


Efficiency of Lactiplantibacillus plantarum JT-PN39 and Paenibacillus motobuensis JT-A29 for Fermented Coffee Applications and Fermented Coffee Characteristics
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolation and Screening for Enzyme Production
2.2. Bacterial Identification
2.2.1. Genomics DNA Preparation and PCR Amplification
2.2.2. DNA Sequencing and Phylogenetic Analysis
2.3. Examination of Coffee Digestion
2.4. Mini-Scale Coffee Fermentation
2.5. Sensory Analysis Based on SCAA Cupping Test
2.6. Coffee Extract Preparation and Biological Property Examinations
2.6.1. Total Phenolic Compounds and Antioxidant Activity
2.6.2. Antibacterial Activity
2.7. Qualitative Examination of Fermented Coffee Properties by LC-MS and GC-MS
2.7.1. Non-Volatile Compounds
2.7.2. Volatile Compounds
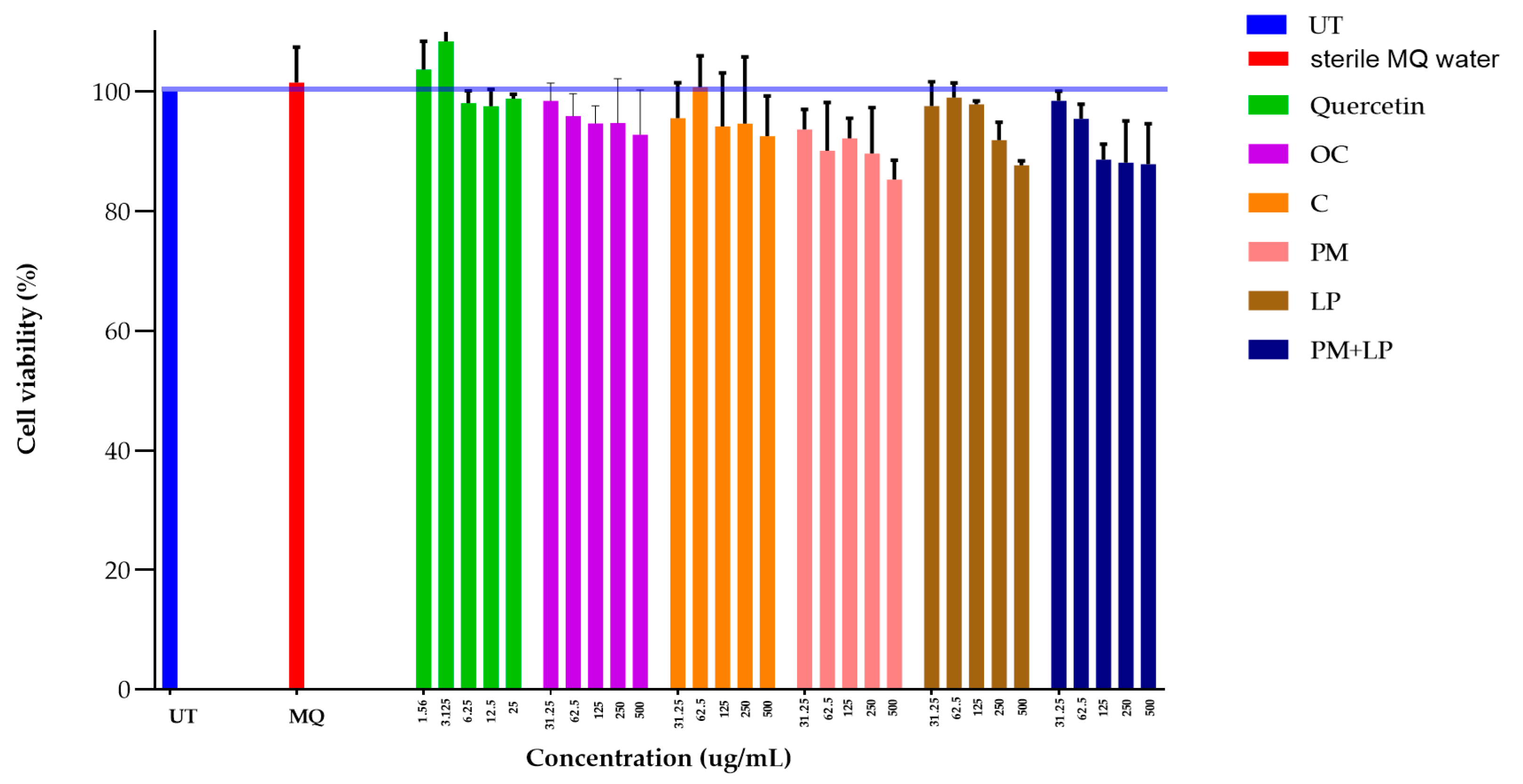
2.8. Toxicity Test in RAW 264.7 Cell
2.9. Statistical Analysis
3. Results
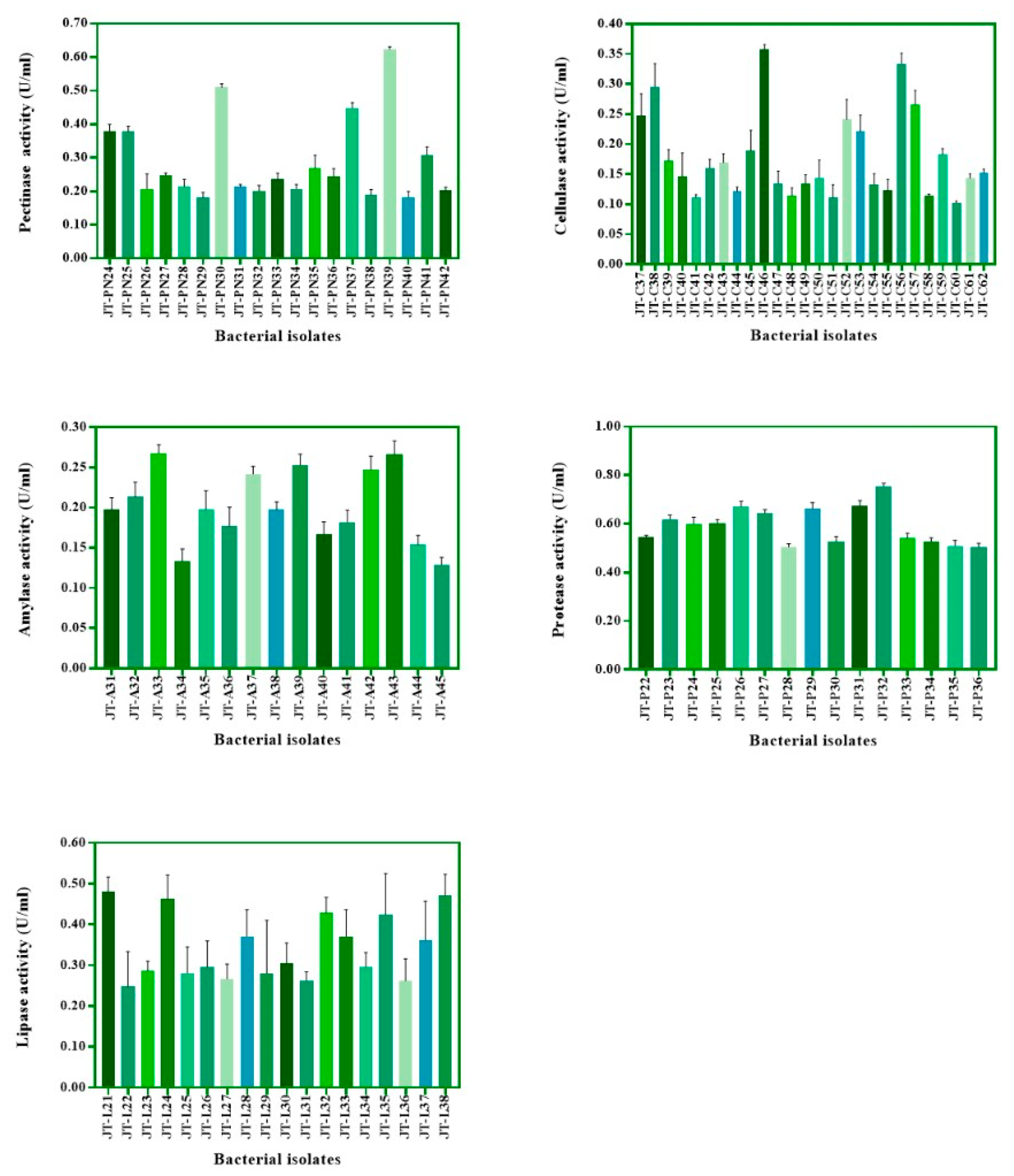
3.1. Isolation and Screening of Bacteria Based on Enzyme Activity
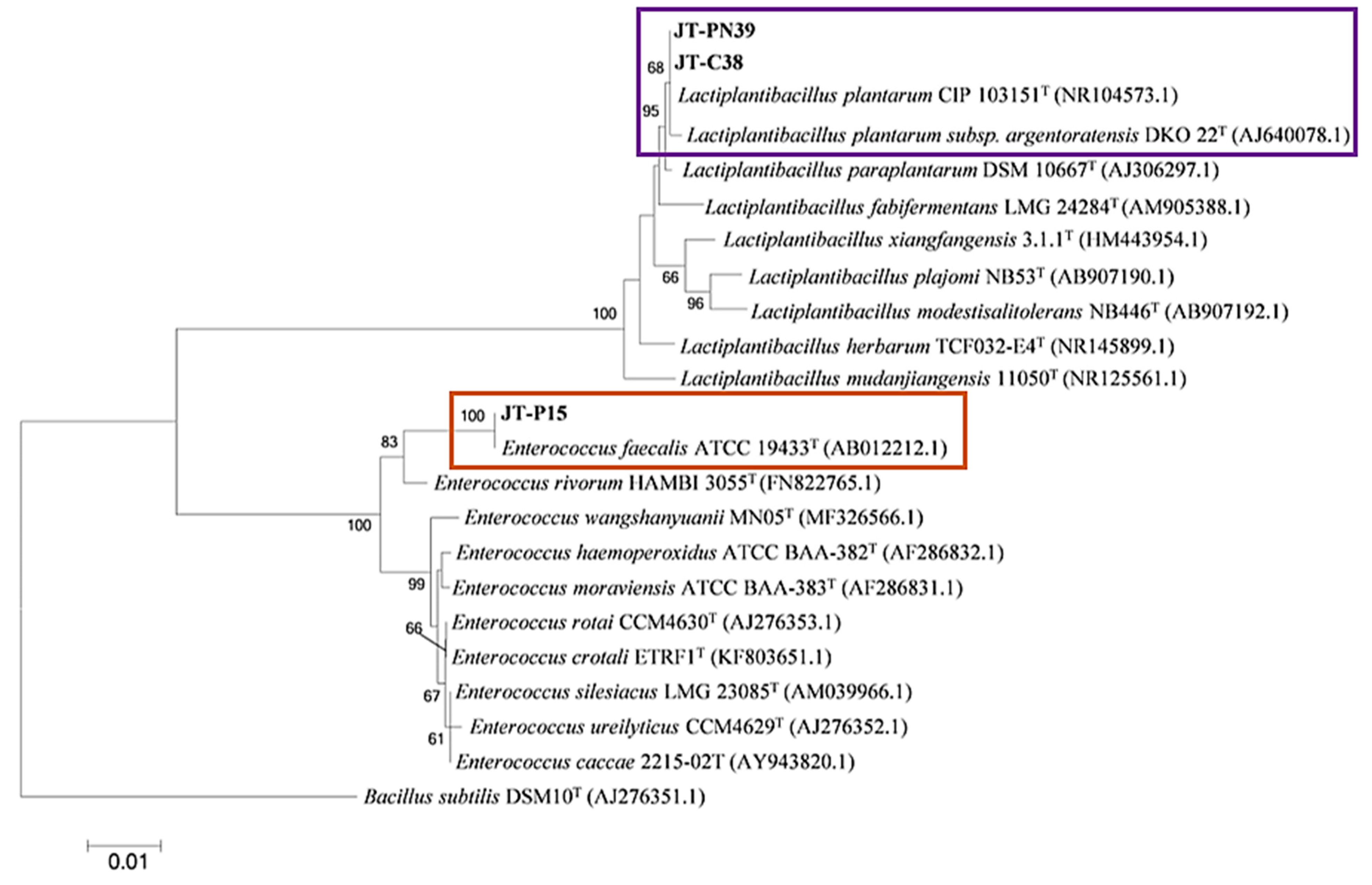
3.2. Identification of Bacteria Producing Enzymes
3.3. Parchment Coffee Digetion by Selected Bacteria
3.4. Coffee Fermentation
3.4.1. Bacterial Growth and pH
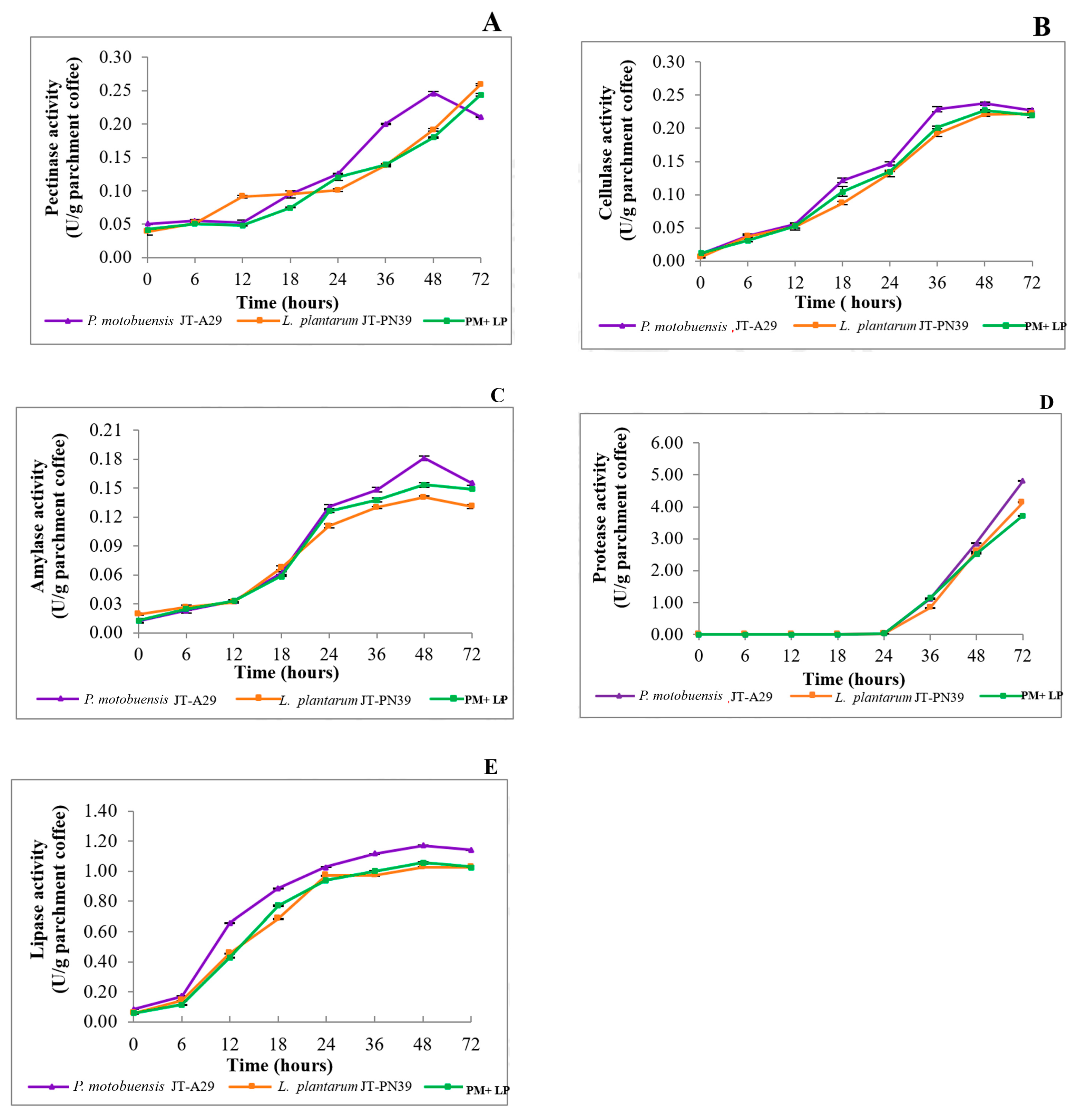
3.4.2. Enzyme Production during Coffee Fermentation
3.5. Sensory Evaluation
3.6. Biological Functions of Roasted Fermented Coffee
3.6.1. Total Phenolic Content and Antioxidants
3.6.2. Antipathogenic Bacterial Activity
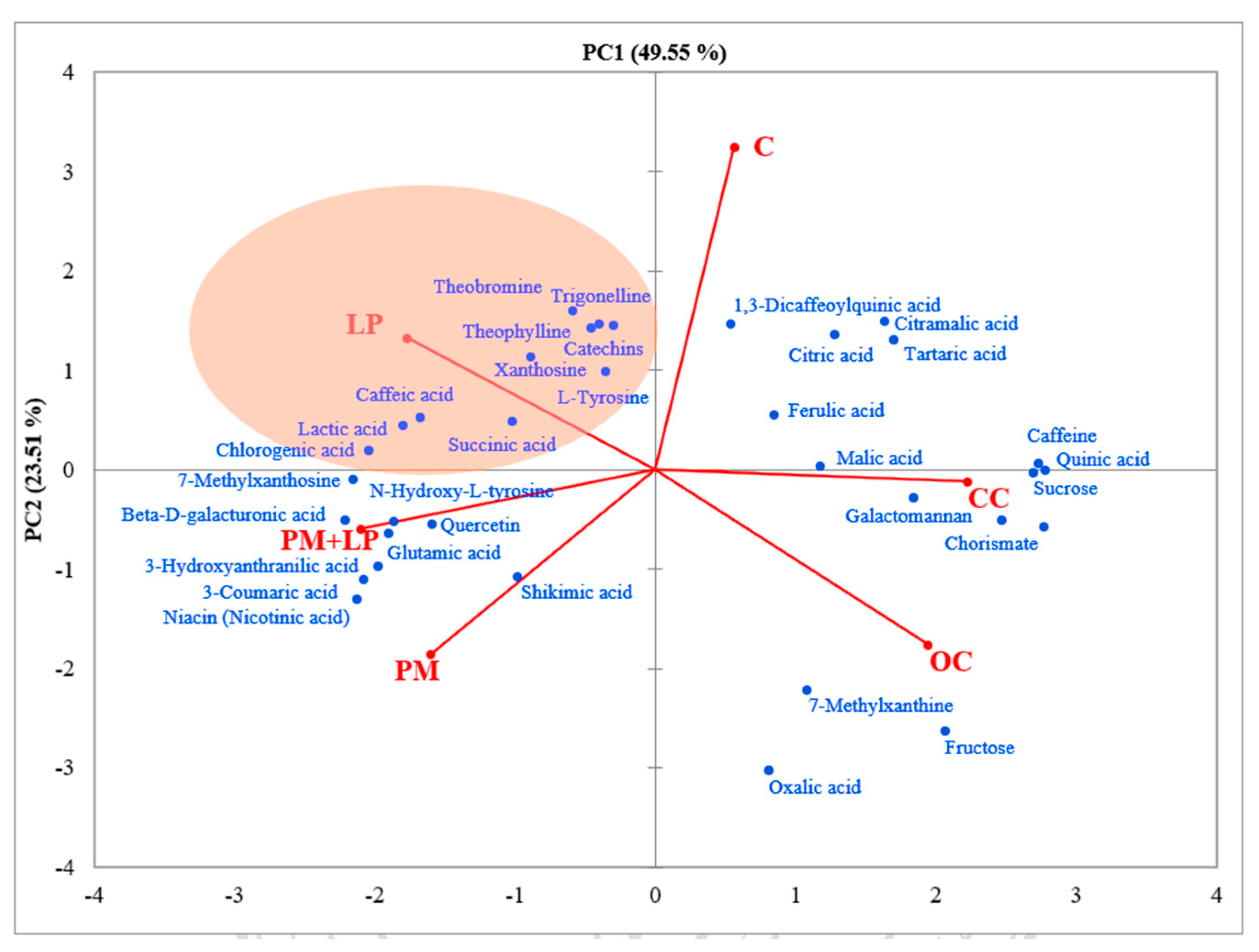
3.7. Non-Volatile and Volatile Compound Qualification in Fermented Coffee
3.8. Toxicity of Fermented Coffee
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ademiluyi, A.O.; Oboh, G. Antioxidant properties of condiment produced from fermented bambara groundnut (Vigna subterranea L. verdc). J. Food Biochem. 2011, 35, 1145–1160. [Google Scholar] [CrossRef]
- Andrade, C.; Perestrelo, R.; Câmara, J.S. Valorization of spent coffee grounds as a natural source of bioactive compounds for several industrial applications—A volatilomic approach. Foods 2022, 11, 1731. [Google Scholar] [CrossRef]
- Andreou, V.; Strati, I.F.; Fotakis, C.; Liouni, M.; Zoumpoulakis, P.; Sinanoglou, V.J. Herbal Distillates: A new era of grape marc distillates with enriched antioxidant profile. Food Chem. 2018, 253, 171–178. [Google Scholar] [CrossRef]
- Avallone, S.; Guyot, B.; Brillouet, J.; Olguin, E.; Guiraud, J. Microbiological and biochemical study of coffee fermentation. Curr. Microbiol. 2001, 42, 252–256. [Google Scholar] [CrossRef]
- Bastian, F.; Hutabarat, O.S.; Dirpan, A.; Nainu, F.; Harapan, H.; Emran, T.B.; Simal-Gandara, J. From plantation to cup: Changes in bioactive compounds during coffee processing. Foods 2021, 10, 2827. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho Neto, D.P.; De Melo Pereira, G.V.; Tanobe, V.O.A.; Thomaz Soccol, V.G.; da Silva, B.J.; Rodrigues, C.; Soccol, C.R. Yeast Diversity and Physicochemical Characteristics Associated with Coffee Bean Fermentation from the Brazilian Cerrado Mineiro Region. Fermentation 2017, 3, 11. [Google Scholar] [CrossRef] [Green Version]
- de Carvalho Neto, D.P.; Gilberto, V.; de Melo Pereira, G.V.; Finco, A.M.O.; Letti, L.A.J.; Silva, B.J.G.; Vandenberghe, L.P.S.; Soccol, C.R. Efficient coffee beans mucilage layer removal using lactic acid fermentation in a stirred-tank bioreactor: Kinetic, metabolic and sensorial studies. Food Biosci. 2018, 26, 80–87. [Google Scholar] [CrossRef]
- Chenna, R.; Sugawara, H.; Koike, T.; Lopez, R.; Gibson, T.J.; Higgins, D.G.; Thompson, J.D. Multiple sequence alignment with the Clustal series of programs. Nucleic Acids Res. 2003, 31, 3497–3500. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standards Institute. Methods for Antimicrobial Susceptibility Testing for Human Mycoplasmas. In Approved Guideline; CLSI Document M43-A; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2011. [Google Scholar]
- Duangjai, A.; Suphrom, N.; Wungrath, J.; Ontawong, A.; Nuengchamnong, N.; Yosboonruang, A. Comparison of antioxidant, antimicrobial activities and chemical profiles of three coffee (Coffea arabica L.) pulp aqueous extracts. Integ. Med. Res. 2016, 5, 324–331. [Google Scholar] [CrossRef] [Green Version]
- Elavarasan, K.; Kumar, A.; Manoharan, A.; Rajan, S.S. The basics of coffee cupping (COFFEE CUPPING). Tea Coffee Trade J. 2016, 188, 30. [Google Scholar]
- Esquivel, P.; Jiménez, V.M. Functional properties of coffee and coffee by-products. Food Res. Inter. 2012, 46, 488–495. [Google Scholar] [CrossRef]
- Farah, A.; Monteiro, M.; Donangelo, C.M.; Lafay, S. Chlorogenic acids from green coffee extract are highly bioavailable in humans. J. Nutr. 2008, 138, 2309–2315. [Google Scholar] [CrossRef] [Green Version]
- García-Díez, J.; Saraiva, C. Use of starter cultures in foods from animal origin to improve their safety. Int. J. Environ. Res. Public Health 2021, 18, 2544. [Google Scholar] [CrossRef]
- Gökcen, B.B.; Şanlier, N. Coffee consumption and disease correlations. J. Food Sci. Nutri. 2019, 59, 336–348. [Google Scholar] [CrossRef]
- Gonzalez-Rios, O.; Suarez-Quiroz, M.L.; Boulanger, R.; Barel, M.; Guyot, B.; Guiraud, J.-P.; Schorr-Galindo, S. Impact of “ecological” post-harvest processing on the volatile fraction of coffee beans: I. Green coffee. J. Food Compos. Anal. 2007, 20, 289–296. [Google Scholar] [CrossRef]
- Gupta, N.; Rathi, P.; Gupta, R. Simplified para-nitrophenyl palmitate assay for lipases and esterases. Anal. Biochem. 2002, 311, 98–99. [Google Scholar] [CrossRef] [PubMed]
- Haile, M.; Kang, W.H. Antioxidant properties of fermented green coffee beans with Wickerhamomyces anomalus (strain KNU18Y3). Fermentation 2020, 6, 18. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nuc. Acids Sym. Ser. 1999, 41, 95–98. [Google Scholar]
- Harwood, C.R.; Kikuchi, Y. The ins and outs of Bacillus proteases: Activities, functions and commercial significance. FEMS Microbiol. Rev. 2022, 46, fuab046. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Jain, R.; Jain, S. Quantitative analysis of reducing sugars by 3, 5-Dinitrosalicylic acid (DNSA Method). In Basic Techniques in Biochemistry, Microbiology and Molecular Biology; Springer Protocols Handbooks: Humana, NY, USA, 2020; pp. 181–183. [Google Scholar] [CrossRef]
- Kamiyama, M.; Moon, J.K.; Jang, H.W.; Shibamoto, T. Role of degradation products of chlorogenic acid in the antioxidant activity of roasted coffee. J. Agric. Food Chem. 2015, 63, 1996–2005. [Google Scholar] [CrossRef] [PubMed]
- Kim, O.S.; Cho, Y.J.; Lee, K.; Yoon, S.H.; Kim, M.; Na, H.; Park, S.C.; Jeon, Y.S.; Lee, J.H.; Yi, H.; et al. Introducing EzTaxon-e: A prokaryotic 16S rRNA gene sequence database with phylotypes that represent uncultured species. Int. J. Syst. Evol. Microbiol. 2012, 62, 716–721. [Google Scholar] [CrossRef] [Green Version]
- Küçükgöz, K.; Trząskowska, M. Nondairy Probiotic Products: Functional Foods That Require More Attention. Nutrients 2022, 14, 753. [Google Scholar] [CrossRef]
- Kumari, A.; Kaur, B.; Srivastava, R.; Sangwan, S. Isolation and immobilization of alkaline protease on mesoporous silica and mesoporous ZSM-5 zeolite materials for improved catalytic properties. Biochem. Biophys Rep. 2015, 2, 108–114. [Google Scholar] [CrossRef] [Green Version]
- Kwak, H.S.; Jeong, Y.; Kim, M. Effect of yeast fermentation of green coffee beans on antioxidant activity and consumer acceptability. J. Food Qual. 2008, 2018, 8. [Google Scholar] [CrossRef] [Green Version]
- Leska, A.; Nowak, A.; Motyl, I. Isolation and some basic characteristics of lactic acid bacteria from honeybee (Apis mellifera L.) Environment—A preliminary study. Agriculture 2022, 12, 1562. [Google Scholar] [CrossRef]
- Liang, N.; Kitts, D.D. Antioxidant property of coffee components: Assessment of methods that define mechanisms of action. Molecules 2014, 19, 19180–19208. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Shi, Y.; Wen, P.; Hu, X.; Wang, L. Free, Conjugated, and bound phenolics in peel and pulp from four wampee varieties: Relationship between phenolic composition and Bio-activities by multivariate analysis. Antioxidants 2022, 11, 1831. [Google Scholar] [CrossRef]
- Lingle, T.R. The Coffee Brewing Handbook: A Systematic Guide to Coffee Preparation, 2nd ed.; Specialty Coffee Association of America: Santa Ana, CA, USA, 2011. [Google Scholar]
- Meerak, J.; Yukphan, P.; Miyashita, M.; Sato, H.; Nakagawa, Y.; Tahara, Y. Phylogeny of γ-polyglutamic acid-producing Bacillus strains isolated from a fermented locust bean product manufactured in West Africa. J. Gen. Appl. Microbiol. 2008, 54, 159–166. [Google Scholar] [CrossRef] [Green Version]
- Mojica, B.E.; Fong, L.E.; Biju, D.; Muharram, A.; Davis, I.M.; Vela, K.O.; Forester, S.C. The impact of the roast levels of coffee extracts on their potential anticancer activities. J. Food Sci. 2018, 83, 1125–1130. [Google Scholar] [CrossRef]
- Monente, C.; Bravo, J.; Vitas, A.I.; Arbillaga, L.; De Peña, M.P.; Cid, C. Coffee and spent coffee extracts protect against cell mutagens and inhibit growth of food-borne pathogen microorganisms. J. Funct. Foods. 2015, 12, 365–374. [Google Scholar] [CrossRef]
- Moon, J.K.; Shibamoto, T. Role of roasting conditions in the profile of volatile flavor chemicals formed from coffee beans. J. Agric. Food Chem. 2009, 57, 5823–5831. [Google Scholar] [CrossRef]
- Moreira, A.S.P.; Nunes, F.M.; Domingues, M.R.; Coimbra, M.A. Coffee melanoidins: Structures, mechanisms of formation and potential health impacts. Food Funct. 2012, 3, 903–915. [Google Scholar] [CrossRef]
- Myo, H.; Nantarat, N.; Khat-Udomkiri, N. Changes in bioactive compounds of coffee pulp through fermentation-based biotransformation using Lactobacillus plantarum TISTR 543 and its antioxidant activities. Fermentation 2021, 7, 292. [Google Scholar] [CrossRef]
- Naczk, M.; Shahidi, F. Extraction and analysis of phenolics in food. J. Chromatogr. 2004, 1054, 95–111. [Google Scholar] [CrossRef]
- Novović, K.; Malešević, M.; Dinić, M.; Gardijan, L.; Kojić, M.; Jovčić, B. RclS Sensor Kinase. Modulates Virulence of Pseudomonas capeferrum. Int. J. Mol. Sci. 2022, 23, 8232. [Google Scholar] [CrossRef]
- de Oliveira Junqueira, A.C.; de Melo Pereira, G.V.; Coral Medina, J.D.; Alvear, M.C.R.; Rosero, R.; de Carvalho Neto, D.P.; Enríquez, H.G.; Soccol, C.R. First description of bacterial and fungal communities in Colombian coffee beans fermentation analysed using Illumina-based amplicon sequencing. Sci. Rep. 2019, 9, 8794. [Google Scholar] [CrossRef] [Green Version]
- Peñuela-Martínez, A.E.; Moreno-Riascos, S.; Medina-Rivera, R. Influence of temperature-controlled fermentation on the quality of mild coffee (Coffea arabica L.) cultivated at different elevations. Agriculture 2023, 13, 1132. [Google Scholar] [CrossRef]
- Pereira, G.V.; Neto, E.; Soccol, V.T.; Medeiros, A.B.P.; Woiciechowski, A.L.; Soccol, C.R. Conducting starter culture-controlled fermentations of coffee beans during on-farm wet processing: Growth, metabolic analyses and sensorial effects. Food Res. Int. 2015, 75, 348–356. [Google Scholar] [CrossRef]
- Pereira, G.V.M.; de Carvalho Neto, D.P.; Medeiros, A.B.P.; Soccol, V.T.; Neto, E.; Woiciechowski, A.L.; Soccol, C.R. Potential of lactic acid bacteria to improve the fermentation and quality of coffee during on-farm processing. Int. J. Food Sci. Technol. 2016, 51, 1689–1695. [Google Scholar] [CrossRef]
- Pregolini, V.B.; de Melo Pereira, G.V.; da Silva Vale, A.; de Carvalho Neto, D.P.; Soccol, C.R. Influence of Environmental Microbiota on the Activity and Metabolism of Starter Cultures Used in Coffee Beans Fermentation. Fermentation 2021, 7, 278. [Google Scholar] [CrossRef]
- Priftis, A.; Stagos, D.; Konstantinopoulos, K.; Tsitsimpikou, C.; Spandidos, D.A.; Tsatsakis, A.M.; Kouretas, D. Comparison of antioxidant activity between green and roasted coffee beans using molecular methods. Mol. Med. Rep. 2015, 12, 7293–7302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, L.S.; Evangelista, S.R.; da Cruz Pedrozo Miguel, M.G.; Mullem, J.; Silva, C.F.; Schwan, R.F. Microbiological and chemical-sensory characteristics of three coffee varieties processed by wet fermentation. Ann. Microbiol. 2018, 68, 705–716. [Google Scholar] [CrossRef]
- Runti, G.; Pacor, S.; Colomban, S.; Gennaro, R.; Navarini, L.; Scocchi, M. Arabica coffee extract shows antibacterial activity against Staphylococcus epidermidis and Enterococcus faecalis and low toxicity towards a human cell line. LWT Food Sci. Technol. 2015, 62, 108–114. [Google Scholar] [CrossRef] [Green Version]
- Ruta, L.L.; Farcasanu, I.C. Coffee and Yeasts: From flavor to biotechnology. Fermentation 2021, 7, 9. [Google Scholar] [CrossRef]
- Sahlany, A.I.; Khassaf, W.H.; Niamah, A.K.; Manhel, A.I. Date juice addition to bio-yogurt: The effects on physicochemical and microbiological properties during storage, as well as blood parameters in vivo. J. Saudi Soc. Agric. Sci. 2023, 22, 71–77. [Google Scholar] [CrossRef]
- Seitzer, P.; Bennett, B.; Melamud, E. MAVEN2: An Updated Open-Source Mass Spectrometry Exploration Platform. Metabolites 2022, 12, 684. [Google Scholar] [CrossRef]
- Shrestha, A.K.; Dahal, N.R.; Ndungutse, V. Bacillus fermentation of soybean: A review. J. Food Sci. Technol. Nepal. 2013, 6, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Smrke, S.; Opitz, S.E.W.; Vovk, I.; Yeretzian, C. How does roasting affect the antioxidants of a coffee brew? Exploring the antioxidant capacity of coffee via on-line antioxidant assays coupled with size exclusion chromatography. Food Funct. 2013, 4, 1082–1092. [Google Scholar] [CrossRef]
- Specialty Coffee Association of America. SCAA Standard. In Golden Cup; Specialty Coffee Association of America: Irvine, CA, USA, 2015. [Google Scholar]
- Sridharan, S.; Das, K.M.S. A study on suitable non dairy food matrix for probiotic bacteria—A systematic review. Curr. Res. Nutr. Food Sci. 2019, 7, 5–16. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [Green Version]
- Trugo, L.C.; Macrae, R. A study of the effect of roasting on the chlorogenic acid composition of coffee using HPLC. Food Chem. 1984, 15, 219–227. [Google Scholar] [CrossRef]
- Vaast, P.; Bertrand, B.; Perriot, J.-J.; Guyot, B.; Génard, M. Fruit thinning and shade improve bean characteristics and beverage quality of coffee (Coffea arabica L.) under optimal conditions. J. Sci. Food Agric. 2006, 86, 197–204. [Google Scholar] [CrossRef]
- Wang, Y.-J.; Zhou, S.-M.; Xu, G.; Gao, Y.-Q. Interference of Phenylethanoid Glycosides from Cistanche tubulosa with the MTT Assay. Molecules 2015, 20, 8060–8071. [Google Scholar] [CrossRef] [Green Version]
- Wright, A.V. Genus Lactococcus. In Lactic acid Bacteria: Microbiological and Functional Aspects; Lahtinen, S., Ouwehand, A.C., Salminen, S., Wright, A.V., Eds.; Marcel Dekker: New York, NY, USA, 2012; pp. 63–76. [Google Scholar]
- Zhao, L.; Wang, Y.; Wang, D.; He, Z.; Gong, J.; Tan, C. Effects of Different Probiotics on the Volatile Components of Fermented Coffee Were Analyzed Based on Headspace-Gas Chromatography-Ion Mobility Spectrometry. Foods 2023, 12, 2015. [Google Scholar] [CrossRef]





 ), JT-PN39
(
), JT-PN39
( ), JT-C38 (
), JT-C38 ( ), JT-P29 (
), JT-P29 ( ), and JT-P32 (
), and JT-P32 ( ) from 0 to 192 h.
), JT-PN39
(), JT-C38 (), JT-P29 (), and JT-P32 () from 0 to 192 h.
) from 0 to 192 h.
), JT-PN39
(), JT-C38 (), JT-P29 (), and JT-P32 () from 0 to 192 h.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolates | Cellulase | Pectinase | Amylase | Protease | Lipase |
|---|---|---|---|---|---|
| JT-A29 | + | + | + | + | + |
| JT-C38 | + | + | − | − | + |
| JT-PN39 | + | + | + | + | + |
| JT-A43 | + | + | + | − | − |
| JT-C14 | + | + | − | − | − |
| JT-L19 | − | − | + | + | + |
| JT-P15 | − | − | + | + | − |
| JT-PN06 | − | + | + | + | + |
| JT-PN16 | − | + | − | − | + |
| JT-P29 | + | + | + | + | + |
| JT-P32 | + | + | + | + | + |
| Attributes and Varieties | Coffee | |||||
|---|---|---|---|---|---|---|
| OC | C | PM | LP | PM + LP | CC | |
| Fragrance/Aroma | 7.00 | 7.00 | 7.00 | 7.25 | 7.00 | 7.00 |
| Acidity | 7.00 | 7.00 | 7.00 | 7.25 | 7.25 | 7.00 |
| Flavor | 7.00 | 7.00 | 7.00 | 7.25 | 7.00 | 7.00 |
| Body | 7.00 | 7.00 | 6.75 | 7.25 | 6.75 | 7.00 |
| Aftertaste | 7.00 | 7.00 | 7.00 | 7.00 | 7.00 | 7.00 |
| Uniformity | 10.00 | 10.00 | 10.00 | 10.00 | 10.00 | 10.00 |
| Balance | 7.00 | 6.25 | 7.00 | 7.25 | 7.00 | 7.00 |
| Clean cup | 10.00 | 10.00 | 10.00 | 10.00 | 10.00 | 10.00 |
| Sweetness | 10.00 | 10.00 | 10.00 | 10.00 | 10.00 | 10.00 |
| Overall | 7.00 | 7.00 | 6.75 | 7.00 | 7.25 | 7.00 |
| Total score | 79.00 | 78.25 | 78.50 | 80.25 | 79.25 | 79.00 |
| Coffee | Aroma | |
|---|---|---|
| Dry Aroma | Wet Aroma | |
| OC | roasted, dark chocolate, cocoa nibs roasted | meat, caramel, brown sugar, smoke, hint of jackfruit |
| C | mozzarella cheese, spicy | cheese, cherry, mint, hint of lemon |
| PM | musty, nutty, dry wood, hint cheese | Musty, ferment, peach, hint of floral, hint cheese |
| LP | tobacco, cigarette, dry wood | caramel, herb, fresh leaf, flower, citrus, yoghurt |
| PM + LP | tobacco, dry wood, spicy | spicy, tea, chocolate, butter, seaweed |
| CC | rose, pinecone seed, dry wood | brown sugar, caramel, spicy, dry wood |
| Coffee Extracts | Antioxidant Activity | |
|---|---|---|
| ABTS Assay IC50 (mg/mL) | TPC (mg GAE/g Coffee) | |
| Roasted Coffee Beans | ||
| Ordinary coffee (OC) | 1.046 ± 0.006 de | 79.72 ± 0.54 de |
| Non-fermented (C) | 1.068 ± 0.008 f | 78.50 ± 0.72 f |
| PM | 0.999 ± 0.006 a | 84.42 ± 0.51 a |
| LP | 1.000 ± 0.002 a | 83.95 ± 0.47 a |
| PM + LP | 1.019 ± 0.017 bc | 82.70 ± 0.78 b |
| Civet coffee (CC) | 1.072 ± 0.006 fg | 78.38 ± 0.71 f |
| Civet coffee commercial | 1.104 ± 0.018 ij | 76.19 ± 0.31 h |
| Commercial Coffee 1 | 1.117 ± 0.010j | 68.50 ± 0.67 i |
| Commercial Coffee 2 | 1.005 ± 0.006 ab | 80.07 ± 0.73 de |
| Commercial Coffee 3 | 1.024 ± 0.009 c | 79.95 ± 0.35 de |
| Green coffee bean | ||
| Ordinary coffee (OC) | 1.060 ± 0.010 ef | 79.33 ± 0.71 def |
| Non-fermented (C) | 1.087 ± 0.002 gh | 79.13 ± 0.42 ef |
| PM | 1.058 ± 0.008 ef | 80.23 ± 0.30 d |
| LP | 1.041 ± 0.004 d | 81.87 ± 0.36 bc |
| PM + LP | 1.059 ± 0.009 ef | 81.64 ± 0.18 c |
| Civet coffee (CC) | 1.097 ± 0.007 hi | 77.33 ± 0.76 g |
| Coffee Extract | Zone of Inhibition (mm) | ||||
|---|---|---|---|---|---|
| S. aureus | S. aureus (MRSA) | L. monocytogenes | E. coli | S. enteritidis | |
| Chloramphenicol | 19.50 ± 0.50 a | 7.00 ± 0.00 h | 24.44 ± 0.51 a | 15.89 ± 0.84 a | 20.56 ± 0.51 a |
| Roasted coffee beans | |||||
| Ordinary coffee (OC) | 11.89 ± 0.10 f | 10.11 ± 0.10 g | 13.00 ± 0.00 h | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Non-fermented (C) | 14.17 ± 0.17 d | 24.83 ± 0.44 bc | 12.78 ± 0.38 h | 0.00 ± 0.00 | 0.00 ± 0.00 |
| PM | 12.11 ± 0.10 f | 24.39 ± 0.25 c | 15.22 ± 0.19 d | 0.00 ± 0.00 | 0.00 ± 0.00 |
| LP | 19.00 ± 0.17 b | 29.72 ± 0.25 a | 16.50 ± 0.44 b | 10.78 ± 0.38 b | 10.89 ± 0.10 b |
| PM + LP | 18.06± 0.25 c | 25.56 ± 0.96 b | 15.89 ± 0.25 c | 10.50 ± 0.50 b | 10.78 ± 0.25 b |
| Civet coffee (CC) | 11.89 ± 0.35 f | 18.44 ± 0.38 f | 14.33 ± 0.33 ef | 10.22 ± 0.38 b | 10.00 ± 0.00 c |
| Commercial civet coffee | 13.28± 0.10 e | 22.83 ± 0.50 d | 14.56 ± 0.51 e | 9.17 ± 0.17 c | 9.33 ± 0.00 d |
| Commercial Coffee 1 | 11.33 ± 0.29 g | 21.72 ± 0.48 e | 12.78 ± 0.25 h | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Commercial Coffee 2 | 9.44 ± 0.51 h | 24.56 ± 0.38 c | 13.78 ± 0.25 fg | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Commercial Coffee 3 | 11.86 ± 0.13 f | 22.56 ± 0.38 d | 13.22 ± 0.19 gh | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Coffee Extracts | Bacteria | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| S. aureus | S. aureus (MRSA) | L. monocytogenes | E. coli | S. enteritidis | ||||||
| MIC (mg/mL) | MBC (mg/mL) | MIC (mg/mL) | MBC (mg/mL) | MIC (mg/mL) | MBC (mg/mL) | MIC (mg/mL) | MBC (mg/mL) | MIC (mg/mL) | MBC (mg/mL) | |
| Chloramphenicol | 0.004 | 0.007 | 0.004 | 0.007 | 0.004 | 0.007 | 0.004 | 0.007 | 0.004 | 0.007 |
| Roasted coffee beans | ||||||||||
| Ordinary coffee (OC) | 41.67 | 166.67 | 41.67 | 83.33 | 41.67 | 166.67 | ND | ND | ND | ND |
| Non-fermented (C) | 41.67 | 166.67 | 41.67 | 83.33 | 83.33 | >166.7 | ND | ND | ND | ND |
| PM | 41.67 | 166.67 | 41.67 | 83.33 | 41.67 | 166.67 | ND | ND | ND | ND |
| LP | 41.67 | 83.33 | 41.67 | 83.33 | 41.67 | 83.33 | 41.67 | 83.33 | 41.67 | 83.33 |
| PM + LP | 41.67 | 83.33 | 41.67 | 83.33 | 41.67 | 166.67 | 41.67 | 166.67 | 41.67 | 166.67 |
| Civet coffee (CC) | 83.33 | >166.7 | 41.67 | 83.33 | 41.67 | 166.67 | 166.67 | 166.67 | 166.67 | 166.67 |
| Commercial civet coffee | 41.67 | 166.67 | 41.67 | 166.67 | 41.67 | 166.67 | 166.67 | 166.67 | 166.67 | 166.67 |
| Commercial Coffee 1 | 41.67 | 166.67 | 41.67 | 166.67 | 41.67 | 166.67 | ND | ND | ND | ND |
| Commercial Coffee 2 | 83.33 | >166.7 | 41.67 | 166.67 | 41.67 | 166.67 | ND | ND | ND | ND |
| Commercial Coffee 3 | 83.33 | >166.7 | 41.67 | 83.33 | 83.33 | >166.7 | ND | ND | ND | ND |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ngamnok, T.; Nimlamool, W.; Amador-Noguez, D.; Palaga, T.; Meerak, J. Efficiency of Lactiplantibacillus plantarum JT-PN39 and Paenibacillus motobuensis JT-A29 for Fermented Coffee Applications and Fermented Coffee Characteristics. Foods 2023, 12, 2894. https://doi.org/10.3390/foods12152894
Ngamnok T, Nimlamool W, Amador-Noguez D, Palaga T, Meerak J. Efficiency of Lactiplantibacillus plantarum JT-PN39 and Paenibacillus motobuensis JT-A29 for Fermented Coffee Applications and Fermented Coffee Characteristics. Foods. 2023; 12(15):2894. https://doi.org/10.3390/foods12152894
Chicago/Turabian StyleNgamnok, Teerawat, Wutigri Nimlamool, Daniel Amador-Noguez, Tanapat Palaga, and Jomkhwan Meerak. 2023. "Efficiency of Lactiplantibacillus plantarum JT-PN39 and Paenibacillus motobuensis JT-A29 for Fermented Coffee Applications and Fermented Coffee Characteristics" Foods 12, no. 15: 2894. https://doi.org/10.3390/foods12152894
APA StyleNgamnok, T., Nimlamool, W., Amador-Noguez, D., Palaga, T., & Meerak, J. (2023). Efficiency of Lactiplantibacillus plantarum JT-PN39 and Paenibacillus motobuensis JT-A29 for Fermented Coffee Applications and Fermented Coffee Characteristics. Foods, 12(15), 2894. https://doi.org/10.3390/foods12152894