
Modified Atmosphere Packaging (MAP) for Seaweed Conservation: Impact on Physicochemical Characteristics and Microbiological Activity

 ,
,  , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.1.1. Raw Materials
2.1.2. Sample Processing
2.2. Composition of Headspace Gases
2.3. Physical Characterization
2.3.1. Color Measurement
2.3.2. Texture Analysis
2.4. Microbiological Analysis
2.5. Chemical Characterization
GC–TOF-MS Analysis
2.6. Statistical Analysis
3. Results
3.1. Composition of Headspace Gases
3.2. Physicochemical Characterization
3.2.1. Color
3.2.2. Texture
3.3. Microbiological Analysis
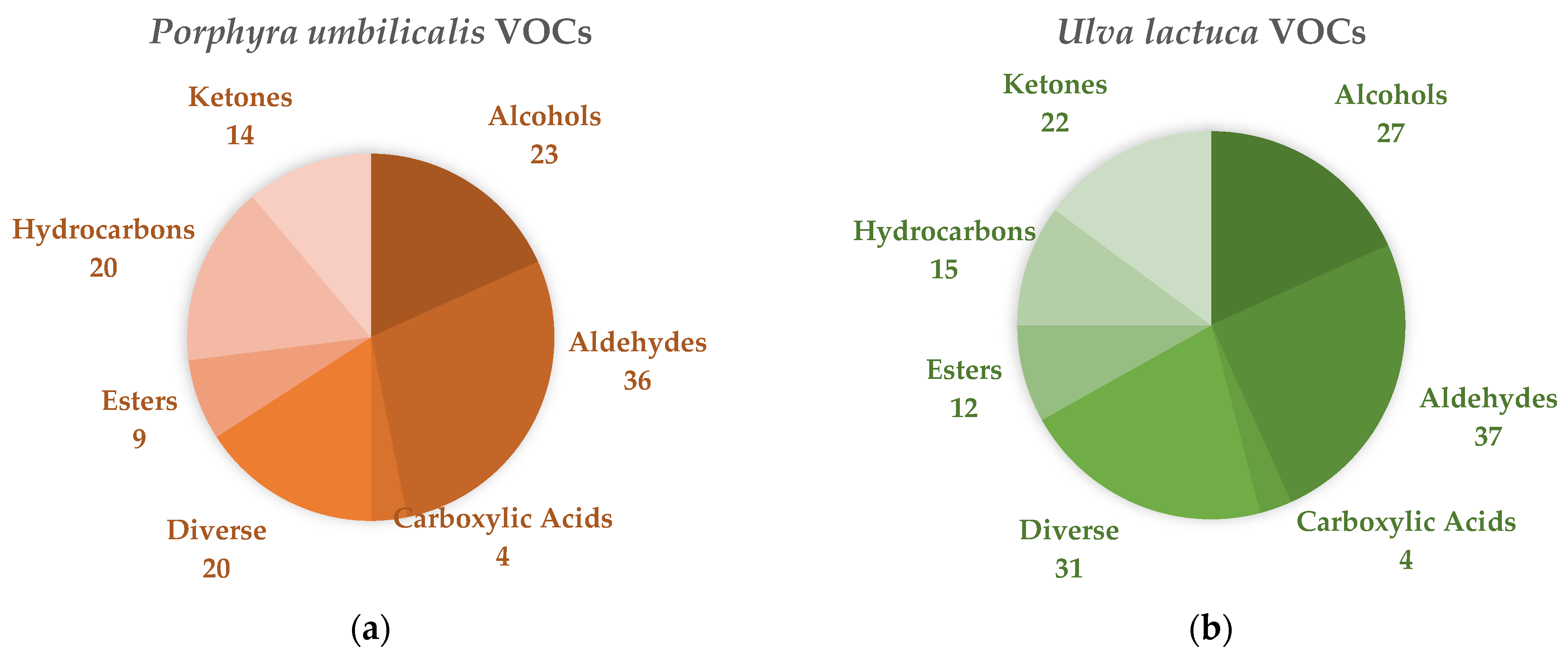
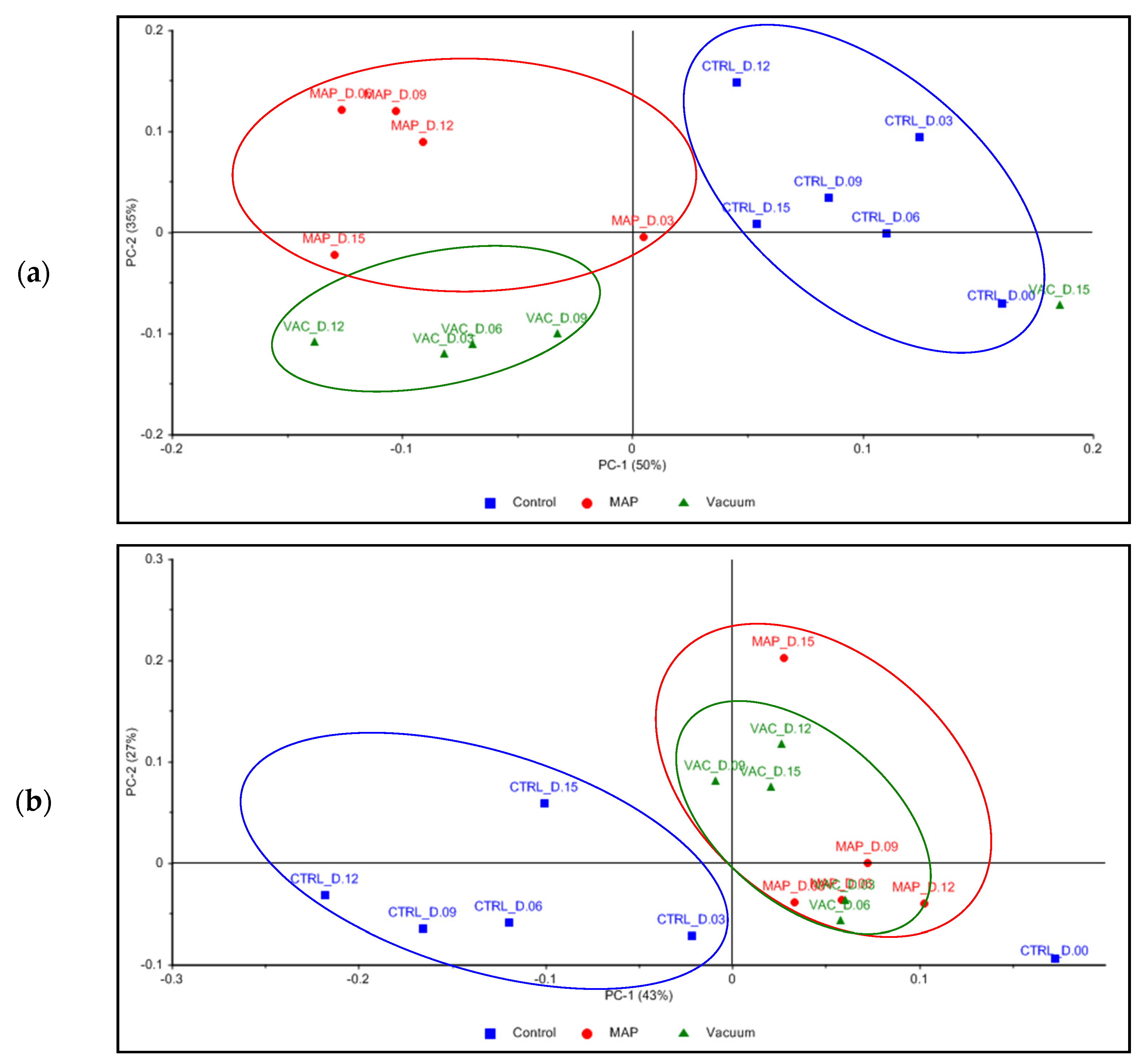
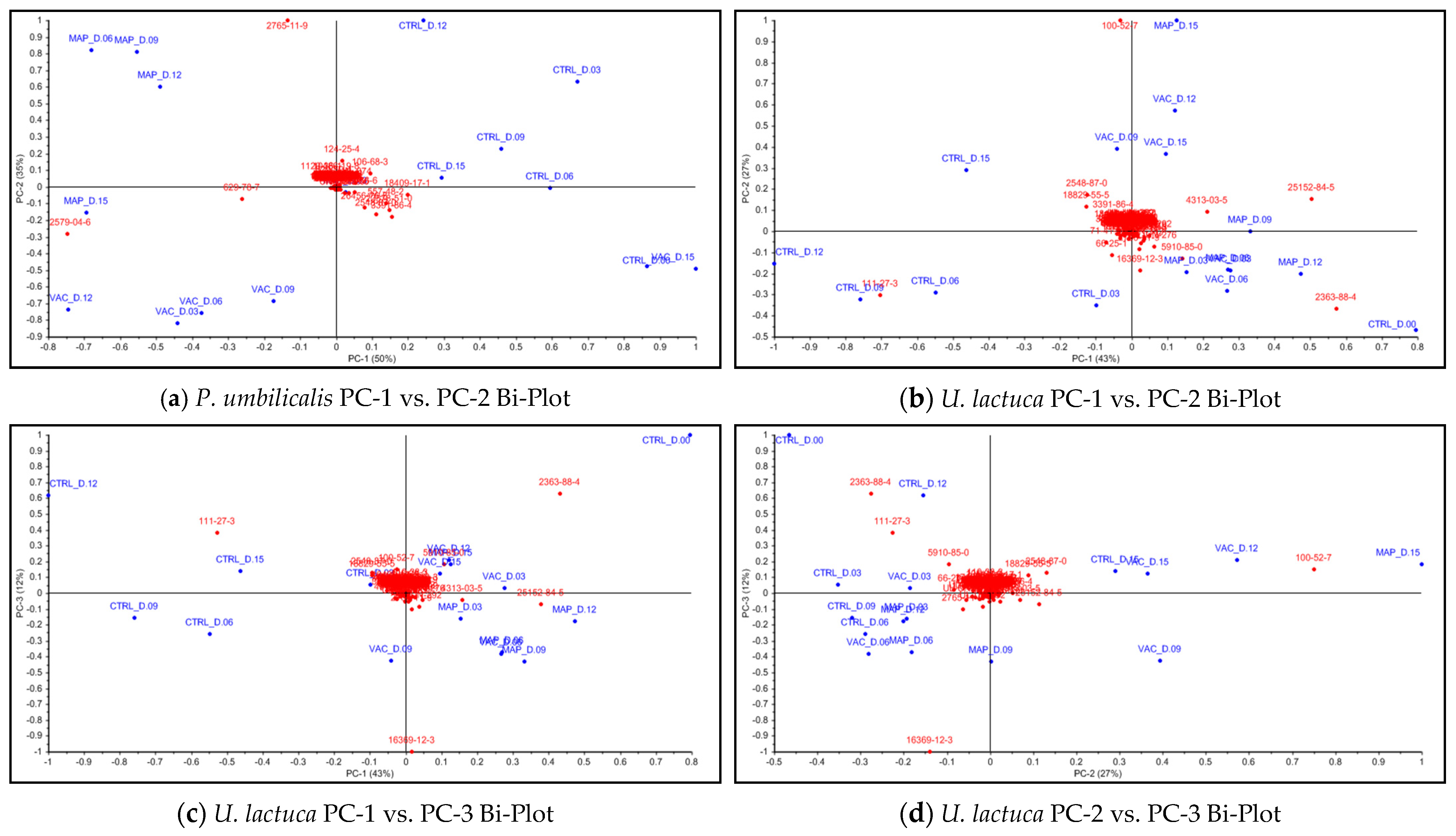
3.4. Chemical Characterization
GC–TOF-MS Analysis
4. Discussion
4.1. Composition of Headspace Gases
4.2. Physicochemical Characterization
4.2.1. Color
4.2.2. Texture
4.3. Microbiological Analysis
4.4. Chemical Characterization
GC–TOF-MS Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, R.E. Phycology, 4th ed.; Cambridge University Press: Cambridge, UK, 2008; ISBN 9780521864084. [Google Scholar]
- Pereira, L. Edible Seaweeds of the World, 1st ed.; CRC Press: Boca Raton, FL, USA, 2016; ISBN 9780429154041. [Google Scholar]
- Naylor, R.L.; Hardy, R.W.; Buschmann, A.H.; Bush, S.R.; Cao, L.; Klinger, D.H.; Little, D.C.; Lubchenco, J.; Shumway, S.E.; Troell, M. A 20-Year Retrospective Review of Global Aquaculture. Nature 2021, 591, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine Natural Products. Nat. Prod. Rep. 2018, 35, 8–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mouritsen, O.G.; Rhatigan, P.; Pérez-Lloréns, J.L. The Rise of Seaweed Gastronomy: Phycogastronomy. Bot. Mar. 2019, 62, 195–209. [Google Scholar] [CrossRef]
- Mišurcová, L. Chemical Composition of Seaweeds. In Handbook of Marine Macroalgae: Biotechnology and Applied Phycology; Kim, S.-K., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2012; pp. 173–192. [Google Scholar]
- Mouritsen, O.G. The Emerging Science of Gastrophysics and Its Application to the Algal Cuisine. Flavour 2012, 1, 6. [Google Scholar] [CrossRef] [Green Version]
- Fleurence, J. Seaweeds as Food. In Seaweed in Health and Disease Prevention; Fleurence, J., Levine, I., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 149–167. ISBN 9780128027936. [Google Scholar]
- Rioux, L.-E.; Beaulieu, L.; Turgeon, S.L. Seaweeds: A Traditional Ingredients for New Gastronomic Sensation. Food Hydrocoll. 2017, 68, 255–265. [Google Scholar] [CrossRef]
- United Nations; Department of Economic and Social Affairs. Population Division World Population Prospects 2019: Highlights (ST/ESA/SER.A/423); Department of Economic and Social Affairs: New York, NY, USA, 2019. [Google Scholar]
- Vieira, E.F.; Soares, C.; Machado, S.; Correia, M.; Ramalhosa, M.J.; Oliva-teles, M.T.; Carvalho, A.P.; Domingues, V.F.; Antunes, F.; Oliveira, T.A.C.; et al. Seaweeds from the Portuguese Coast as a Source of Proteinaceous Material: Total and Free Amino Acid Composition Profile. Food Chem. 2018, 269, 264–275. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.S. Food Preservation: An Overview. In Handbook of Food Preservation; Rahman, M.S., Ed.; CRC Press: Boca Raton, FL, USA, 2020; pp. 7–18. ISBN 9780429091483. [Google Scholar]
- Rahman, M.S. Types of Foods and Food Products. In Handbook of Food Preservation; Rahman, M.S., Ed.; CRC Press: Boca Raton, FL, USA, 2020; pp. 3–6. ISBN 9780429091483. [Google Scholar]
- Maarse, H. Volatile Compounds in Foods and Beverages. In Food Science and Technology, 1st ed.; Marcel Dekker, Inc.: New York, NY, USA, 1991; ISBN 0-8247-8390-5. [Google Scholar]
- De Quirós, A.R.B.; López-Hernández, J. An Overview on Effects of Processing on the Nutritional Content and Bioactive Compounds in Seaweeds. Foods 2021, 10, 2168. [Google Scholar] [CrossRef]
- Kadam, S.U.; Álvarez, C.; Tiwari, B.K.; O’Donnell, C.P. Processing of Seaweeds. In Seaweed Sustainability; Elsevier: Amsterdam, The Netherlands, 2015; pp. 61–78. ISBN 9780124199583. [Google Scholar]
- Berk, Z. Thermal Processes, Methods, and Equipment. In Food Process Engineering and Technology; Academic Press: London, UK, 2018; pp. 421–438. [Google Scholar]
- Zhu, X.; Healy, L.E.; Sevindik, O.; Sun, D.W.; Selli, S.; Kelebek, H.; Tiwari, B.K. Impacts of Novel Blanching Treatments Combined with Commercial Drying Methods on the Physicochemical Properties of Irish Brown Seaweed Alaria Esculenta. Food Chem. 2022, 369, 130949. [Google Scholar] [CrossRef]
- Sappati, P.K.; Nayak, B.; van Walsum, G.P. Effect of Glass Transition on the Shrinkage of Sugar Kelp (Saccharina Latissima) during Hot Air Convective Drying. J. Food Eng. 2017, 210, 50–61. [Google Scholar] [CrossRef]
- Pinheiro, V.F.; Marçal, C.; Abreu, H.; da Silva, J.A.L.; Silva, A.M.S.S.; Cardoso, S.M. Physicochemical Changes of Air-Dried and Salt-Processed Ulva Rigida over Storage Time. Molecules 2019, 24, 2955. [Google Scholar] [CrossRef] [Green Version]
- Nongonierma, A.; Voilley, A.; Cayot, P.; Le Quéré, J.-L.; Springett, M. Mechanisms of Extraction of Aroma Compounds from Foods, Using Adsorbents. Effect of Various Parameters. Food Rev. Int. 2006, 22, 51–94. [Google Scholar] [CrossRef]
- López-Pérez, O.; del Olmo, A.; Picon, A.; Nuñez, M. Volatile Compounds and Odour Characteristics during Long-Term Storage of Kombu Seaweed (Laminaria Ochroleuca) Preserved by High Pressure Processing, Freezing and Salting. LWT 2020, 118, 108710. [Google Scholar] [CrossRef]
- Akomea-Frempong, S.; Perry, J.J.; Skonberg, D.I. Effects of Pre-Freezing Blanching Procedures on the Physicochemical Properties and Microbial Quality of Frozen Sugar Kelp. J. Appl. Phycol. 2022, 34, 609–624. [Google Scholar] [CrossRef]
- Li, D.; Zhu, Z.; Sun, D.W. Effects of Freezing on Cell Structure of Fresh Cellular Food Materials: A Review. Trends Food Sci. Technol. 2018, 75, 46–55. [Google Scholar] [CrossRef]
- Obluchinskaya, E.; Daurtseva, A. Effects of Air Drying and Freezing and Long-Term Storage on Phytochemical Composition of Brown Seaweeds. J. Appl. Phycol. 2020, 32, 4235–4249. [Google Scholar] [CrossRef]
- Fujimura, T.; Kawai, T. Enzymes and Seaweed Flavor. In Seafood Enzymes; Haard, N.F., Simpson, B.K., Eds.; CRC Press: New York, NY, USA, 2000; pp. 411–436. [Google Scholar]
- Le Pape, M.-A.; Grua-Priol, J.; Demaimay, M.; Crua-Priol, J. Effect of Two Storage Conditions on the Odor of an Edible Seaweed, Palmaria Palmata, and Optimization of an Extraction Procedure Preserving Its Odor Characteristics. J. Food Sci. 2002, 67, 3135–3139. [Google Scholar] [CrossRef]
- Bennamoun, L.; Afzal, M.T.; Léonard, A. Drying of Alga as a Source of Bioenergy Feedstock and Food Supplement—A Review. Renew. Sustain. Energy Rev. 2015, 50, 1203–1212. [Google Scholar] [CrossRef]
- Uribe, E.; Pardo-Orellana, C.M.; Vega-Gálvez, A.; Ah-Hen, K.S.; Pastén, A.; García, V.; Aubourg, S.P. Effect of Drying Methods on Bioactive Compounds, Nutritional, Antioxidant, and Antidiabetic Potential of Brown Alga Durvillaea Antarctica . Dry. Technol. 2020, 38, 1915–1928. [Google Scholar] [CrossRef]
- Amorim, A.M.; Nardelli, A.E.; Chow, F. Effects of Drying Processes on Antioxidant Properties and Chemical Constituents of Four Tropical Macroalgae Suitable as Functional Bioproducts. J. Appl. Phycol. 2020, 32, 1495–1509. [Google Scholar] [CrossRef]
- Badmus, U.O.; Taggart, M.A.; Boyd, K.G. The Effect of Different Drying Methods on Certain Nutritionally Important Chemical Constituents in Edible Brown Seaweeds. J. Appl. Phycol. 2019, 31, 3883–3897. [Google Scholar] [CrossRef] [Green Version]
- Del Olmo, A.; Picon, A.; Nuñez, M. Preservation of Five Edible Seaweeds by High Pressure Processing: Effect on Microbiota, Shelf Life, Colour, Texture and Antioxidant Capacity. Algal Res. 2020, 49, 101938. [Google Scholar] [CrossRef]
- Del Olmo, A.; Picon, A.; Nuñez, M. High Pressure Processing for the Extension of Laminaria Ochroleuca (Kombu) Shelf-Life: A Comparative Study with Seaweed Salting and Freezing. Innov. Food Sci. Emerg. Technol. 2019, 52, 420–428. [Google Scholar] [CrossRef]
- Zhang, M.; Meng, X.; Bhandari, B.; Fang, Z. Recent Developments in Film and Gas Research in Modified Atmosphere Packaging of Fresh Foods. Crit. Rev. Food Sci. Nutr. 2016, 56, 2174–2182. [Google Scholar] [CrossRef] [PubMed]
- Paull, R.E.; Chen, N.J. Postharvest Handling and Storage of the Edible Red Seaweed Gracilaria. Postharvest Biol. Technol. 2008, 48, 302–308. [Google Scholar] [CrossRef]
- Araujo, G.S.; Cotas, J.; Morais, T.; Leandro, A.; García-Poza, S.; Gonçalves, A.M.M.; Pereira, L. Calliblepharis Jubata Cultivation Potential—A Comparative Study between Controlled and Semi-Controlled Aquaculture. Appl. Sci. 2020, 10, 7553. [Google Scholar] [CrossRef]
- García-Poza, S.; Morais, T.; Leandro, A.; Cotas, J.; Marques, J.C.; Pereira, L.; Gonçalves, A.M.M.M. A Comparative Study of the Fatty Acids and Monosaccharides of Wild and Cultivated Ulva sp. J. Mar. Sci. Eng. 2022, 10, 233. [Google Scholar] [CrossRef]
- Galgano, F.; Caruso, M.C.; Ventura, N.M.; Magno, C.; Favati, F. Effects of Anti-UV Film and Protective Atmosphere on Fresh-Cut Iceberg Lettuce Preservation. Acta Aliment. 2017, 46, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Brenes, A.L.M.; Brown, W.; Steinmaus, S.; Brecht, J.K.; Xie, Y.; Bornhorst, E.R.; Luo, Y.; Zhou, B.; Shaw, A.; Vorst, K. Temperature Profiling of Open- and Closed-Doored Produce Cases in Retail Grocery Stores. Food Control 2020, 113, 107158. [Google Scholar] [CrossRef]
- Licciardello, F.; Cipri, L.; Muratore, G. Influence of Packaging on the Quality Maintenance of Industrial Bread by Comparative Shelf Life Testing. Food Packag. Shelf Life 2014, 1, 19–24. [Google Scholar] [CrossRef]
- Wrolstad, R.E.; Smith, D.E. Color Analysis. In Food Analysis; Food Science Text Series; Nielsen, S.S., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 545–555. ISBN 978-3-319-45774-1. [Google Scholar]
- Lubsch, A.; Timmermans, K. Texture Analysis of Laminaria Digitata (Phaeophyceae) Thallus Reveals Trade-off between Tissue Tensile Strength and Toughness along Lamina. Bot. Mar. 2017, 60, 229–237. [Google Scholar] [CrossRef]
- Balbas, J.; Hamid, N.; Liu, T.; Kantono, K.; Robertson, J.; White, W.L.; Ma, Q.; Lu, J. Comparison of Physicochemical Characteristics, Sensory Properties and Volatile Composition between Commercial and New Zealand Made Wakame from Undaria Pinnatifida. Food Chem. 2015, 186, 168–175. [Google Scholar] [CrossRef] [PubMed]
- ISO 16649-2:2001; Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Enumeration of Beta-Glucuronidase-Positive Escherichia Coli—Part 2: Colony-Count Technique at 44 °C Using 5-Bromo-4-chloro-3-indolyl Beta-D-glucuronide. 1st. International Organization for Standardization: Geneva, Switzerland, 2001.
- ISO 6888-2:2021; Microbiology of the Food Chain—Horizontal Method for the Enumeration of Coagulase-Positive Staphylococci (Staphylococcus Aureus and Other Species)—Part 2: Method Using Rabbit Plasma Fibrinogen Agar Medium. 2nd. International Organization for Standardization: Geneva, Switzerland, 2021.
- ISO 21872-1:2017; Microbiology of the Food Chain—Horizontal Method for the Determination of Vibrio spp.—Part 1: Detection of Potentially Enteropathogenic Vibrio Parahaemolyticus, Vibrio Cholerae and Vibrio Vulnificus. 1st. International Organization for Standardization: Geneva, Switzerland, 2017.
- ISO 6579-1:2017; Microbiology of the Food Chain—Horizontal Method for the Detection, Enumeration and Serotyping of Salmonella—Part 1: Detection of Salmonella spp. 1st. International Organization for Standardization: Geneva, Switzerland, 2017.
- ISO 11290-2:2017; Microbiology of the Food Chain—Horizontal Method for the Detection and Enumeration of Listeria Monocytogenes and of Listeria spp.—Part 2: Enumeration Method. 2nd. International Organization for Standardization: Geneva, Switzerland, 2017.
- Biokar Diagnostics. Technical Data Sheet—Compass® Ecc Agar—Enumeration of Escherichia Coli and Other Coliforms. Biokar Diagnostics: Beauvais, France. 2016. Available online: https://www.solabia.com/fr/Produto_291,9/BIOKAR-Diagnostics/COMPASS-ECC-AGAR.html (accessed on 17 July 2022).
- Condalab Marine Agar. Available online: https://www.condalab.com/int/en/index.php?controller=attachment&id_attachment=24225 (accessed on 2 May 2022).
- CEVA Macroalgues et Microalgues Alimentaires—Statut Règlementaire En France et En Europe (Synthèse CEVA 2019); Centre d’Étude et de Valorisation des Algues: Pleubian, France, 2019; Available online: https://www.ceva-algues.com/document/synthese-reglementaire-algues-alimentaires/ (accessed on 17 July 2022).
- INSA Interpretação de Resultados de Ensaios Microbiológicos Em Alimentos Prontos Para Consumo e Em Superfícies Do Ambiente de Preparação e Distribuição Alimentar (Valores-Guia); Instituto Nacional de Saúde Doutor Ricardo Jorge: Lisboa, Portugal, 2019; Available online: https://www.insa.min-saude.pt/wp-content/uploads/2019/12/INSA_Valores-guia.pdf (accessed on 17 July 2022).
- Van Den Dool, H.; Kratz, P.D. A Generalization of the Retention Index System Including Linear Temperature Programmed Gas—Liquid Partition Chromatography. J. Chromatogr. A 1963, 11, 463–471. [Google Scholar] [CrossRef]
- RStudio Team. RStudio: Integrated Development for R 2022; RStudio Team: Boston, MA, USA, 2022. [Google Scholar]
- Elkin, L.A.; Kay, M.; Higgins, J.J.; Wobbrock, J.O. An Aligned Rank Transform Procedure for Multifactor Contrast Tests. In Proceedings of the 34th Annual ACM Symposium on User Interface Software and Technology, Virtual, 10–14 October 2021; ACM: New York, NY, USA, 2021; Volume 1, pp. 754–768. [Google Scholar] [CrossRef]
- López-Pérez, O.; del Olmo, A.; Picon, A.; Nuñez, M. Volatile Compounds and Odour Characteristics of Five Edible Seaweeds Preserved by High Pressure Processing: Changes during Refrigerated Storage. Algal Res. 2021, 53, 102137. [Google Scholar] [CrossRef]
- Ren, C.; Liu, Z.; Wang, X.; Qin, S. The Seaweed Holobiont: From Microecology to Biotechnological Applications. Microb. Biotechnol. 2022, 15, 738–754. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Gionfriddo, E.; Acquaro, V.; Pawliszyn, J. Direct Immersion Solid-Phase Microextraction Analysis of Multi-Class Contaminants in Edible Seaweeds by Gas Chromatography-Mass Spectrometry. Anal. Chim. Acta 2018, 1031, 83–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Gemert, L.J. Odour Thresholds: Compilations of Odour Threshold Values in Air, Water and Other Media, 2nd ed.; Oliemans Punter & Partners: Utrecht, The Netherlands, 2011; ISBN 978-90-810894-0-1. [Google Scholar]
- Allende, A.; Luo, Y.; McEvoy, J.L.; Artés, F.; Wang, C.Y. Microbial and Quality Changes in Minimally Processed Baby Spinach Leaves Stored under Super Atmospheric Oxygen and Modified Atmosphere Conditions. Postharvest Biol. Technol. 2004, 33, 51–59. [Google Scholar] [CrossRef]
- Bidwell, R.G.S.; Mclachlan, J. Carbon Nutrition of Seaweeds: Photosynthesis, Photorespiration and Respiration. J. Exp. Mar. Bio. Ecol. 1985, 86, 15–46. [Google Scholar] [CrossRef]
- Fernández, P.A.; Hurd, C.L.; Roleda, M.Y. Bicarbonate Uptake via an Anion Exchange Protein Is the Main Mechanism of Inorganic Carbon Acquisition by the Giant Kelp Macrocystis Pyrifera (Laminariales, Phaeophyceae) under Variable PH. J. Phycol. 2014, 50, 998–1008. [Google Scholar] [CrossRef]
- Mrkić, S.; Galić, K.; Ivanković, M.; Hamin, S.; Ciković, N. Gas Transport and Thermal Characterization of Mono- and Di-Polyethylene Films Used for Food Packaging. J. Appl. Polym. Sci. 2006, 99, 1590–1599. [Google Scholar] [CrossRef]
- Pinheiro, A.C.D.A.S.; Urbinati, E.; Tappi, S.; Picone, G.; Patrignani, F.; Lanciotti, R.; Romani, S.; Rocculi, P. The Impact of Gas Mixtures of Argon and Nitrous Oxide (N2O) on Quality Parameters of Sardine (Sardina Pilchardus) Fillets during Refrigerated Storage. Food Res. Int. 2019, 115, 268–275. [Google Scholar] [CrossRef]
- Carvalho, M.; Eyre, B. Carbon Stable Isotope Discrimination during Respiration in Three Seaweed Species. Mar. Ecol. Prog. Ser. 2011, 437, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Rosenberg, G.; Littler, D.S.; Littler, M.M.; Oliveira, E.C. Primary Production and Photosynthetic Quotients of Seaweeds from São Paulo State, Brazil. Bot. Mar. 1995, 38, 369–377. [Google Scholar] [CrossRef]
- Sand-Jensen, K.; Binzer, T.; Middelboe, A.L. Scaling of Photosynthetic Production of Aquatic Macrophytes: A Review. Oikos 2007, 116, 280–294. [Google Scholar] [CrossRef]
- Nakamura, M.; Kumagai, N.H.; Tamaoki, M.; Arita, K.; Ishii, Y.; Nakajima, N.; Yabe, T. Photosynthesis and Growth of Ulva Ohnoi and Ulva Pertusa (Ulvophyceae) under High Light and High Temperature Conditions, and Implications for Green Tide in Japan. Phycol. Res. 2020, 68, 152–160. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.; Xu, D.; Wang, Y.; Zhang, X.; Cao, S.; Mou, S.; Ye, N. The Effect of Nutrient Concentrations, Nutrient Ratios and Temperature on Photosynthesis and Nutrient Uptake by Ulva Prolifera: Implications for the Explosion in Green Tides. J. Appl. Phycol. 2014, 26, 537–544. [Google Scholar] [CrossRef]
- Figueroa, F.; Israel, A.; Neori, A.; Martínez, B.; Malta, E.; Ang, P.; Inken, S.; Marquardt, R.; Korbee, N. Effects of Nutrient Supply on Photosynthesis and Pigmentation in Ulva Lactuca (Chlorophyta): Responses to Short-Term Stress. Aquat. Biol. 2009, 7, 173–183. [Google Scholar] [CrossRef]
- Mercado, J.M.; Salles, S.; Cortés, D. Ecophysiology of Marine Algae. In Encyclopedia of Marine Biotechnology; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2020; pp. 341–357. [Google Scholar] [CrossRef]
- Sampath-Wiley, P.; Neefus, C.D.; Jahnke, L.S. Seasonal Effects of Sun Exposure and Emersion on Intertidal Seaweed Physiology: Fluctuations in Antioxidant Contents, Photosynthetic Pigments and Photosynthetic Efficiency in the Red Alga Porphyra Umbilicalis Kützing (Rhodophyta, Bangiales). J. Exp. Mar. Bio. Ecol. 2008, 361, 83–91. [Google Scholar] [CrossRef]
- Feng, Z.; Wu, L.; Sun, Z.; Yang, J.; Liu, G.; Niu, J.; Wang, G. Control of Reactive Oxygen Species through Antioxidant Enzymes Plays a Pivotal Role during the Cultivation of Neopyropia Yezoensis . J. Mar. Sci. Eng. 2022, 10, 109. [Google Scholar] [CrossRef]
- Edreva, A. Generation and Scavenging of Reactive Oxygen Species in Chloroplasts: A Submolecular Approach. Agric. Ecosyst. Environ. 2005, 106, 119–133. [Google Scholar] [CrossRef]
- Vermaat, J.E.; Sand-Jensen, K. Survival, Metabolism and Growth of Ulva Lactuca under Winter Conditions: A Laboratory Study of Bottlenecks in the Life Cycle. Mar. Biol. 1987, 95, 55–61. [Google Scholar] [CrossRef]
- Lee, H.; Park, D.H.; Kim, E.J.; Choi, M.-J. Freshness Analysis of Raw Laver (Pyropia Yenzoensis) Conserved under Supercooling Conditions. Foods 2023, 12, 510. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.; Lee, E.; Choe, E. Light Effects on Lipid Oxidation, Antioxidants, and Pigments in Dried Laver (Porphyra) during Storage. Food Sci. Biotechnol. 2014, 23, 701–709. [Google Scholar] [CrossRef]
- Munier, M.; Jubeau, S.; Wijaya, A.; Morançais, M.; Dumay, J.; Marchal, L.; Jaouen, P.; Fleurence, J. Physicochemical Factors Affecting the Stability of Two Pigments: R-Phycoerythrin of Grateloupia Turuturu and B-Phycoerythrin of Porphyridium Cruentum. Food Chem. 2014, 150, 400–407. [Google Scholar] [CrossRef]
- Sánchez-García, F.; Hernández, I.; Palacios, V.M.; Roldán, A.M. Freshness Quality and Shelf Life Evaluation of the Seaweed Ulva Rigida through Physical, Chemical, Microbiological, and Sensory Methods. Foods 2021, 10, 181. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.; Abreu, H.; Silva, A.; Cardoso, S. Effect of Oven-Drying on the Recovery of Valuable Compounds from Ulva Rigida, Gracilaria Sp. and Fucus Vesiculosus . Mar. Drugs 2019, 17, 90. [Google Scholar] [CrossRef] [Green Version]
- Harrysson, H.; Krook, J.L.; Larsson, K.; Tullberg, C.; Oerbekke, A.; Toth, G.; Pavia, H.; Undeland, I. Effect of Storage Conditions on Lipid Oxidation, Nutrient Loss and Colour of Dried Seaweeds, Porphyra Umbilicalis and Ulva Fenestrata, Subjected to Different Pretreatments. Algal Res. 2021, 56, 102295. [Google Scholar] [CrossRef]
- Baldermann, S.; Yamamoto, M.; Yang, Z.; Kawahashi, T.; Kuwano, K.; Watanabe, N. C13-Apocarotenoids: More than Flavor Compounds? In Carotenoid Cleavage Products; American Chemical Society: Washington, DC, USA, 2013; pp. 73–80. [Google Scholar] [CrossRef]
- Pérez-Gálvez, A.; Roca, M. Phyllobilins: A New Group of Bioactive Compounds. In Studies in Natural Products Chemistry; Atta-ur-Rahman, F., Ed.; Elsevier B.V: Amsterdam, The Netherlands, 2017; Volume 52, pp. 159–191. ISBN 9780444639318. [Google Scholar]
- Chen, K.; Ríos, J.J.; Pérez-Gálvez, A.; Roca, M. Comprehensive Chlorophyll Composition in the Main Edible Seaweeds. Food Chem. 2017, 228, 625–633. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.; Shin, M.; Lee, K.; Choe, E. Effects of Water Activity on Pigments in Dried Laver (Porphyra) during Storage. Food Sci. Biotechnol. 2013, 22, 1523–1529. [Google Scholar] [CrossRef]
- Eismann, A.I.; Reis, R.P.; Da Silva, A.F.; Cavalcanti, D.N. Ulva Spp. Carotenoids: Responses to Environmental Conditions. Algal Res. 2020, 48, 101916. [Google Scholar] [CrossRef]
- Spencer, K.C.; Humphreys, D.J. Argon Packaging and Processing Preserves and Enhances Flavor, Freshness, and Shelf Life of Foods. In Freshness and Shelf Life of Foods; Cadwallader, K.R., Weenen, H., Eds.; American Chemical Society: Washington, DC, USA, 2002; Volume 836, pp. 270–291. [Google Scholar]
- Zhu, S.; Gu, D.; Lu, C.; Zhang, C.; Chen, J.; Yang, R.; Luo, Q.; Wang, T.; Zhang, P.; Chen, H. Cold Stress Tolerance of the Intertidal Red Alga Neoporphyra Haitanensis . BMC Plant Biol. 2022, 22, 114. [Google Scholar] [CrossRef]
- Takahashi, M.; Kumari, P.; Li, C.; Mikami, K. Low Temperature Causes Discoloration by Repressing Growth and Nitrogen Transporter Gene Expression in the Edible Red Alga Pyropia Yezoensis . Mar. Environ. Res. 2020, 159, 105004. [Google Scholar] [CrossRef] [PubMed]
- Kakinuma, M.; Coury, D.A.; Nakamoto, C.; Sakaguchi, K.; Amano, H. Molecular Analysis of Physiological Responses to Changes in Nitrogen in a Marine Macroalga, Porphyra Yezoensis (Rhodophyta). Cell Biol. Toxicol. 2008, 24, 629–639. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.B.; Lee, W.-C.; Kim, H.C.; Hong, S. Photosynthetic Characteristics of Pyropia Yezoensis (Ueda) Hwang & Choi Measured Using Diving-PAM in the Jindo-Haenam Region on the Southwestern Coast of the Korean Peninsula. J. Appl. Phycol. 2020, 32, 2631–2640. [Google Scholar] [CrossRef]
- Nayyar, D.; Skonberg, D.I. Contrasting Effects of Two Storage Temperatures on the Microbial, Physicochemical, and Sensory Properties of Two Fresh Red Seaweeds, Palmaria Palmata and Gracilaria Tikvahiae . J. Appl. Phycol. 2019, 31, 731–739. [Google Scholar] [CrossRef]
- Gupta, S.; Rajauria, G.; Abu-Ghannam, N. Study of the Microbial Diversity and Antimicrobial Properties of Irish Edible Brown Seaweeds. Int. J. Food Sci. Technol. 2010, 45, 482–489. [Google Scholar] [CrossRef] [Green Version]
- Liot, F.; Colin, A.; Mabeau, S. Microbiology and Storage Life of Fresh Edible Seaweeds. J. Appl. Phycol. 1993, 5, 243–247. [Google Scholar] [CrossRef]
- Moore, J.E.; Xu, J.; Millar, B.C. Diversity of the Microflora of Edible Macroalga (Palmaria Palmata). Food Microbiol. 2002, 19, 249–257. [Google Scholar] [CrossRef]
- Uchida, M.; Miyoshi, T.; Yoshida, G.; Niwa, K.; Mori, M.; Wakabayashi, H. Isolation and Characterization of Halophilic Lactic Acid Bacteria Acting as a Starter Culture for Sauce Fermentation of the Red Alga Nori (Porphyra Yezoensis). J. Appl. Microbiol. 2014, 116, 1506–1520. [Google Scholar] [CrossRef] [PubMed]
- Embleni, A.; Emblem, A. Modified Atmosphere Packaging and Other Active Packaging Systems for Food, Beverages and Other Fast-Moving Consumer Goods. In Trends in Packaging of Food, Beverages and Other Fast-Moving Consumer Goods (FMCG); Woodhead Publishing: Cambridge, UK, 2013; pp. 22–34. [Google Scholar] [CrossRef]
- Løvdal, T.; Lunestad, B.T.; Myrmel, M.; Rosnes, J.T.; Skipnes, D. Microbiological Food Safety of Seaweeds. Foods 2021, 10, 2719. [Google Scholar] [CrossRef]
- Del Olmo, A.; Picon, A.; Nuñez, M. The Microbiota of Eight Species of Dehydrated Edible Seaweeds from North West Spain. Food Microbiol. 2018, 70, 224–231. [Google Scholar] [CrossRef]
- Picon, A.; del Olmo, A.; Nuñez, M. Bacterial Diversity in Six Species of Fresh Edible Seaweeds Submitted to High Pressure Processing and Long-Term Refrigerated Storage. Food Microbiol. 2021, 94, 103646. [Google Scholar] [CrossRef] [PubMed]
- Cox, S.; Abu-Ghannam, N.; Gupta, S. An Assessment of the Antioxidant and Antimicrobial Activity of Six Species of Edible Irish Seaweeds. Int. Food Res. J. 2010, 17, 205–220. [Google Scholar] [CrossRef]
- Goecke, F.; Labes, A.; Wiese, J.; Imhoff, J. Chemical Interactions between Marine Macroalgae and Bacteria. Mar. Ecol. Prog. Ser. 2010, 409, 267–299. [Google Scholar] [CrossRef]
- Hollants, J.; Leliaert, F.; De Clerck, O.; Willems, A. What We Can Learn from Sushi: A Review on Seaweed-Bacterial Associations. FEMS Microbiol. Ecol. 2013, 83, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Figueroa, V.; Bunger, A.; Ortiz, J.; Aguilera, J.M. Sensory Descriptors for Three Edible Chilean Seaweeds and Their Relations to Umami Components and Instrumental Texture. J. Appl. Phycol. 2022, 34, 3141–3156. [Google Scholar] [CrossRef]
- Salgado, A.; Moreira-Leite, B.; Afonso, A.; Infante, P.; Mata, P. Chocolates Enriched with Seaweed: Sensory Profiling and Consumer Segmentation. Int. J. Gastron. Food Sci. 2023, 33, 100747. [Google Scholar] [CrossRef]
- López-Pérez, O.; Picon, A.; Nuñez, M. Volatile Compounds and Odour Characteristics of Seven Species of Dehydrated Edible Seaweeds. Food Res. Int. 2017, 99, 1002–1010. [Google Scholar] [CrossRef]
- Kladi, M.; Vagias, C.; Roussis, V. Volatile Halogenated Metabolites from Marine Red Algae. Phytochem. Rev. 2004, 3, 337–366. [Google Scholar] [CrossRef]
- Vilar, E.G.; O’Sullivan, M.G.; Kerry, J.P.; Kilcawley, K.N. A Chemometric Approach to Characterize the Aroma of Selected Brown and Red Edible Seaweeds/Extracts. J. Sci. Food Agric. 2021, 101, 1228–1238. [Google Scholar] [CrossRef]
- Zhang, M.; Li, R.-X.; Hu, C.-M.; Yang, L.-E.; Tang, J.; Lu, Q.-Q.; Zhang, T.; Shen, Z.-G.; Shen, S.-D.; Xu, P.; et al. The Metabolism of 8-Heptadecene in Pyropia (Bangiaceae, Rhodophyta). J. Appl. Phycol. 2014, 26, 1181–1187. [Google Scholar] [CrossRef]
- He, L.; Wang, L.; Wang, L.; Shen, S. Effects of Methyl Jasmonate on the Composition of Volatile Compounds in Pyropia Yezoensis . J. Ocean Univ. China 2018, 17, 291–296. [Google Scholar] [CrossRef]
- Vilar, E.G.; O’Sullivan, M.G.; Kerry, J.P.; Kilcawley, K.N. Volatile Compounds of Six Species of Edible Seaweed: A Review. Algal Res. 2020, 45, 101740. [Google Scholar] [CrossRef]
- Li, S.; Hu, M.; Tong, Y.Y.; Xia, Z.; Tong, Y.Y.; Sun, Y.; Cao, J.; Zhang, J.; Liu, J.; Zhao, S.; et al. A Review of Volatile Compounds in Edible Macroalgae. Food Res. Int. 2023, 165, 112559. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Zhu, Z.; Zhu, Z.; Yang, R.; Qian, F.; Chen, H.; Yan, X. Different Responses to Heat Shock Stress Revealed Heteromorphic Adaptation Strategy of Pyropia Haitanensis (Bangiales, Rhodophyta). PLoS ONE 2014, 9, e94354. [Google Scholar] [CrossRef] [PubMed]
- Kamenarska, Z.; Ivanova, A.; Stancheva, R.; Stoyneva, M.; Stefanov, K.; Dimitrova-Konaklieva, S.; Popov, S. Volatile Compounds from Some Black Sea Red Algae and Their Chemotaxonomic Application. Bot. Mar. 2006, 49, 47–56. [Google Scholar] [CrossRef]
- The Good Scents Company TGSC Information System. Available online: http://www.thegoodscentscompany.com/ (accessed on 15 May 2023).
- Sugisawa, H.; Nakamura, K.; Tamura, H. The Aroma Profile of the Volatiles in Marine Green Algae (Ulva Pertusa). Food Rev. Int. 1990, 6, 573–589. [Google Scholar] [CrossRef]
- Kajiwara, T.; Matsui, K.; Akakabe, Y. Biogeneration of Volatile Compounds via Oxylipins in Edible Seaweeds. In Biotechnology for Improved Foods and Flavors; American Chemical Society: Washington, DC, USA, 1996; pp. 146–166. [Google Scholar] [CrossRef]
- Akakabe, Y.; Matsui, K.; Kajiwara, T. 2,4-Decadienals Are Produced via (R)-11-HPITE from Arachidonic Acid in Marine Green Alga Ulva Conglobata . Bioorganic Med. Chem. 2003, 11, 3607–3609. [Google Scholar] [CrossRef]
- Wu, T.; Wang, M.; Wang, P.; Tian, H.; Zhan, P. Advances in the Formation and Control Methods of Undesirable Flavors in Fish. Foods 2022, 11, 2504. [Google Scholar] [CrossRef]
- Pan, X.; Ushio, H.; Ohshima, T. Photo-Oxidation of Lipids Impregnated on the Surface of Dried Seaweed (Porphyra Yezoensis Ueda). Characterization of Volatiles. J. Am. Oil Chem. Soc. 2004, 81, 773. [Google Scholar] [CrossRef]
- Vieira, K.R.; Pinheiro, P.N.; Zepka, L.Q. Volatile Organic Compounds from Microalgae. In Handbook of Microalgae-Based Processes and Products; Elsevier: Amsterdam, The Netherlands, 2020; pp. 659–686. ISBN 9780128185360. [Google Scholar]
- Xu, Y.X.; Jiang, Z.D.; Du, X.P.; Zheng, M.J.; Fan-Yang, Y.; Ni, H.; Chen, F. The Identification of Biotransformation Pathways for Removing Fishy Malodor from Bangia Fusco-Purpurea Using Fermentation with Saccharomyces Cerevisiae. Food Chem. 2022, 380, 132103. [Google Scholar] [CrossRef]
- Sun, S.-M.; Chung, G.; Shin, T.-S. Volatile Compounds of the Green Alga, Capsosiphon Fulvescens . J. Appl. Phycol. 2012, 24, 1003–1013. [Google Scholar] [CrossRef]
- Flodin, C.; Whitfield, F.B. Biosynthesis of Bromophenols in Marine Algae. Water Sci. Technol. 1999, 40, 53–58. [Google Scholar] [CrossRef]
- Hidalgo, F.J.; Zamora, R. Formation of Phenylacetic Acid and Benzaldehyde by Degradation of Phenylalanine in the Presence of Lipid Hydroperoxides: New Routes in the Amino Acid Degradation Pathways Initiated by Lipid Oxidation Products. Food Chem. X 2019, 2, 100037. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Chen, J.; Chen, L.; Shi, L.; Liu, H. Characteristic Volatile Composition of Seven Seaweeds from the Yellow Sea of China. Mar. Drugs 2021, 19, 192. [Google Scholar] [CrossRef] [PubMed]
- Akakabe, Y.; Iwamoto, S.; Miyamura, S.; Kajiwara, T. Induction of (Z)-8-Heptadecene in Marine Green Alga Bryopsis Maxima by Mechanical Wounding. Fish. Sci. 2007, 73, 1057–1060. [Google Scholar] [CrossRef]
- Stévant, P.; Ólafsdóttir, A.; Déléris, P.; Dumay, J.; Fleurence, J.; Ingadóttir, B.; Jónsdóttir, R.; Ragueneau, É.; Rebours, C.; Rustad, T. Semi-Dry Storage as a Maturation Process for Improving the Sensory Characteristics of the Edible Red Seaweed Dulse (Palmaria Palmata). Algal Res. 2020, 51, 102048. [Google Scholar] [CrossRef]
- Mirzayeva, A.; Castro, R.; Barroso, C.G.; Durán-Guerrero, E. Characterization and Differentiation of Seaweeds on the Basis of Their Volatile Composition. Food Chem. 2021, 336, 127725. [Google Scholar] [CrossRef]
- Hung, Y.-H.R.; Peng, C.-Y.; Huang, M.-Y.; Lu, W.-J.; Lin, H.-J.; Hsu, C.-L.; Fang, M.-C.; Lin, H.-T.V. Monitoring the Aroma Compound Profiles in the Microbial Fermentation of Seaweeds and Their Effects on Sensory Perception. Fermentation 2023, 9, 135. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| VOCs | Days | Porphyra umbilicalis | ||
|---|---|---|---|---|
| CTRL | MAP | VAC | ||
| 8-Heptadecene (isomer 2) CAS# N/D  Threshold: N/D | Day 0 | 10.878 ± 4.982 | — | — |
| Day 3 | 7.346 ± 0.801 | 19.952 ± 1.836 | 28.141 ± 2.848 | |
| Day 6 | 12.577 ± 1.852 | 24.026 ± 3.459 | 29.272 ± 1.133 | |
| Day 9 | 10.939 ± 1.969 | 23.277 ± 2.356 | 22.992 ± 1.849 | |
| Day 12 | 12.219 ± 0.368 | 25.666 ± 1.829 | 32.619 ± 3.041 | |
| Day 15 | 14.501 ± 1.913 | 27.679 ± 3.405 | 5.527 ± 1.823 | |
| Heptadecane CAS# 629-78-7  Threshold: N/D | Day 0 | 3.037 ± 1.524 | — | — |
| Day 3 | 0.924 ± 0.024 | 7.098 ± 0.955 | 6.817 ± 0.667 | |
| Day 6 | 1.109 ± 0.058 | 7.92 ± 1.481 | 8.552 ± 0.732 | |
| Day 9 | 0.961 ± 0.290 | 6.77 ± 0.710 | 6.648 ± 0.185 | |
| Day 12 | 1.192 ± 0.048 | 7.41 ± 0.338 | 7.271 ± 0.559 | |
| Day 15 | 2.037 ± 0.262 | 10.283 ± 0.975 | 1.143 ± 0.418 | |
| Pentadecanal CAS# 2765-11-9  Threshold: 0.715 mg/kg | Day 0 | 1.826 ± 0.464 | — | — |
| Day 3 | 17.544 ± 0.702 | 8.747 ± 0.709 | 0.695 ± 0.011 | |
| Day 6 | 10.309 ± 0.124 | 25.939 ± 3.018 | 1.529 ± 0.689 | |
| Day 9 | 12.627 ± 3.127 | 25.229 ± 2.091 | 1.993 ± 0.531 | |
| Day 12 | 23.097 ± 0.329 | 19.565 ± 1.615 | 2.827 ± 0.132 | |
| Day 15 | 12.968 ± 1.836 | 11.103 ± 0.539 | 1.732 ± 0.549 | |
| VOCs | Days | Ulva lactuca | ||
|---|---|---|---|---|
| CTRL | MAP | VAC | ||
| 2,4-Decadienal (isomer) CAS# 25152-84-5 (?)  Threshold: 0.00013 mg/kg | Day 0 | 27.565 ± 0.082 | — | — |
| Day 3 | 9.401 ± 0.596 | 8.748 ± 0.206 | 10.866 ± 1.603 | |
| Day 6 | 2.166 ± 0.003 | 9.163 ± 0.299 | 9.387 ± 1.902 | |
| Day 9 | 0.695 ± 0.105 | 9.796 ± 0.081 | 2.521 ± 0.002 | |
| Day 12 | 0.506 ± 0.021 | 8.731 ± 0.827 | 6.464 ± 0.024 | |
| Day 15 | 0.810 ± 0.163 | 4.250 ± 0.712 | 6.128 ± 0.142 | |
| 2,4-Decadienal, (E,E)- CAS# 2363-88-4  Threshold: 0.0003 mg/kg | Day 0 | 12.425 ± 0.412 | — | — |
| Day 3 | 4.018 ± 0.248 | 8.110 ± 1.208 | 6.853 ± 0.463 | |
| Day 6 | 1.775 ± 0.022 | 12.590 ± 0.687 | 8.237 ± 0.381 | |
| Day 9 | 0.701 ± 0.118 | 12.584 ± 0.104 | 5.377 ± 0.004 | |
| Day 12 | 0.950 ± 0.027 | 20.548 ± 1.950 | 10.449 ± 0.038 | |
| Day 15 | 1.210 ± 0.253 | 12.312 ± 1.823 | 11.744 ± 1.109 | |
| 1-Hexanol CAS# 111-27-3  Threshold: 1.21 mg/kg | Day 0 | 0.133 ± 0.025 | — | — |
| Day 3 | 6.025 ± 0.050 | 0.856 ± 0.053 | 0.504 ± 0.029 | |
| Day 6 | 14.377 ± 1.316 | 1.074 ± 0.070 | 0.611 ± 0.011 | |
| Day 9 | 17.684 ± 0.261 | 0.443 ± 0.004 | 0.879 ± 0.001 | |
| Day 12 | 24.474 ± 0.584 | 1.102 ± 0.103 | 0.850 ± 0.003 | |
| Day 15 | 4.670 ± 0.477 | 0.828 ± 0.030 | 1.594 ± 0.176 | |
| Benzaldehyde CAS# 100-52-7  Threshold: 0.55 mg/kg | Day 0 | 2.287 ± 0.027 | — | — |
| Day 3 | 2.939 ± 0.571 | 4.004 ± 0.207 | 5.146 ± 0.094 | |
| Day 6 | 5.586 ± 2.270 | 4.136 ± 0.325 | 3.371 ± 1.052 | |
| Day 9 | 3.804 ± 0.431 | 8.441 ± 0.070 | 15.221 ± 0.010 | |
| Day 12 | 6.023 ± 1.384 | 11.037 ± 0.812 | 17.494 ± 0.064 | |
| Day 15 | 9.642 ± 0.367 | 33.800 ± 9.784 | 11.8 ± 2.147 | |
| 8-Heptadecene (isomer 1) CAS# N/D  Threshold: N/D | Day 0 | 6.000 ± 1.107 | — | — |
| Day 3 | 13.026 ± 0.003 | 12.674 ± 1.198 | 13.193 ± 2.260 | |
| Day 6 | 15.353 ± 1.827 | 16.715 ± 0.625 | 15.777 ± 4.864 | |
| Day 9 | 16.710 ± 0.912 | 19.334 ± 0.160 | 16.086 ± 0.011 | |
| Day 12 | 6.424 ± 0.752 | 12.419 ± 1.177 | 8.505 ± 0.031 | |
| Day 15 | 8.799 ± 0.376 | 7.026 ± 2.471 | 9.521 ± 0.380 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreira-Leite, B.; Antunes, R.; Cotas, J.; Martins, N.; Costa, N.; Noronha, J.P.; Mata, P.; Diniz, M. Modified Atmosphere Packaging (MAP) for Seaweed Conservation: Impact on Physicochemical Characteristics and Microbiological Activity. Foods 2023, 12, 2736. https://doi.org/10.3390/foods12142736
Moreira-Leite B, Antunes R, Cotas J, Martins N, Costa N, Noronha JP, Mata P, Diniz M. Modified Atmosphere Packaging (MAP) for Seaweed Conservation: Impact on Physicochemical Characteristics and Microbiological Activity. Foods. 2023; 12(14):2736. https://doi.org/10.3390/foods12142736
Chicago/Turabian StyleMoreira-Leite, Bruno, Rafael Antunes, João Cotas, Nuno Martins, Nuno Costa, João P. Noronha, Paulina Mata, and Mário Diniz. 2023. "Modified Atmosphere Packaging (MAP) for Seaweed Conservation: Impact on Physicochemical Characteristics and Microbiological Activity" Foods 12, no. 14: 2736. https://doi.org/10.3390/foods12142736
APA StyleMoreira-Leite, B., Antunes, R., Cotas, J., Martins, N., Costa, N., Noronha, J. P., Mata, P., & Diniz, M. (2023). Modified Atmosphere Packaging (MAP) for Seaweed Conservation: Impact on Physicochemical Characteristics and Microbiological Activity. Foods, 12(14), 2736. https://doi.org/10.3390/foods12142736