
TMT-Based Proteomic Analysis of Hannaella sinensis-Induced Apple Resistance-Related Proteins



Abstract
:1. Introduction
2. Materials and Methods
2.1. H. sinensis
2.2. Apple
2.3. Protein Extraction from Apple Samples
2.4. Proteolysis and TMT Labeling
2.5. High pH Reverse-Phase Separation
2.6. Bioinformatics Analysis
3. Results
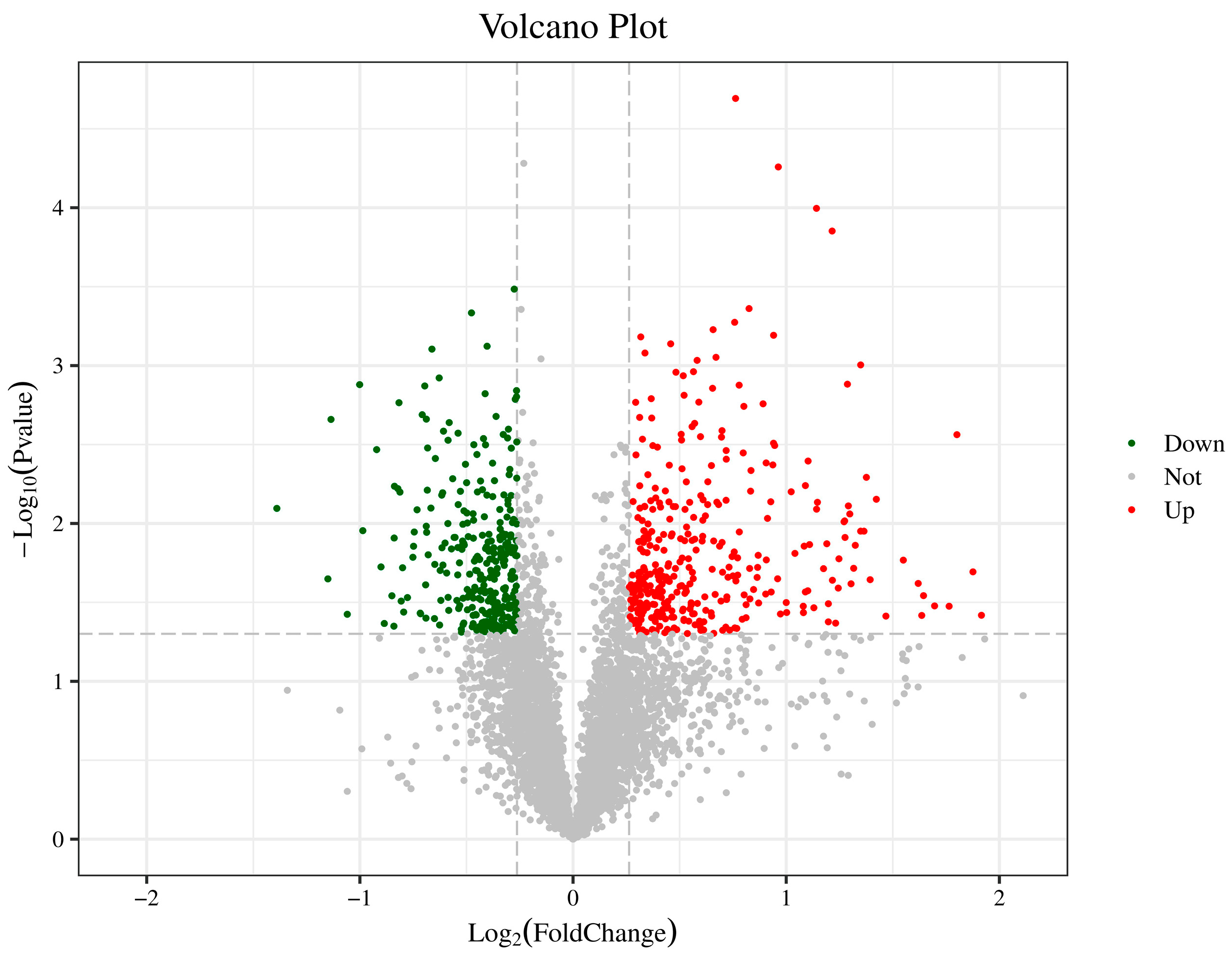
3.1. Protein Differential Analysis
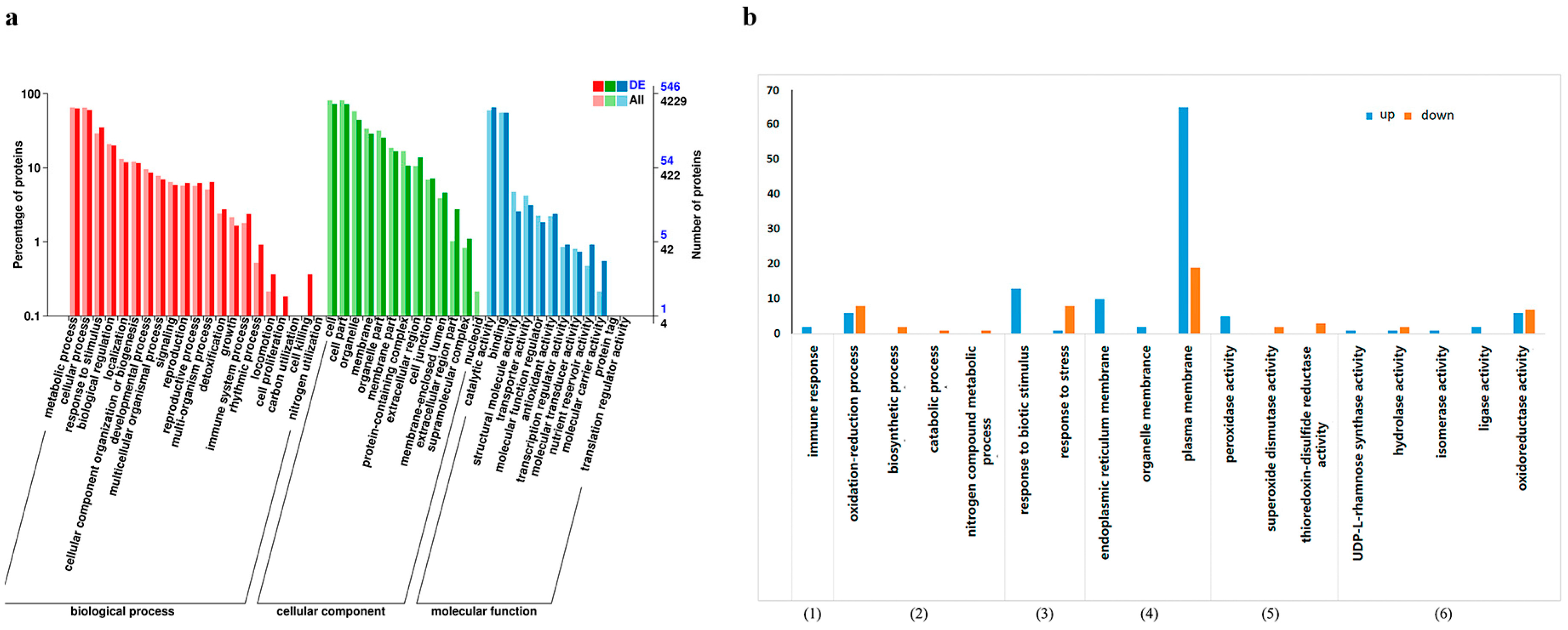
3.2. Differential Protein GO Enrichment Analysis
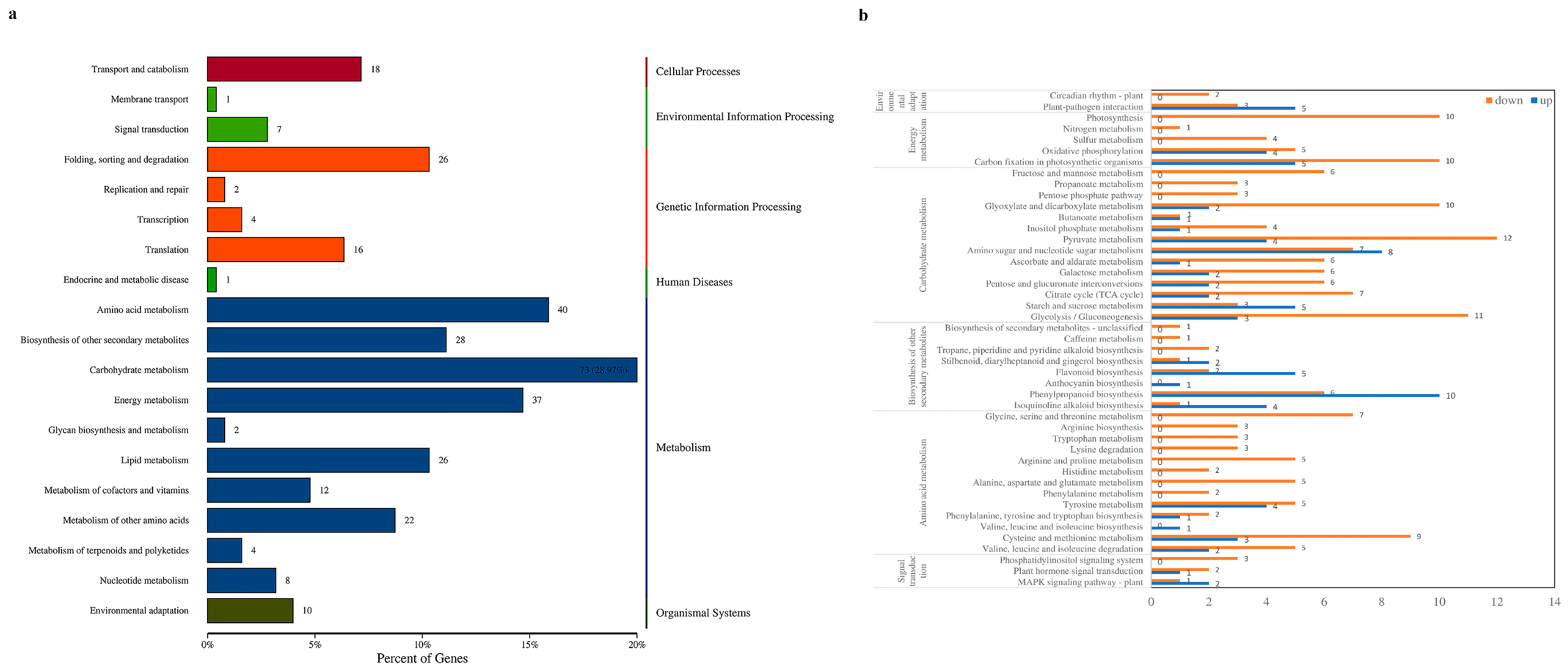
3.3. Differential Protein KEGG Enrichment Analysis
3.4. Differential Protein KEGG Enrichment Analysis
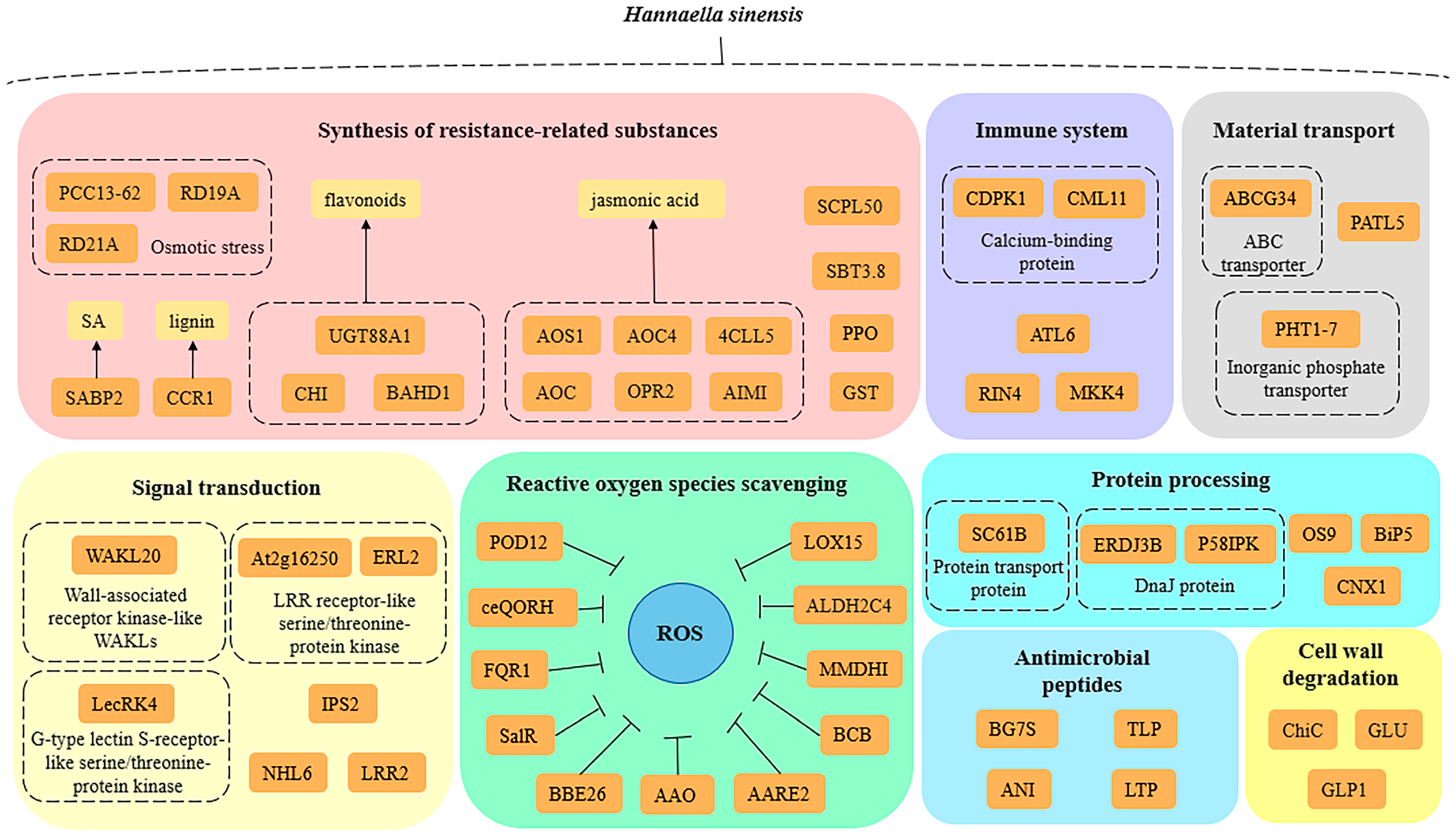
3.5. Functional Classification of Related DEPs
4. Discussion
4.1. Scavenging Reactive Oxygen Species
4.2. Improve the Synthesize of Resistance-Related Substances
4.3. Improve Plant Disease Resistance
4.4. Degrade Pathogen Cell Walls
4.5. Improve Plant Signal Transduction Ability
4.6. Antimicrobial-Related Activity
4.7. Substance-Related Transport
4.8. Protein Processing
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Janisiewicz, W.J.; Tworkoski, T.J.; Kurtzman, C.P. Biocontrol potential of Metchnikowia pulcherrima strains against blue mold of apple. Phytopathology 2001, 91, 1098–1108. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Ngea, G.L.N.; Godana, E.A.; Shi, Y.; Lanhuang, B.; Zhang, X.; Zhao, L.; Yang, Q.; Wang, S.; Zhang, H. Recent advances in Penicillium expansum infection mechanisms and current methods in controlling P. Expans in postharvest apples. Crit. Rev. Food Sci. Nutr. 2021, 63, 2598–2611. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Montiel, L.G.; Droby, S.; Preciado-Rangel, P.; Rivas-Garcia, T.; Gonzalez-Estrada, R.R.; Gutierrez-Martinez, P.; Avila-Quezada, G.D. A sustainable alternative for postharvest disease management and phytopathogens biocontrol in fruit: Antagonistic yeasts. Plants 2021, 10, 2641. [Google Scholar] [CrossRef]
- Dukare, A.S.; Paul, S.; Nambi, V.E.; Gupta, R.K.; Singh, R.; Sharma, K.; Vishwakarma, R.K. Exploitation of microbial antagonists for the control of postharvest diseases of fruits: A review. Crit. Rev. Food Sci. Nutr. 2019, 59, 1498–1513. [Google Scholar] [CrossRef]
- Liu, J.; Sui, Y.; Wisniewski, M.; Droby, S.; Liu, Y. Review: Utilization of antagonistic yeasts to manage postharvest fungal diseases of fruit. Int. J. Food Microbiol. 2013, 167, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Zaynab, M.; Fatima, M.; Abbas, S.; Sharif, Y.; Umair, M.; Zafar, M.H.; Bahadar, K. Role of secondary metabolites in plant defense against pathogens. Microb. Pathog. 2018, 124, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Das, J.; Yadav, S.K.; Ghosh, S.; Tyagi, K.; Magotra, A.; Krishnan, A.; Jha, G. Enzymatic and non-enzymatic functional attributes of plant microbiome. Curr. Opin. Biotechnol. 2021, 69, 162–171. [Google Scholar] [CrossRef]
- Ruan, J.; Zhou, Y.; Zhou, M.; Yan, J.; Khurshid, M.; Weng, W.; Cheng, J.; Zhang, K. Jasmonic acid signaling pathway in plants. Int. J. Mol. Sci. 2019, 20, 2479. [Google Scholar] [CrossRef] [Green Version]
- Kumar, D.; Klessig, D.F. High-affinity salicylic acid-binding protein 2 is required for plant innate immunity has salicylic acid-stimulated lipase activity. Proc. Natl. Acad. Sci. USA 2003, 100, 16101–16106. [Google Scholar]
- Watanabe, M.; Chiba, Y.; Hirai, M.Y. Metabolism and regulatory functions of O-acetylserine, S-adenosylmethionine, homocysteine, and serine in plant development and environmental responses. Front. Plant Sci. 2021, 12, 643403. [Google Scholar] [CrossRef]
- Farooq, M.A.; Niazi, A.K.; Akhtar, J.; Saifullah; Farooq, M.; Souri, Z.; Karimi, N.; Rengel, Z. Acquiring control: The evolution of ROS-Induced oxidative stress and redox signaling pathways in plant stress responses. Plant Physiol. Biochem. 2019, 141, 353–369. [Google Scholar] [CrossRef]
- Gu, N.; Zhang, X.; Gu, X.; Zhao, L.; Godana, E.A.; Xu, M.; Zhang, H. Transcriptomic and proteomic analysis of the mechanisms involved in enhanced disease resistance of strawberries induced by Rhodotorula mucilaginosa cultured with chitosan. Postharvest Biol. Technol. 2021, 172, 111355. [Google Scholar] [CrossRef]
- Lin, R.L.; Yang, Q.Y.; Xiao, J.W.; Solairaj, D.; Ngea, G.L.N.; Zhang, H.Y. Study on the biocontrol effect and physiological mechanism of Hannaella sinensis on the blue mold decay of apples. Int. J. Food Microbiol. 2022, 382, 109931. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.Y.; Zhang, X.; Solairaj, D.; Lin, R.L.; Ackah, M.; Ngea, G.L.N.; Zhang, H.Y. Transcriptomic analyses reveal robust changes in the defense response of apples induced by Hannaella sinensis. Biol. Control 2023, 182, 105237. [Google Scholar] [CrossRef]
- Xu, L.S.; Liu, H.L.; Zhu, S.; Meng, Y.G.; Wang, Y.H.; Li, J.Y.; Zhang, F.R.; Huang, L.L. VmPacC-mediated pH regulation of Valsa mali confers to host acidification identified by comparative proteomics analysis. Stress Biol. 2023, 3, 18. [Google Scholar] [CrossRef]
- Son, S.; Park, S.R. Climate change impedes plant immunity mechanisms. Front. Plant Sci. 2022, 13, 1032820. [Google Scholar] [CrossRef] [PubMed]
- Roston, R.L.; Jouhet, J.; Yu, F.; Gao, H. Editorial: Structure and function of chloroplasts. Front. Plant Sci. 2018, 9, 6–7. [Google Scholar] [CrossRef]
- Liao, G.; Chen, L.; He, Y.; Li, X.; Lv, Z.; Yi, S.; Zhong, M.; Huang, C.; Jia, D.; Qu, X.; et al. Three metabolic pathways are responsible for the accumulation and maintenance of high AsA content in kiwifruit (Actinidia eriantha). BMC Genom. 2021, 22, 13. [Google Scholar] [CrossRef]
- Chen, Z.; Chen, M.; Xu, Z.S.; Li, L.C.; Chen, X.P.; Ma, Y.Z. Characteristics and expression patterns of the aldehyde dehydrogenase (ALDH) gene superfamily of foxtail millet (Setaria italica L.). PLoS ONE 2014, 9, e101136. [Google Scholar] [CrossRef]
- Kaszycki, P.; Dubicka-Lisowska, A.; Augustynowicz, J.; Piwowarczyk, B.; Wesolowski, W. Callitriche cophocarpa (water starwort) proteome under chromate stress: Evidence for induction of a quinone reductase. Environ. Sci. Pollut. Res. 2018, 25, 8928–8942. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Niazi, A.K.; Young, D.; Rosado, L.A.; Vertommen, D.; Bodra, N.; Abdelgawwad, M.R.; Vignols, F.; Wei, B.; Wahni, K.; et al. Self-protection of cytosolic malate dehydrogenase against oxidative stress in Arabidopsis. J. Exp. Bot. 2018, 69, 3491–3505. [Google Scholar] [CrossRef] [Green Version]
- Araji, S.; Grammer, T.A.; Gertzen, R.; Anderson, S.D.; Mikulic-Petkovsek, M.; Veberic, R.; Phu, M.L.; Solar, A.; Leslie, C.A.; Dandekar, A.M.; et al. Novel roles for the polyphenol oxidase enzyme in secondary metabolism and the regulation of cell death in walnut. Plant Physiol. 2014, 164, 1191–1203. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Sun, X. Recent advances in polyphenol oxidase-mediated plant stress responses. Phytochemistry 2021, 181, 112588. [Google Scholar] [CrossRef]
- Vaish, S.; Gupta, D.; Mehrotra, R.; Mehrotra, S.; Basantani, M.K. Glutathione S-transferase: A versatile protein family. 3 Biotech 2020, 10, 321. [Google Scholar] [CrossRef] [PubMed]
- Mugford, S.T.; Milkowski, C. Serine carboxypeptidase-like acyltransferases from plants. Methods Enzym. 2012, 516, 279–297. [Google Scholar] [CrossRef]
- Stuhrwohldt, N.; Buhler, E.; Sauter, M.; Schaller, A. Phytosulfokine (PSK) precursor processing by subtilase SBT3.8 and PSK signaling improve drought stress tolerance in Arabidopsis. J. Exp. Bot. 2021, 72, 3427–3440. [Google Scholar] [CrossRef] [PubMed]
- Lauvergeat, V.; Lacomme, C.; Lacombe, E.; Lasserre, E. Two cinnamoyl-CoA reductase (CCR) genes from Arabidopsis thaliana are differentially expressed during development and in response to infection with pathogenic bacteria. Phytochemistry 2001, 57, 1187–1195. [Google Scholar]
- Yuan, Z.; Yang, H.; Pan, L.; Zhao, W.; Liang, L.; Gatera, A.; Tucker, M.R.; Xu, D. Systematic identification and expression profiles of the BAHD superfamily acyltransferases in barley (Hordeum vulgare). Sci. Rep. 2022, 12, 5063. [Google Scholar] [CrossRef] [PubMed]
- Campos, M.L.; Kang, J.H.; Howe, G.A. Jasmonate-triggered plant immunity. J. Chem. Ecol. 2014, 40, 657–675. [Google Scholar] [CrossRef] [Green Version]
- Zipfel, C. Early molecular events in PAMP-triggered immunity. Curr. Opin. Plant Biol. 2009, 12, 414–420. [Google Scholar] [CrossRef]
- Zhao, G.; Guo, D.; Wang, L.; Li, H.; Wang, C.; Guo, X. Functions of RPM1-interacting protein 4 in plant immunity. Planta 2021, 253, 11. [Google Scholar] [CrossRef]
- Maekawa, S.; Sato, T.; Asada, Y.; Yasuda, S.; Yoshida, M.; Chiba, Y.; Yamaguchi, J. The Arabidopsis ubiquitin ligases ATL31 and ATL6 control the defense response as well as the carbon/nitrogen response. Plant Mol. Biol. 2012, 79, 217–227. [Google Scholar] [CrossRef]
- Oh, I.S.; Park, A.R.; Bae, M.S.; Kwon, S.J.; Kim, Y.S.; Lee, J.E.; Kang, N.Y.; Lee, S.; Cheong, H.; Park, O.K. Secretome analysis reveals an Arabidopsis lipase involved in defense against Alternaria brassicicola. Plant Cell 2005, 17, 2832–2847. [Google Scholar] [CrossRef] [Green Version]
- Kebede, A.; Kebede, M. In silico analysis of promoter region and regulatory elements of glucan endo-1,3-beta-glucosidase encoding genes in Solanum tuberosum: Cultivar DM 1-3 516 R44. J. Genet. Eng. Biotechnol. 2021, 19, 145. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Schumaker, K.S.; Zhu, J.K. Cell signaling during cold, drought, and salt stress. Plant Cell 2002, 14 (Suppl. S1), S165–S183. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.; Song, Y.; Chen, D.; Zang, Y.; Zhang, Q.; Yi, Y.; Qu, G. Genome-wide identification, classification, characterization, and expression analysis of the wall-associated kinase family during fruit development and under wound stress in tomato (Solanum lycopersicum L.). Genes 2020, 11, 1186. [Google Scholar] [CrossRef]
- De Jesus-Pires, C.; Ferreira-Neto, J.R.C.; Pacifico Bezerra-Neto, J.; Kido, E.A.; de Oliveira Silva, R.L.; Pandolfi, V.; Wanderley-Nogueira, A.C.; Binneck, E.; da Costa, A.F.; Pio-Ribeiro, G.; et al. Plant thaumatin-like proteins: Function, evolution and biotechnological applications. Curr. Protein Pept. Sci. 2020, 21, 36–51. [Google Scholar] [CrossRef] [PubMed]
- Hirano, H. Basic 7S globulin in plants. J. Proteom. 2021, 240, 104209. [Google Scholar] [CrossRef]
- Citores, L.; Iglesias, R.; Ferreras, J.M. Antiviral activity of ribosome-Inactivating proteins. Toxins 2021, 13, 80. [Google Scholar] [CrossRef]
- Khare, D.; Choi, H.; Huh, S.U.; Bassin, B.; Kim, J.; Martinoia, E.; Sohn, K.H.; Paek, K.H.; Lee, Y. Arabidopsis ABCG34 contributes to defense against necrotrophic pathogens by mediating the secretion of camalexin. Proc. Natl. Acad. Sci. USA 2017, 114, E5712–E5720. [Google Scholar] [CrossRef]
- Li, R.L.; Wang, J.L.; Xu, L.; Sun, M.H.; Yi, K.K.; Zhao, H.Y. Functional analysis of phosphate transporter OsPHT4 family members in rice. Rice Sci. 2020, 27, 493–503. [Google Scholar] [CrossRef]
- Manghwar, H.; Li, J. Endoplasmic reticulum stress and unfolded protein response signaling in plants. Int. J. Mol. Sci. 2022, 23, 828. [Google Scholar] [CrossRef] [PubMed]
- Su, W.; Liu, Y.; Xia, Y.; Hong, Z.; Li, J. The Arabidopsis homolog of the mammalian OS-9 protein plays a key role in the endoplasmic reticulum-associated degradation of misfolded receptor-like kinases. Mol. Plant 2012, 5, 929–940. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Xu, W.; Xi, H.; Ma, W.; He, Z.; Ma, M. The ER luminal binding protein (BiP) alleviates Cd(2+)-induced programmed cell death through endoplasmic reticulum stress-cell death signaling pathway in tobacco cells. J. Plant Physiol. 2013, 170, 1434–1441. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.J.; Hanisch, S.; Kwaaitaal, M.; Pedersen, C.; Thordal-Christensen, H. A component of the Sec61 ER protein transporting pore is required for plant susceptibility to powdery mildew. Front. Plant Sci. 2013, 4, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Protein Name | Fold Change | Description |
|---|---|---|---|
| Oxidative stress | |||
| MD05G1168400 | ceQORH | 2.35 | Chloroplast envelope quinone oxidoreductase homolog |
| MD08G1005500 | AARE2 | 2.26 | Acylamino-acid-releasing enzyme 2 |
| MD15G1328900 | AAO | 2.21 | L-ascorbate oxidase |
| MD00G1190800 | POD12 | 1.88 | Peroxidase 12 |
| MD12G1218000 | LOX15 | 1.78 | Probable linoleate 9S-lipoxygenase 5 |
| MD15G1187300 | ALDH2C4 | 1.7 | Aldehyde dehydrogenase family 2 member C4 |
| MD02G1028600 | BCB | 1.68 | Blue copper protein |
| MD05G1334200 | FQR1 | 1.65 | NAD(P)H dehydrogenase (quinone) FQR1 |
| MD06G1077500 | SalR | 1.46 | Salutaridine reductase |
| MD03G1240100 | MMDHI | 1.38 | Malate dehydrogenase, mitochondrial |
| Improving plant resistance | |||
| MD05G1320800 | PPO | 2.44 | Polyphenol oxidase latent form, chloroplastic |
| MD06G1003300 | CCR1 | 2.12 | Cinnamoyl-CoA reductase 1 |
| MD16G1149300 | PCC13-62 | 2.00 | Desiccation-related protein PCC13-62 |
| MD02G1273000 | BAHD1 | 1.85 | BAHD acyltransferase At5g47980 |
| MD05G1184300 | GST | 1.8 | Probable glutathione S-transferase |
| MD16G1027500 | SCPL50 | 1.68 | Serine carboxypeptidase-like 50 |
| MD16G1071800 | HTH | 1.57 | Protein HOTHEAD |
| MD15G1078300 | RD21A | 1.53 | Cysteine proteinase RD21A |
| MD01G1118100 | CHI | 1.7 | Chalcone-flavonone isomerase |
| MD15G1194300 | SKIP | 1.52 | SNW/SKI-interacting protein |
| MD10G1066900 | RD19A | 1.48 | Cysteine protease RD19A |
| MD00G1035000 | SBT3.8 | 1.48 | Subtilisin-like protease SBT3.8 |
| MD02G1162200 | SABP2 | 1.47 | Salicylic acid-binding protein 2 |
| MD07G1306900 | FGT | 1.45 | Anthocyanidin 3-O-glucosyltransferase 2 |
| MD04G1139800 | PHOS32 | 1.43 | Universal stress protein PHOS32 |
| MD06G1085500 | 1.37 | Ribosome-inactivating protein bryodin II | |
| MD17G1224900 | HST | 1.64 | Shikimate O-hydroxycinnamoyltransferase |
| MD07G1280800 | UGT88A1 | 1.33 | UDP-glycosyltransferase 88A1 |
| MD00G1038400 | AOS1 | 1.25 | Allene oxide synthase 1, chloroplastic |
| MD16G1047500 | AOC | 1.48 | Allene oxide cyclase, chloroplastic |
| MD15G1402700 | OPR2 | 1.44 | 12-oxophytodienoate reductase 2 |
| MD15G1003500 | 4CLL5 | 1.21 | 4-coumarate—CoA ligase-like 5 |
| MD15G1181900 | AIM1 | 1.48 | Peroxisomal fatty acid beta-oxidation multifunctional protein AIM1 |
| Improving plant disease resistance | |||
| MD05G1172400 | RIN4 | 1.53 | RPM1-interacting protein 4 |
| MD16G1202100 | CML11 | 2.46 | Calmodulin-like protein 11 |
| MD02G1257300 | CDPK1 | 1.6 | Calcium-dependent protein kinase 1 |
| MD09G1157600 | MKK4 | 1.26 | Mitogen-activated protein kinase kinase 4 |
| MD15G1377900 | ATL6 | 1.51 | E3 ubiquitin-protein ligase ATL6 |
| Pathogen cell wall degradation | |||
| MD15G1037300 | ChiC | 2.63 | Class V chitinase |
| MD11G1107600 | GLU | 2.15 | Glucan endo-1,3-beta-glucosidase |
| MD17G1151800 | GLIP1 | 2.93 | GDSL esterase/lipase 1 |
| Signal transduction | |||
| MD03G1020000 | LecRK4 | 2.68 | G-type lectin S-receptor-like serine/threonine-protein kinase LECRK4 |
| MD01G1139800 | WAKL20 | 1.62 | Wall-associated receptor kinase-like 20 |
| MD09G1187500 | NHL6 | 1.47 | NDR1/HIN1-like protein 6 |
| MD08G1031300 | At2g16250 | 1.46 | Probable LRR receptor-like serine/threonine-protein kinase At2g16250 |
| MD10G1102700 | LRR2 | 1.43 | Leucine-rich repeat protein 2 |
| MD16G1004400 | ERL2 | 1.42 | LRR receptor-like serine/threonine-protein kinase ERL2 |
| MD00G1040900 | IPS2 | 1.4 | Inositol-3-phosphate synthase |
| Antibacterial activity | |||
| MD04G1064400 | TLP | 2.11 | Thaumatin-like protein |
| MD11G1287900 | BG7S | 1.94 | Basic 7S globulin |
| MD04G1173600 | LTP | 1.59 | Non-specific lipid-transfer protein |
| MD08G1089100 | SN1 | 1.37 | Snakin-1 |
| Substance transport | |||
| MD14G1018100 | ABCG34 | 2.16 | ABC transporter G family member 34 |
| MD13G1213400 | PHT1-7 | 1.87 | Probable inorganic phosphate transporter 1–7 |
| MD15G1406000 | PATL5 | 1.7 | Patellin-5 |
| Protein processing | |||
| MD03G1297000 | ERDJ3B | 1.39 | DnaJ protein ERDJ3B |
| MD03G1115300 | SC61B | 1.28 | Protein transport protein Sec61 subunit beta |
| MD12G1215200 | BIP5 | 1.21 | Luminal-binding protein 5 |
| MD04G1100100 | P58IPK | 1.27 | DnaJ protein P58IPK homolog |
| MD14G1144700 | CNX1 | 1.28 | Calnexin homolog 1 |
| MD02G1018200 | OS9 | 1.32 | Protein OS-9 homolog |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Q.; Zhang, X.; Solairaj, D.; Lin, R.; Wang, K.; Zhang, H. TMT-Based Proteomic Analysis of Hannaella sinensis-Induced Apple Resistance-Related Proteins. Foods 2023, 12, 2637. https://doi.org/10.3390/foods12142637
Yang Q, Zhang X, Solairaj D, Lin R, Wang K, Zhang H. TMT-Based Proteomic Analysis of Hannaella sinensis-Induced Apple Resistance-Related Proteins. Foods. 2023; 12(14):2637. https://doi.org/10.3390/foods12142637
Chicago/Turabian StyleYang, Qiya, Xi Zhang, Dhanasekaran Solairaj, Rouling Lin, Kaili Wang, and Hongyin Zhang. 2023. "TMT-Based Proteomic Analysis of Hannaella sinensis-Induced Apple Resistance-Related Proteins" Foods 12, no. 14: 2637. https://doi.org/10.3390/foods12142637
APA StyleYang, Q., Zhang, X., Solairaj, D., Lin, R., Wang, K., & Zhang, H. (2023). TMT-Based Proteomic Analysis of Hannaella sinensis-Induced Apple Resistance-Related Proteins. Foods, 12(14), 2637. https://doi.org/10.3390/foods12142637