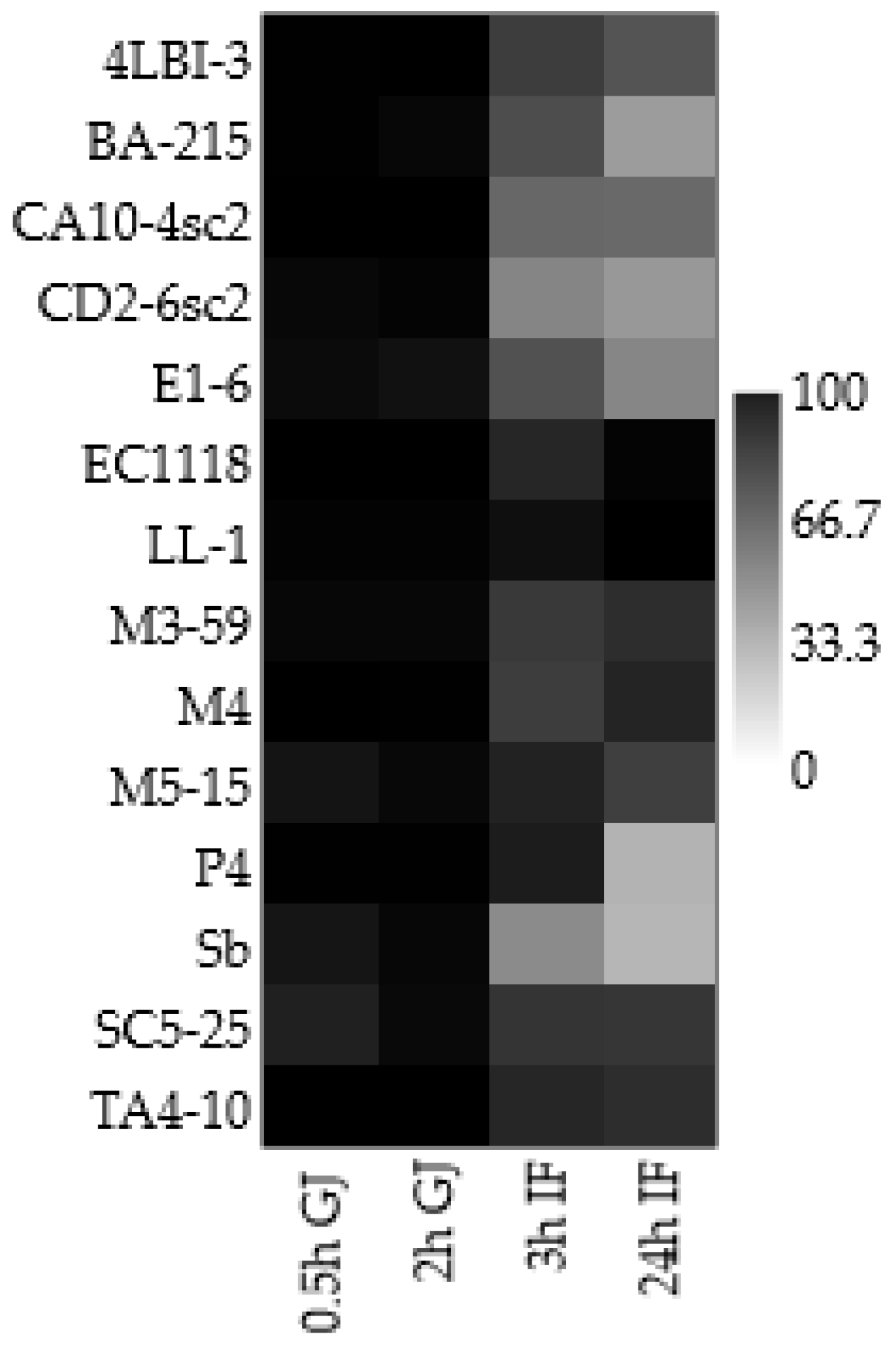
3.2. Screening for Potentially Probiotic Traits: Simulation of the Transit into Gut
3.3. Evaluation of Functional Characteristics on Four Selected Strains
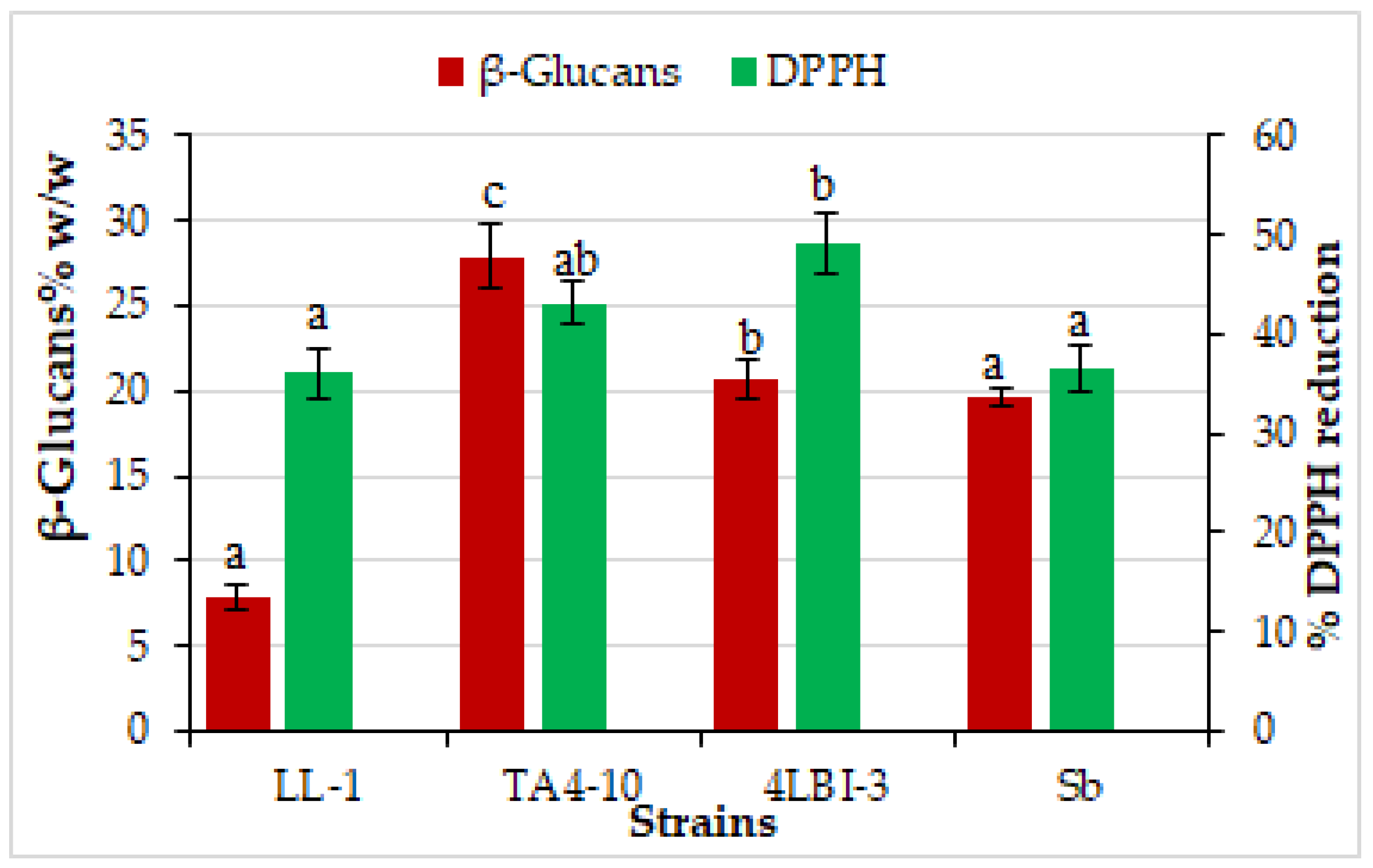
3.3.1. Antioxidant Activity and Glucans Content
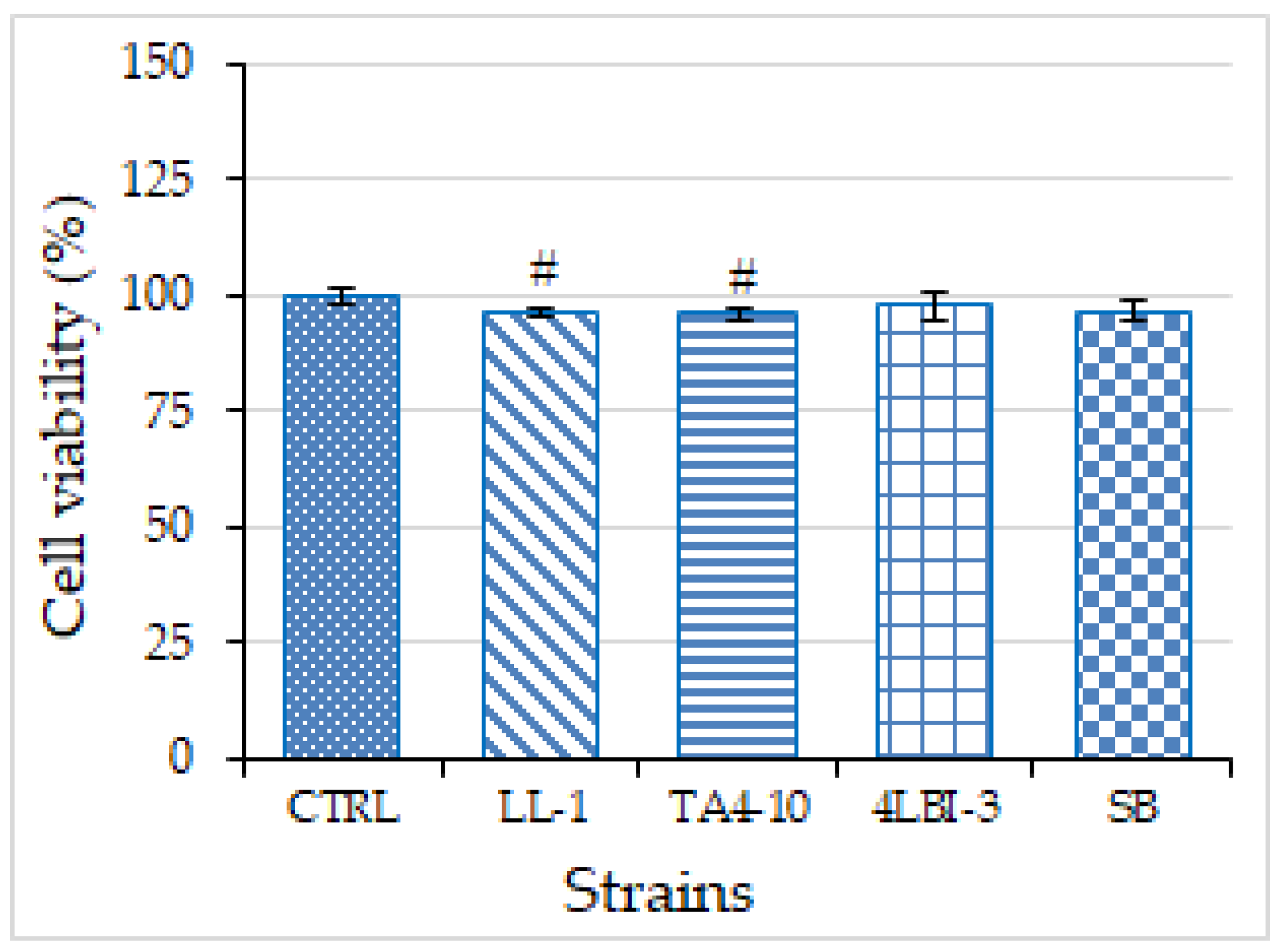
3.3.2. Cytotoxicity Assay and Anti-Inflammatory Activity
3.3.3. ROS and NO Detection
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document: The international scientific association for probiotics and prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markowiak, P.; Śliżewska, K. Effects of Probiotics, Prebiotics, and Synbiotics on Human Health. Nutrients 2017, 9, 1021. [Google Scholar] [CrossRef] [PubMed]
- Mallappa, R.H.; Singh, D.K.; Rokana, N.; Pradhan, D.; Batish, V.K.; Grover, S. Screening and selection of probiotic Lactobacillus strains of Indian gut origin based on assessment of desired probiotic attributes combined with principal component and heatmap analysis. LWT-Food Sci. Technol. 2019, 105, 272–281. [Google Scholar] [CrossRef]
- Katarzyna, D.; Magdalena, H.-M.; Marcin, S.; Jakub, S.; Tomasz, B. Probiotics in the mechanism of protection against gut inflammation and therapy of gastrointestinal disorders. Curr. Pharm. Des. 2014, 20, 1149–1155. [Google Scholar]
- Rokana, N.; Mallappa, R.H.; Batish, V.K.; Grover, S. Interaction between putative probiotic Lactobacillus strains of Indian gut origin and Salmonella: Impact on intestinal barrier function. LWT-Food Sci. Technol. 2016, 84, 851–860. [Google Scholar] [CrossRef]
- Kerry, R.G.; Patra, J.K.; Gouda, S.; Park, Y.; Shin, H.-S.; Das, G. Benefaction of probiotics for human health: A review. J. Food Drug. Anal. 2018, 26, 927–939. [Google Scholar] [CrossRef] [Green Version]
- Bevilacqua, A.; Campaniello, D.; Speranza, B.; Sinigaglia, M.; Corbo, M.R. Selection of promising probiotic strains for foods: Proposal for a possible flowchart with a special focus on the dark side of probiotics. In Probiotics in Health and Disease. New Research; Roma, A., Ed.; Nova Publisher: New York, NY, USA, 2014; p. 122. [Google Scholar]
- Almada, C.N.; Martinez, R.C.; Sant’Ana, A.S. Characterization of the intestinal microbiota and its interaction with probiotics and health impacts. Appl. Microbiol. Biotechnol. 2015, 99, 4175–4199. [Google Scholar] [CrossRef]
- Rodríguez, P.F.-P.; Arévalo-Villena, M.; Rosa, I.Z.; Pérez, A.B. Selection of potential non-Sacharomyces probiotic yeasts from food origin by a step-by-step approach. Food Res. Int. 2018, 112, 143–151. [Google Scholar] [CrossRef] [Green Version]
- McFarland, L.V. From yaks to yogurt: The history, development, and current use of probiotics. Clin. Infect. Dis. 2015, 60, S85–S90. [Google Scholar] [CrossRef] [Green Version]
- Rivera-Espinoza, Y.; Gallardo-Navarro, Y. Non-dairy probiotic products. Food Microbiol. 2010, 27, 1–11. [Google Scholar] [CrossRef]
- Fijan, S. Microorganisms with claimed probiotic properties: An overview of recent literature. Int. J. Environ. Res. Public Health 2014, 11, 4745–4767. [Google Scholar] [CrossRef] [PubMed]
- Vrese, M.; Schrezenmeir, J. Probiotics, prebiotics, and synbiotics. Adv. Biochem. Eng. Biotechnol. 2008, 111, 1–66. [Google Scholar] [PubMed]
- Chen, L.S.; Ma, Y.; Maubois, J.L.; Chen, L.J.; Liu, Q.H.; Guo, J.P. Identification of yeasts from raw milk and selection for some specific antioxidant properties. Int. J. Dairy Technol. 2010, 63, 47–54. [Google Scholar] [CrossRef]
- Czerucka, D.; Rampal, P. Experimental effects of Saccharomyces boulardii on diarrheal pathogens. Microbes Infect. 2002, 4, 733–739. [Google Scholar] [CrossRef]
- Hatoum, R.; Labrie, S.; Fliss, I. Antimicrobial and probiotic properties of yeasts: From fundamental to novel applications. Front. Microbiol. 2012, 3, 421. [Google Scholar] [CrossRef] [Green Version]
- Arroyo-López, F.N.; Romero-Gil, V.; Bautista-Gallego, J.; Rodríguez-Gómez, F.; Jiménez-Díaz, R.; García-García, P.; Querol, A.; Garrido-Fernández, A. Potential benefits of the application of yeast starters in table olive processing. Front. Microbiol. 2012, 5, 34. [Google Scholar] [CrossRef] [Green Version]
- Moreira, N.; Pina, C.; Mendes, F.; Couto, J.A.; Hogg, T.; Vasconcelos, I. Volatile compounds contribution of Hanseniaspora guilliermondii and Hanseniaspora uvarum during red wine vinifications. Food Control. 2011, 22, 662–667. [Google Scholar] [CrossRef]
- Moslehi-Jenabian, S.; Lindegaard, L.; Jespersen, L. Beneficial effects of probiotic and food borne yeasts on human health. Nutrients 2010, 2, 449–473. [Google Scholar] [CrossRef]
- Arévalo-Villena, M.; Fernández-Pacheco, P.; Castillo, N.; Bevilacqua, A.; Briones, A. Probiotic capability in Saccharomyces yeasts: Set up of a screening method. LWT-Food Sci. Technol. 2018, 89, 657–665. [Google Scholar] [CrossRef]
- Diosma, G.; Romanin, D.E.; Rey-Burusco, M.F.; Londero, A.; Garrote, G.L. Yeasts from kefir grains: Isolation, identification and probiotic characterization. World J. Microbiol. Biotechnol. 2014, 30, 43–53. [Google Scholar] [CrossRef]
- Gil-Rodríguez, A.M.; Carrascosa, A.V.; Requena, T. Yeasts in food and beverages: In vitro characterization of probiotic traits. LWT-Food Sci. Technol. 2015, 64, 1156–1162. [Google Scholar] [CrossRef] [Green Version]
- Perricone, M.; Bevilacqua, A.; Corbo, M.R.; Sinigaglia, M. Technological characterization and probiotic traits of yeasts isolated from Altamura sourdough to select promising microorganisms as functional starter cultures for cereal-based products. Food Microbiol. 2014, 38, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Silva, T.; Reto, M.; Sol, M.; Peito, A.; Peres, C.M.; Peres, C.; Malcata, F.X. Characterization of yeasts from Portuguese brined olives, with a focus on their potentially probiotic behavior. LWT-Food Sci. Technol. 2011, 44, 1349–1354. [Google Scholar] [CrossRef]
- Xu, C.; Ma, J.; Wang, W.; Liu, Z.; Gu, L.; Qian, S.; Hou, J.; Jiang, Z. Preparation of pectin-based nanofibers encapsulating Lactobacillus rhamnosus 1.0320 by electrospinning. Food Hydrocoll. 2022, 124, 107216. [Google Scholar] [CrossRef]
- Cordonnier, C.; Thévenot, J.; Etienne-Mesmin, L.; Denis, S.; Alric, M.; Livrelli, V.; Blanquet-Diot, S. Dynamic In Vitro Models of the Human Gastrointestinal Tract as Relevant Tools to Assess the Survival of Probiotic Strains and Their Interactions with Gut Microbiota. Microorganisms 2015, 3, 725–745. [Google Scholar] [CrossRef]
- Mu, Z.; Yang, Y.; Xia, Y.; Wang, F.; Sun, Y.; Yang, Y.; Ai, L. Probiotic yeast BR14 ameliorates DSS-induced colitis by restoring the gut barrier and adjusting the intestinal microbiota. Food Funct. 2021, 12, 8386–8398. [Google Scholar] [CrossRef]
- Pérez-Sotelo, L.S.; Talavera-Rojas, M.; Monroy-Salazar, H.G.; Lagunas-Bernabé, S.; Cuarón-Ibargüengoytia, J.A.; Jiménez, R.M. In vitro evaluation of the binding capacity of Saccharomyces cerevisiae Sc47 to adhere to the wall of Salmonella spp. Rev. Latinoam. Microbiol. 2005, 47, 70–75. [Google Scholar]
- Fooks, L.J.; Gibson, G.R. Probiotics as modulators of the gut flora. Br. J. Nutr. 2002, 88, 39–49. [Google Scholar] [CrossRef] [Green Version]
- Lokesh, J.; Fernandes, J.; Korsnes, K.; Bergh, O.; Brinchmann, M.; Kiron, V. Transcriptional regulation of cytokines in the intestine of Atlantic cod fed yeast derived mannan oligosaccharide or beta-Glucan and challenged with Vibrio anguillarum. Fish Shellfish Immunol. 2012, 33, 626–631. [Google Scholar] [CrossRef]
- Anderson, J.W. Dietary fibre, complex carbohydrate and coronary artery disease. Can. J. Cardiol. 1995, 11, 55G–62G. [Google Scholar]
- Pais, P.; Almeida, V.; Yılmaz, M.; Teixeira, M.C. Saccharomyces boulardii: What Makes It Tick as Successful Probiotic? J. Fungi 2020, 6, 78. [Google Scholar] [CrossRef] [PubMed]
- Martins, F.S.; Dalmasso, G.; Arantes, R.M.E.; Doye, A.; Lemichez, E.; Lagadec, P.; Imbert, V.; Peyron, J.-F.; Rampal, P.; Nicoli, J.R.; et al. Interaction of Saccharomyces boulardii with Salmonella enterica Serovar Typhimurium Protects Mice and Modifies T84 Cell Response to the Infection. PLoS ONE 2010, 5, e8925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santarsiero, A.; Convertini, P.; Todisco, S.; Pierri, C.L.; De Grassi, A.; Williams, N.C.; Iacobazzi, D.; De Stefano, G.; O’Neill, L.A.J.; Infantino, V. ACLY Nuclear Translocation in Human Macrophages Drives Proinflammatory Gene Expression by NF-κB Acetylation. Cells 2021, 10, 2962. [Google Scholar] [CrossRef] [PubMed]
- Infantino, V.; Pierri Leonardo, C.; Iacobazzi, V. Metabolic Routes in Inflammation: The Citrate Pathway and its Potential as Therapeutic Target. Curr. Med. Chem. 2019, 26, 7104–7116. [Google Scholar] [CrossRef] [PubMed]
- Menga, A.; Serra, M.; Todisco, S.; Riera-Domingo, C.; Ammarah, U.; Ehling, M.; Palmieri, E.M.; Di Noia, M.A.; Gissi, R.; Favia, M.; et al. Glufosinate constrains synchronous and metachronous metastasis by promoting anti-tumor macrophages. EMBO Mol. Med. 2020, 12, 11210. [Google Scholar] [CrossRef] [PubMed]
- Todisco, S.; Convertini, P.; Iacobazzi, V.; Infantino, V. TCA Cycle Rewiring as Emerging Metabolic Signature of Hepatocellular Carcinoma. Cancers 2020, 12, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santarsiero, A.; Bochicchio, A.; Funicello, M.; Lupattelli, P.; Choppin, S.; Colobert, F.; Hanquet, G.; Schiavo, L.; Convertini, P.; Chiummiento, L.; et al. New synthesized polyoxygenated diarylheptanoids suppress lipopolysaccharide-induced neuroinflammation. Biochem. Biophys. Res. Commun. 2020, 529, 1117–1123. [Google Scholar] [CrossRef]
- Infantino, V.; Iacobazzi, V.; Menga, A.; Avantaggiati, M.L.; Palmieri, F. A key role of the mitochondrial citrate carrier (SLC25A1) in TNFα- and IFNγ-triggered inflammation. Biochim. Biophys. Acta 2014, 1839, 1217–1225. [Google Scholar] [CrossRef] [Green Version]
- Santarsiero, A.; Convertini, P.; Vassallo, A.; Santoro, V.; Todisco, S.; Iacobazzi, D.; Fondufe-Mittendorf, Y.; Martelli, G.; de Oliveira, M.R.; Montanaro, R.; et al. Phenolic Compounds of Red Wine Aglianico del Vulture Modulate the Functional Activity of Macrophages via Inhibition of NF-κB and the Citrate Pathway. Oxid. Med. Cell. Longev. 2021, 793. [Google Scholar] [CrossRef]
- Capece, A.; Pietrafesa, R.; Romano, P. Experimental approach for target selection of wild wine yeasts from spontaneous fermentation of ‘‘Inzolia’’ grapes. World J. Microbiol. Biotechnol. 2011, 27, 2775–2783. [Google Scholar] [CrossRef]
- Capece, A.; Romaniello, R.; Siesto, G.; Pietrafesa, R.; Massari, C.; Poeta, C.; Romano, P. Selection of indigenous Saccharomyces cerevisiae strains for Nero d’Avola wine and evaluation of selected starter implantation in pilot fementation. Int. J. Food Microbiol. 2010, 144, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Capece, A.; Siesto, G.; Poeta, C.; Pietrafesa, R.; Romano, P. Indigenous yeast population from Georgian aged wines produced by traditional “Kakhetian” method. Food Microbiol. 2013, 36, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Capece, A.; Siesto, G.; Romaniello, R.; Lagreca, V.M.; Pietrafesa, R.; Calabretti, A.; Romano, P. Assessment of competition in wine fermentation among wild Saccharomyces cerevisiae strains isolated from Sangiovese grapes in Tuscany region. LWT-Food Sci. Technol. 2013, 54, 485–492. [Google Scholar] [CrossRef]
- Siesto, G.; Capece, A.; Sipiczki, M.; Csoma, H.; Romano, P. Polymorphism detection among wild Saccharomyces cerevisiae strains of different wine origin. Ann. Microbiol. 2013, 63, 661–668. [Google Scholar] [CrossRef]
- Brandolini, V.; Tedeschi, P.; Capece, A.; Maietti, A.; Mazzotta, D.; Salzano, G.; Paparella, A.; Romano, P. Saccharomyces cerevisiae wine strains differing in copper resistance exhibit different capability to reduce copper content in wine. World J. Microbiol. Biotechnol. 2002, 18, 499–503. [Google Scholar] [CrossRef]
- Capece, A.; Pietrafesa, R.; Siesto, G.; Romaniello, R.; Condelli, N.; Romano, P. Selected Indigenous Saccharomyces cerevisiae Strains as Profitable Strategy to Preserve Typical Traits of Primitivo Wine. Fermentation 2019, 5, 87. [Google Scholar] [CrossRef] [Green Version]
- Capece, A.; Romaniello, R.; Pietrafesa, R.; Romano, P. Indigenous Saccharomyces cerevisiae yeasts as a source of biodiversity for the selection of starters for specific fermentations. BIO Web Conf. 2014, 3, 02003. [Google Scholar] [CrossRef] [Green Version]
- Capece, A.; Romaniello, R.; Pietrafesa, A.; Siesto, G.; Pietrafesa, R.; Zambuto, M.; Romano, P. Use of Saccharomyces cerevisiae var. boulardii in co-fermentations with S. cerevisiae for the production of craft beers with potential healthy value added. Int. J. Food Microbiol. 2018, 284, 22–30. [Google Scholar] [CrossRef]
- Capece, A.; Romaniello, R.; Siesto, G.; Romano, P. Diversity of Saccharomyces cerevisiae yeasts associated to spontaneously fermenting grapes from an Italian “heroic vine-growing area”. Food Microbiol. 2012, 31, 159–166. [Google Scholar] [CrossRef]
- Bevilacqua, A.; Perricone, M.; Cannarsi, M.; Corbo, M.R.; Sinigaglia, M. Technological and spoiling characteristics of the yeast microflora isolated from Bella Di Cerignola table olives. Int. J. Food Sci. 2009, 44, 2198–2207. [Google Scholar] [CrossRef]
- Chelliah, R.; Ramakrishnan, S.R.; Prabhu, P.R.; Antony, U. Evaluation of antimicrobial activity and probiotic properties of wild-strain Pichia kudriavzevii isolated from frozen idli batter. Yeast 2016, 33, 385–401. [Google Scholar] [CrossRef] [PubMed]
- NCCLS (National Committee for Clinical Laboratory Standards) Performance Standard for Antimicrobial Disc Susceptibility Tests; Approved Standard. National Committee for Clinical Laboratory Standards: Villanova, PA, USA, 1993; Publication M2-A5; Available online: https://archive.org/details/performancestand00nati (accessed on 13 July 2021).
- Picot, A.; Lacroix, C. Encapsulation of bifidobacteria in whey protein-based microcapsules and survival in simulated gastrointestinal conditions and in yoghurt. Int. Dairy J. 2004, 14, 505–515. [Google Scholar] [CrossRef]
- McCleary, B.V.; Draga, A. Measurement of β-Glucan in Mushrooms and Mycelial Products. J. AOAC Int. 2016, 99, 364–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laurita, T.; Pappalardo, I.; Chiummiento, L.; D’Orsi, R.; Funicello, M.; Santarsiero, A.; Marsico, M.; Infantino, V.; Todisco, S.; Lupattelli, P. Synthesis of new methoxy derivatives of trans 2,3-diaryl-2,3-dihydrobenzofurans and evaluation of their anti-inflammatory activity. Bioorg. Med. Chem. Lett. 2021, 49, 128264. [Google Scholar] [CrossRef] [PubMed]
- Chimento, A.; Santarsiero, A.; Iacopetta, D.; Ceramella, J.; De Luca, A.; Infantino, V.; Parisi, O.I.; Avena, P.; Bonomo, M.G.; Saturnino, C.; et al. A Phenylacetamide Resveratrol Derivative Exerts Inhibitory Effects on Breast Cancer Cell Growth. Int. J. Mol. Sci. 2021, 22, 5255. [Google Scholar] [CrossRef] [PubMed]
- Ongena, K.; Das, C.; Smith, J.L.; Gil, S.; Johnston, G. Determining cell number during cell culture using the Scepter cell counter. J. Vis. Exp. 2010, 45, 2204. [Google Scholar] [CrossRef]
- Santarsiero, A.; Onzo, A.; Pascale, R.; Acquavia, M.A.; Coviello, M.; Convertini, P.; Todisco, S.; Marsico, M.; Pifano, C.; Iannece, P.; et al. Pistacia lentiscus Hydrosol: Untargeted Metabolomic Analysis and Anti-Inflammatory Activity Mediated by NF-κB and the Citrate Pathway. Oxid. Med. Cell. Longev. 2020. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Romero-Gil, V.; Bautista-Gallego, J.; Rodríguez-Gómez, F.; García-García, P.; Jimenez-Díaz, R.; Garrido-Fernández, A.; Arroyo-López, F.N. Evaluating the individual effects of temperature and salt on table olive related microorganisms. Food Microbiol. 2013, 33, 178–184. [Google Scholar] [CrossRef]
- Menezes, A.G.T.; Ramos, C.L.; Cenzi, G.; Melo, D.S.; Dias, D.R.; Schwan, R.F. Probiotic Potential, Antioxidant Activity, and Phytase Production of Indigenous Yeasts Isolated from Indigenous Fermented Foods. Probiotics Antimicrob. Proteins 2020, 12, 280–288. [Google Scholar] [CrossRef]
- Todorov, S.; Botes, M.; Guigas, C.; Schillinger, U.; Wiid, I.; Wachsman, M.; Dicks, L. Boza, a natural source of probiotic lactic acid bacteria. J. Appl. Microbiol. 2008, 104, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Ailioaie, L.M.; Litscher, G. Probiotics, Photobiomodulation, and Disease Management: Controversies and Challenges. Int. J. Mol. Sci. 2021, 22, 4942. [Google Scholar] [CrossRef] [PubMed]
- Fadda, M.E.; Mossa, V.; Deplano, M.; Pisano, M.B.; Cosentino, S. In vitro screening of Kluyveromyces strains isolated from Fiore Sardo cheese for potential use as probiotics. LWT-Food Sci. Technol. 2017, 75, 100–106. [Google Scholar] [CrossRef]
- Gut, A.M.; Vasiljevic, T.; Yeager, T.; Donkor, O.N. Characterization of yeasts isolated from traditional kefir grains for potential probiotic properties. J. Funct. Foods 2019, 58, 56–66. [Google Scholar] [CrossRef]
- Syal, P.; Vohra, A. Probiotic attributes of a yeast-like fungus, Geotrichum klebahnii. Afr. J. Microbiol. Res. 2014, 8, 2037–2043. [Google Scholar]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food e an international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [Green Version]
- Binetti, A.; Carrasco, M.; Reinheimer, J.; Suárez, V. Yeasts from autochthonal cheese starters: Technological and functional properties. J. Appl. Microbiol. 2013, 115, 434–444. [Google Scholar] [CrossRef]
- Merchán, A.V.; Benito, M.J.; Galván, A.I.; Ruiz-Moyano, S. Identification and selection of yeast with functional properties for future application in soft paste cheese. LWT-Food Sci. Technol. 2020, 124, 109173. [Google Scholar] [CrossRef]
- Ogunremi, O.R.; Agrawal, R.; Sanni, A.I. Development of cereal-based functional food using cereal-mix substrate fermented with probiotic strain—Pichia kudriavzevii OG32. Food Sci. Nutr. 2015, 3, 486–494. [Google Scholar] [CrossRef] [Green Version]
- Abbas, C.A. Production of antioxidants, aromas, colours, flavours, and vitamins by yeasts. In Yeasts in Food and Beverages; Querol, A., Fleet, G., Eds.; Springer: Berlin, Heidelberg, 2006; pp. 285–334. [Google Scholar]
- Jaehrig, S.C.; Rohn, S.; Kroh, L.W.; Fleischer, L.G.; Kurz, T. In vitro potential antioxidant activity of (1→3),(1→6)-beta-D-glucan and protein fractions from Saccharomyces cerevisiae cell walls. J. Agric. Food Chem. 2007, 55, 4710–4716. [Google Scholar] [CrossRef]
- Hong, J.Y.; Son, S.H.; Hong, S.P.; Yi, S.H.; Kang, S.H.; Lee, N.K.; Paik, H.D. Production of β-glucan, glutathione, and glutathione derivatives by probiotic Saccharomyces cerevisiae isolated from cucumber jangajji. LWT-Food Sci. Technol. 2019, 100, 114–118. [Google Scholar] [CrossRef]
- Boutros, J.A.; Magee, A.S.; Cox, D. Comparison of structural differences between yeast β-glucan sourced from different strains of Saccharomyces cerevisiae and processed using proprietary manufacturing processes. Food Chem. 2022, 367, 130708. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.H.; Kim, H.R.; Kim, J.H.; Ahn, B.H.; Kim, T.W.; Lee, J.E. Identification of wild yeast strains and analysis of their β-glucan and glutathione levels for use in makgeolli brewing. Microbiology 2014, 42, 361–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, N.K.; Hong, J.Y.; Yi, S.H.; Hong, S.P.; Lee, J.E.; Paik, H.D. Bioactive compounds of probiotic Saccharomyces cerevisiae strains isolated from cucumber jangajji. J. Funct. Foods 2019, 58, 324–329. [Google Scholar] [CrossRef]
- Goodridge, H.S.; Wolf, A.J.; Underhill, D.M. Beta-glucan recognition by the innate immune system. Immunol. Rev. 2009, 230, 38–50. [Google Scholar] [CrossRef]
- Xu, X.; Yasuda, M.; Mizuno, M.; Ashida, H. β-Glucan from Saccharomyces cerevisiae reduces lipopolysaccharide-induced inflammatory responses in RAW264.7 macrophages. Biochim. Biophys. Acta 2012, 1820, 1656–1663. [Google Scholar] [CrossRef]
- Catania, A.; Cutuli, M.; Garofalo, L.; Carlin, A.; Airaghi, L.; Barcellini, W.; Lipton, J.M. The neuropeptide alpha-MSH in host defense. In Neuroimmunomodulation: Perspectives at the New Millennium; Conti, A., Maestroni, G.J.M., McCann, S.M., Sternberg, E.M., Lipton, J.M., Smith, C.C., Eds.; New York Academy of Sciences: New York, NY, USA, 2000; Volume 917, pp. 227–231. [Google Scholar]
- Williams, R.; Dias, D.A.; Jayasinghe, N.; Roessner, U.; Bennett, L.E. Beta-glucan-depleted, glycopeptide-rich extracts from Brewer’s and Baker’s yeast (Saccharomyces cerevisiae) lower interferon-gamma production by stimulated human blood cells in vitro. Food Chem. 2016, 197, 761–768. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Code | Origin | Reference |
|---|---|---|
| AC3-1; TA4-10 | Inzolia Grape Must, Sicily (Italy) | [41] |
| CA10-4sc2; CB1-3sv2; CB2-8sr3; CD2-6sc2; CD7-3sv2; TA8-4sc2; RA2-1sb2; RA8-1sv2; RE8-7sr3 | Nero d’Avola Grape Must, Sicily (Italy) | [42] |
| ANF1-9; ANF8-3; ANFA-12; ANFB-11; ANFC-3; ANFC-30; ANFD-18; ANFD-21; GEO2-8 | Kaketian Wine, (Georgia) | [43] |
| SA7-13; SA10-19; SB5-18; SC5-25; SC5-37 | Sangiovese Grape Must, Tuscany (Italy) | [44] |
| BA-215; SN-41 | Sangiovese Grape Must, Emilia Romagna (Italy) | [45,46] |
| BP2-33; Mpr2-18 | Primitivo Grape Must, Basilicata (Italy) | [47] |
| E1-6; FIMA-3; M1-47; M1-110; M3-59; M3-60; M3-80; VEME-2; 4LBI-3 | Aglianico Grape Must, Basilicata (Italy) | [45,48] |
| FD-3; LL-1; M5-15; P4 | Sourdough, Basilicata (Italy) | [49] |
| M4 | Beer, Basilicata (Italy) | [49] |
| 5TB8-20; 5TB8-28; 5TB8-60; 5TB8-73 | Bosco Grape Must, Liguria (Italy) | [50] |
| 9-15b; 9-15G; 12-15b | Honey, (Malta) | [49] |
| Strains | GI (%) pH2.5 | GI (%) pH3.0 | MATS (%) |
|---|---|---|---|
| 4LBI-3 | 50–75 | >75 | 91.98 ± 10.25 |
| BA-215 | 50–75 | 50–75 | 55.95 ± 5.80 |
| CA10-4sc2 | >75 | >75 | 51.18 ± 4.99 |
| CD2-6sc2 | 50–75 | >75 | 52.51 ± 5.60 |
| E1-6 | 50–75 | >75 | 74.53 ± 7.50 |
| EC1118 | 50–75 | >75 | 10.19 ± 1.05 |
| LL-1 | 50–75 | >75 | 98.79 ± 9.60 |
| M3-59 | <50 | >75 | 58.17 ± 4.84 |
| M4 | 50–75 | >75 | 57.98 ± 5.75 |
| M5-15 | >75 | >75 | 78.50 ± 6.88 |
| P4 | 50–75 | >75 | 92.37 ± 8.98 |
| Sb | <50 | >75 | 36.48 ± 3.55 |
| SC5-25 | 50–75 | >75 | 59.30 ± 5.13 |
| TA4-10 | >75 | >75 | 96.77 ± 9.42 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siesto, G.; Pietrafesa, R.; Infantino, V.; Thanh, C.; Pappalardo, I.; Romano, P.; Capece, A. In Vitro Study of Probiotic, Antioxidant and Anti-Inflammatory Activities among Indigenous Saccharomyces cerevisiae Strains. Foods 2022, 11, 1342. https://doi.org/10.3390/foods11091342
Siesto G, Pietrafesa R, Infantino V, Thanh C, Pappalardo I, Romano P, Capece A. In Vitro Study of Probiotic, Antioxidant and Anti-Inflammatory Activities among Indigenous Saccharomyces cerevisiae Strains. Foods. 2022; 11(9):1342. https://doi.org/10.3390/foods11091342
Chicago/Turabian StyleSiesto, Gabriella, Rocchina Pietrafesa, Vittoria Infantino, Channmuny Thanh, Ilaria Pappalardo, Patrizia Romano, and Angela Capece. 2022. "In Vitro Study of Probiotic, Antioxidant and Anti-Inflammatory Activities among Indigenous Saccharomyces cerevisiae Strains" Foods 11, no. 9: 1342. https://doi.org/10.3390/foods11091342
APA StyleSiesto, G., Pietrafesa, R., Infantino, V., Thanh, C., Pappalardo, I., Romano, P., & Capece, A. (2022). In Vitro Study of Probiotic, Antioxidant and Anti-Inflammatory Activities among Indigenous Saccharomyces cerevisiae Strains. Foods, 11(9), 1342. https://doi.org/10.3390/foods11091342