

Multivariate Profiling of Metabolites and Volatile Organic Compounds in Citrus depressa Hayata Fruits from Kagoshima, Okinawa, and Taiwan

 , ,
, ,  , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Standards and Reagents
2.3. Metabolite Profiling Analysis
2.4. VOC Analysis
2.5. Statistical Analysis
3. Results
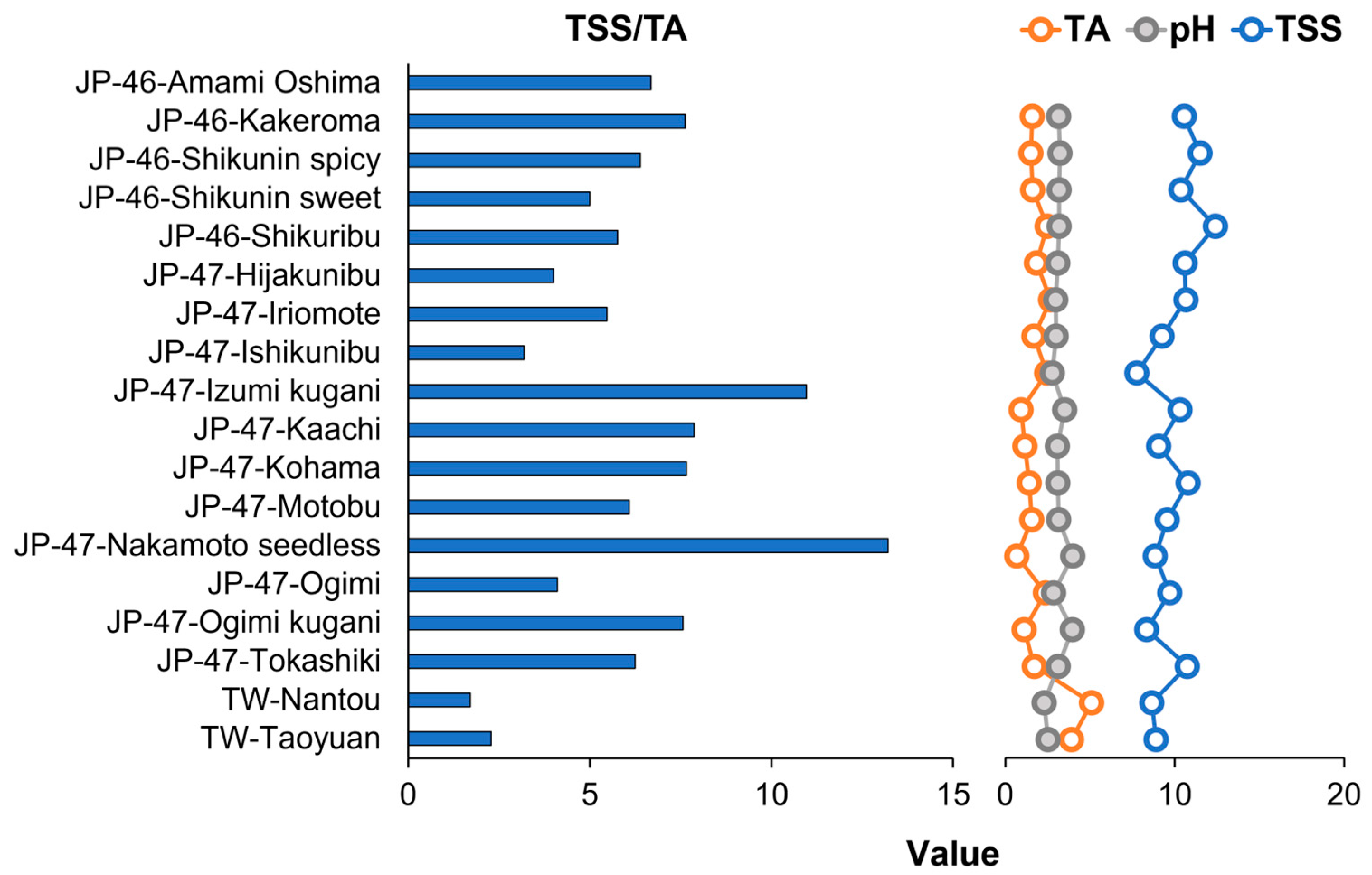
3.1. Physicochemical Properties of C. depressa Fruits
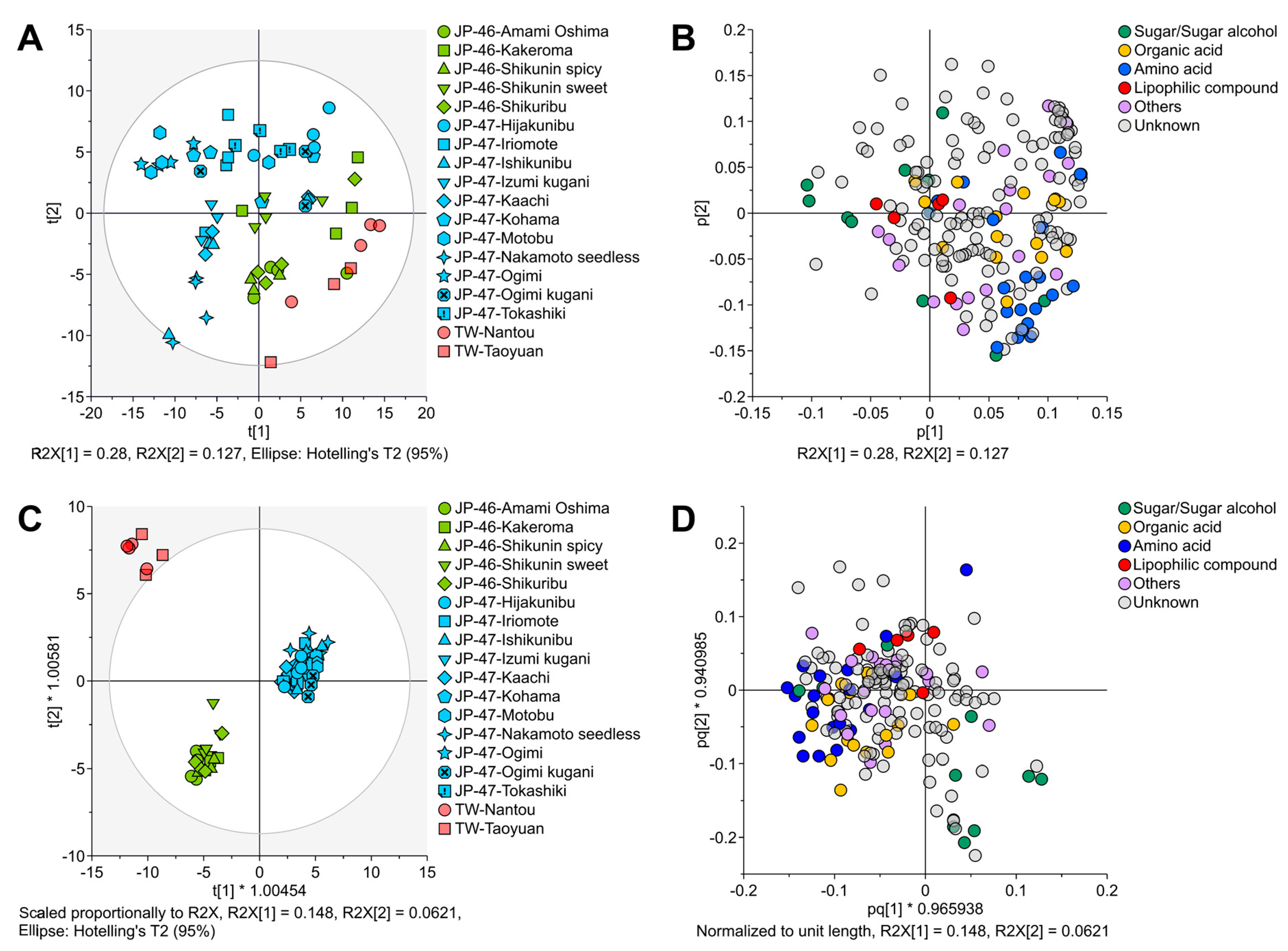
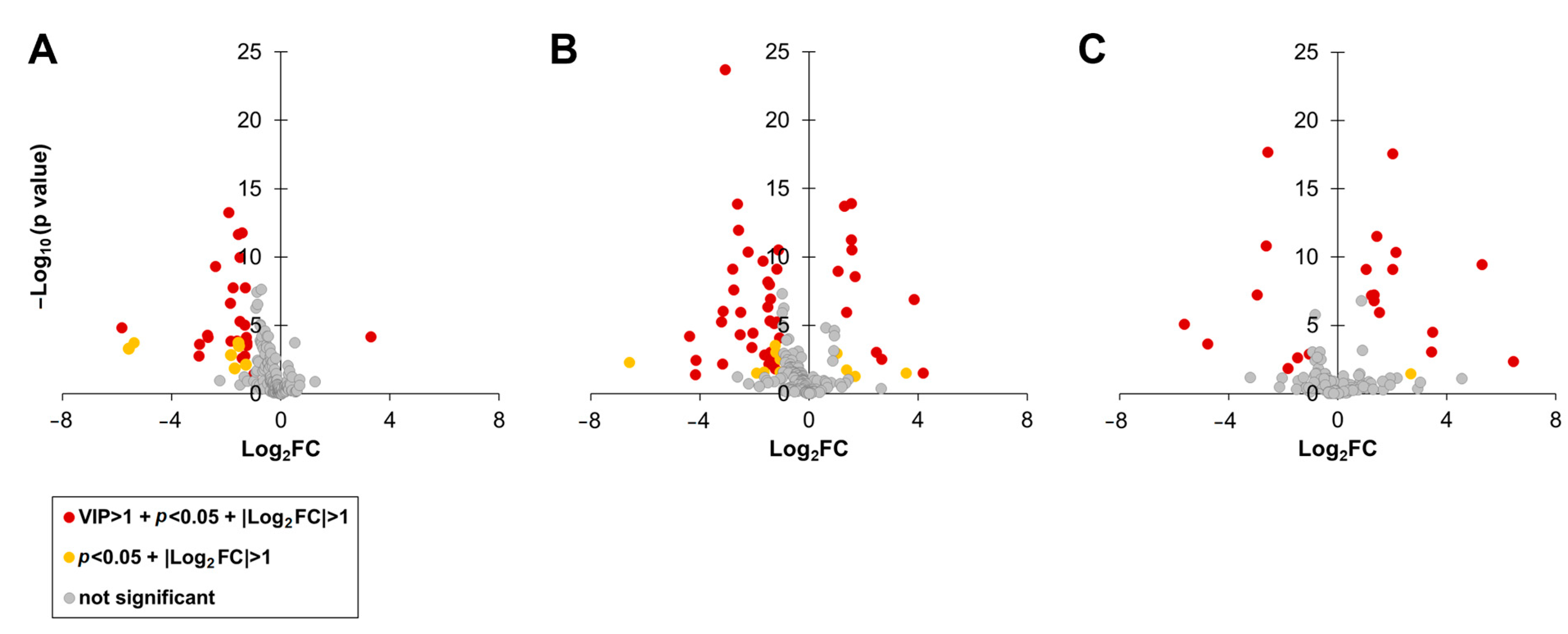
3.2. Metabolite Profiles of C. depressa Fruits
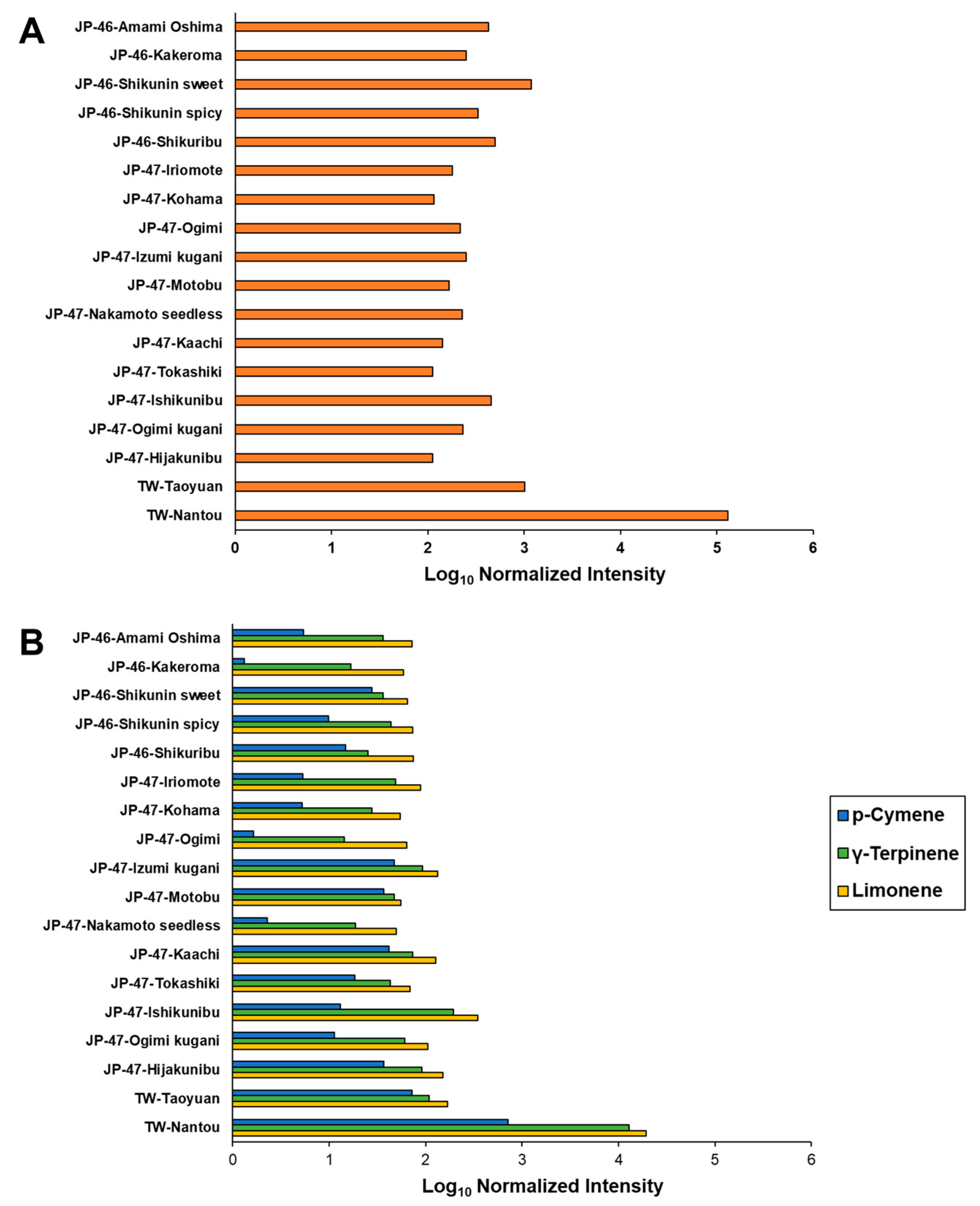
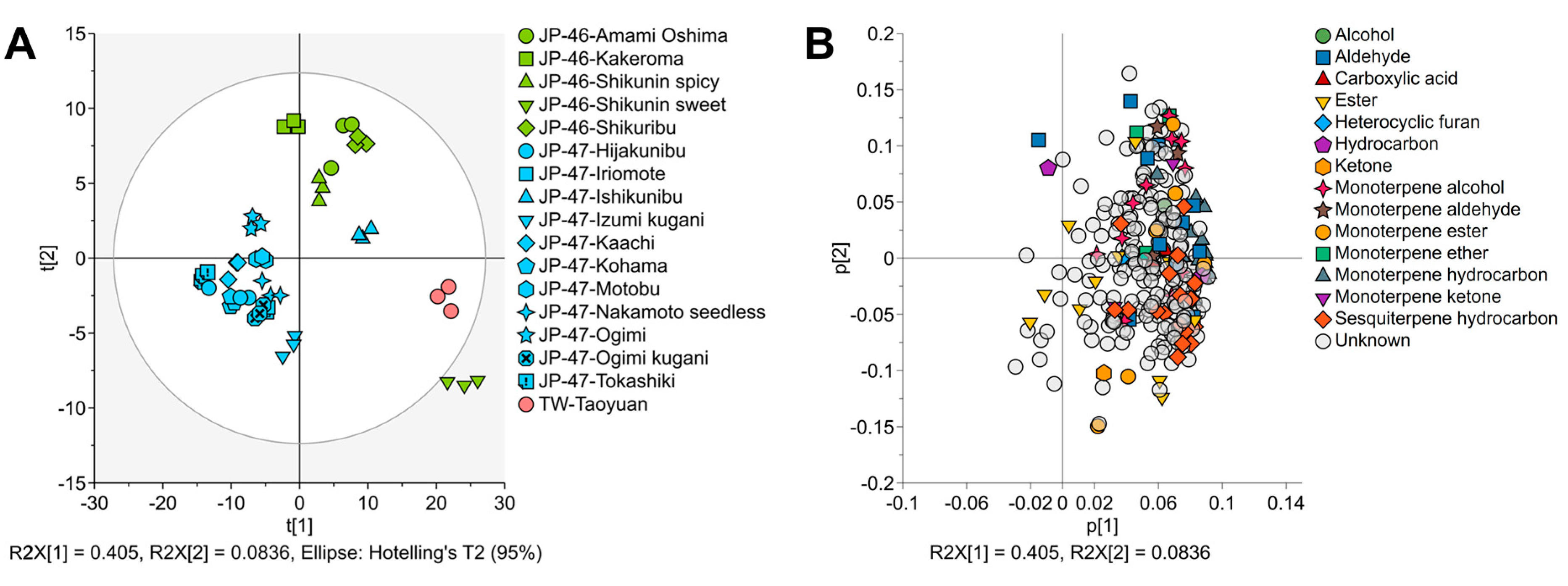
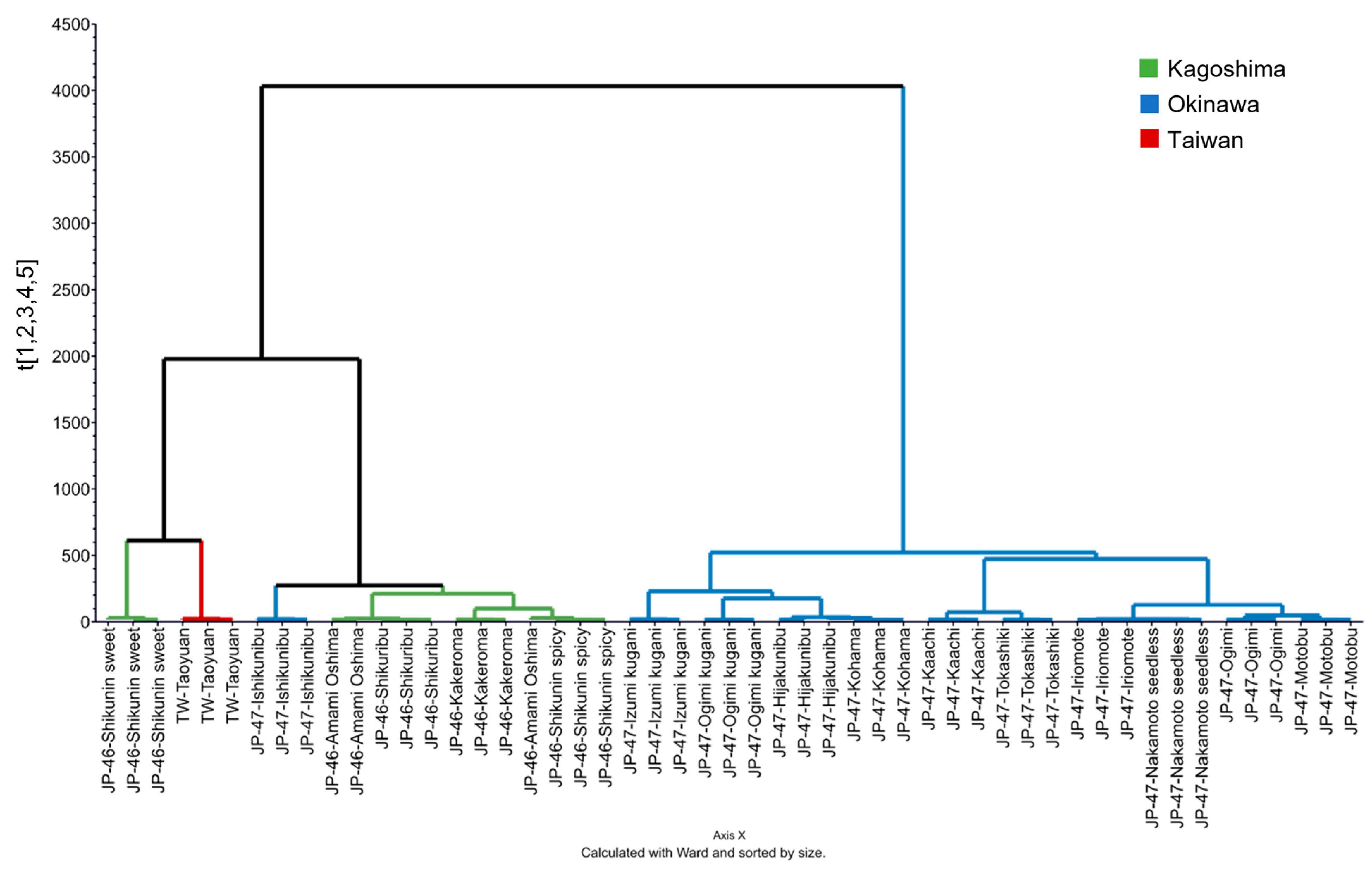
3.3. VOC Profiles of C. depressa Fruits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Asikin, Y.; Taira, I.; Inafuku-Teramoto, S.; Sumi, H.; Ohta, H.; Takara, K.; Wada, K. The composition of volatile aroma components, flavanones, and polymethoxylated flavones in Shiikuwasha (Citrus depressa Hayata) peels of different cultivation lines. J. Agric. Food Chem. 2012, 60, 7973–7980. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.Y.; Liao, Y.Y.; Chen, P.A. Leaf volatiles and relevant gene expression as the specific characteristics in Citrus depressa accession discrimination. Horticulturae 2022, 8, 773. [Google Scholar] [CrossRef]
- Asikin, Y.; Fukunaga, H.; Yamano, Y.; Hou, D.X.; Maeda, G.; Wada, K. Effect of cultivation line and peeling on food composition, taste characteristic, aroma profile, and antioxidant activity of Shiikuwasha (Citrus depressa Hayata) juice. J. Sci. Food Agric. 2014, 94, 2384–2392. [Google Scholar] [CrossRef] [PubMed]
- Asikin, Y.; Shimizu, K.; Iwasaki, H.; Oku, H.; Wada, K. Stress amelioration and anti-inflammatory potential of Shiikuwasha (Citrus depressa Hayata) essential oil, limonene, and γ-terpinene. J. Food Drug Anal. 2022, 30, 10. [Google Scholar] [CrossRef]
- Sadka, A.; Shlizerman, L.; Kamara, I.; Blumwald, E. Primary metabolism in citrus fruit as affected by its unique structure. Front. Plant Sci. 2019, 10, 1167. [Google Scholar] [CrossRef]
- Kim, D.S.; Lee, S.; Park, S.M.; Yun, S.H.; Gab, H.S.; Kim, S.S.; Kim, H.J. Comparative metabolomics analysis of citrus varieties. Foods 2021, 10, 2826. [Google Scholar] [CrossRef]
- Pan, T.; Kong, L.; Zhang, X.; Wang, Y.; Zhou, J.; Fu, Z.; Pan, H.; She, W.; Yu, Y. Fruit quality and volatile constituents of a new very early-ripening pummelo (Citrus maxima) cultivar ‘Liuyuezao’. Front. Plant Sci. 2023, 13, 1089009. [Google Scholar] [CrossRef]
- Sdiri, S.; Rambla, J.L.; Besada, C.; Granell, A.; Salvador, A. Changes in the volatile profile of citrus fruit submitted to postharvest degreening treatment. Postharvest. Biol. Technol. 2017, 133, 48–56. [Google Scholar] [CrossRef]
- Ren, J.N.; Tai, Y.N.; Dong, M.; Shao, J.H.; Yang, S.Z.; Pan, S.Y.; Fan, G. Characterisation of free and bound volatile compounds from six different varieties of citrus fruits. Food Chem. 2015, 185, 25–32. [Google Scholar] [CrossRef]
- Asikin, Y.; Kawahira, S.; Goki, M.; Hirose, N.; Kyoda, S.; Wada, K. Extended aroma extract dilution analysis profile of Shiikuwasha (Citrus depressa Hayata) pulp essential oil. J. Food Drug Anal. 2018, 26, 268–276. [Google Scholar] [CrossRef]
- Guarino, S.; Abbate, L.; Mercati, F.; Bosco, S.F.D.; Motisi, A.; Arif, M.A.; Cencetti, G.; Palagano, E.; Michelozzi, M. Citrus varieties with different tolerance grades to Tristeza Virus show dissimilar volatile terpene profiles. Agronomy 2021, 11, 1120. [Google Scholar] [CrossRef]
- Feng, S.; Niu, L.; Suh, J.H.; Hung, W.L.; Wang, Y. Comprehensive metabolomics analysis of Mandarins (Citrus reticulata) as a tool for variety, rootstock, and grove discrimination. J. Agric. Food Chem. 2018, 66, 10317–10326. [Google Scholar] [CrossRef]
- Centonze, V.; Lippolis, V.; Cervellieri, S.; Damascelli, A.; Casiello, G.; Pascale, M.; Logrieco, A.F.; Longobardi, F. Discrimination of geographical origin of oranges (Citrus sinensis L. Osbeck) by mass spectrometry-based electronic nose and characterization of volatile compounds. Food Chem. 2019, 277, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Tarancón, P.; Giménez-Sanchis, A.; Aleza, P.; Besada, C. Selection of new late-season Mandarin cultivars based on sensory changes and consumer acceptance after fruit cold storage. Agronomy 2021, 11, 116. [Google Scholar] [CrossRef]
- Tamura, Y.; Mori, T.; Nakabayashi, R.; Kobayashi, M.; Saito, K.; Okazaki, S.; Wang, N.; Kusano, M. Metabolomic evaluation of the quality of leaf lettuce grown in practical plant factory to capture metabolite signature. Front. Plant Sci. 2018, 9, 665. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Bai, J.; Chen, C.; Plotto, A.; Baldwin, E.A.; Gmitter, F.G. Comparative analysis of juice volatiles in selected mandarins, mandarin relatives and other citrus genotypes. J. Sci. Food Agric. 2018, 98, 1124–1131. [Google Scholar] [CrossRef]
- Kusano, M.; Kobayashi, M.; Iizuka, Y.; Fukushima, A.; Saito, K. Unbiased profiling of volatile organic compounds in the headspace of Allium plants using an in-tube extraction device. BMC Res. Notes 2016, 9, 133. [Google Scholar] [CrossRef] [Green Version]
- Tounsi, M.S.; Wannes, W.A.; Ouerghemmi, I.; Jegham, S.; Njima, Y.B.; Hamdaoui, G.; Zemni, H.; Marzouk, B. Juice components and antioxidant capacity of four Tunisian Citrus varieties. J. Sci. Food Agric. 2011, 91, 142–151. [Google Scholar] [CrossRef]
- Xu, G.; Liu, D.; Chen, J.; Ye, X.; Ma, Y.; Shi, J. Juice components and antioxidant capacity of citrus varieties cultivated in China. Food Chem. 2008, 106, 545–551. [Google Scholar] [CrossRef]
- Arbona, V.; Iglesias, D.J.; Gómez-Cadenas, A. Non-targeted metabolite profiling of citrus juices as a tool for variety discrimination and metabolite flow analysis. BMC Plant Biol. 2015, 15, 38. [Google Scholar] [CrossRef] [Green Version]
- Rokaya, P.R.; Baral, D.R.; Gautam, D.M.; Shrestha, A.K.; Paudyal, K.P. Effect of postharvest treatments on quality and shelf life of Mandarin (Citrus reticulata Blanco). Am. J. Plant Sci. 2016, 7, 1098–1105. [Google Scholar] [CrossRef] [Green Version]
- Montgomery, K.H.; Brar, G.; Krishnan, V.V. Metabolomics study at the postharvest conditions of cold storage and fungicide (Imazalil Sulfate) treatment in navel oranges and clementine mandarins. ACS Agric. Sci. Technol. 2022, 2, 79–89. [Google Scholar] [CrossRef]
- Lin, Q.; Wang, C.; Dong, W.; Jiang, Q.; Wang, D.; Li, S.; Chen, M.; Liu, C.; Sun, C.; Chen, K. Transcriptome and metabolome analyses of sugar and organic acid metabolism in Ponkan (Citrus reticulata) fruit during fruit maturation. Gene 2015, 554, 64–74. [Google Scholar] [CrossRef]
- Kim, S.S.; Kim, H.J.; Park, K.J.; Kang, S.B.; Park, Y.; Han, S.G.; Kim, M.; Song, Y.H.; Kim, D.S. Metabolomic profiling of Citrus unshiu during different stages of fruit development. Plants 2022, 11, 967. [Google Scholar] [CrossRef]
- Kimura, Y.; Naeshiro, M.; Tominaga, Y.; Anai, T.; Komai, F. Metabolite composition of grapefruit (Citrus paradisi) grown in Japan depends on the growing environment and harvest period. Hort. J. 2017, 86, 543–551. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Wang, J.; Tang, L.; Tang, J.; Liu, D.; Geng, F. Quantitative metabolomic analysis reveals the fractionation of active compounds during lemon fruit juicing. Food Res. Int. 2023, 169, 112829. [Google Scholar] [CrossRef]
- Liu, X.; Gmitter, F.G., Jr.; Grossera, J.W.; Wang, Y. Effects of rootstocks on the flavor quality of huanglongbing-affected sweet orange juices using targeted flavoromics strategy. RSC Adv. 2023, 13, 5590–5599. [Google Scholar] [CrossRef] [PubMed]
- Amit, S.K.; Uddin, M.M.; Rahman, R.; Islam, S.M.R.; Khan, M.S. A review on mechanisms and commercial aspects of food preservation and processing. Agric. Food Secur. 2017, 6, 51. [Google Scholar] [CrossRef]
- Drewnowski, A.; Gomez-Carneros, C. Bitter taste, phytonutrients, and the consumer: A review. Am. J. Clin. Nutr. 2000, 72, 1424–1435. [Google Scholar] [CrossRef] [Green Version]
- Asikin, Y.; Maeda, G.; Tamaki, H.; Mizu, M.; Oku, H.; Wada, K. Cultivation line and fruit ripening discriminations of Shiikuwasha (Citrus depressa Hayata) peel oils using aroma compositional, electronic nose, and antioxidant analyses. Food Res. Int. 2015, 67, 102–110. [Google Scholar] [CrossRef]
- Yamamoto, K.; Yahada, A.; Sasaki, K.; Funakoshi-Yoshida, A.; Ohta, C.; Koga, N.; Ohta, H. Detection of adulterated Shiikuwasha juice by sensory evaluation, colorimetric value and volatile components. Food Sci. Technol. Res. 2013, 19, 843–848. [Google Scholar] [CrossRef] [Green Version]
- Galvan-Lima, Â.; Cunha, S.C.; Martins, Z.E.; Soares, A.G.; Ferreira, I.M.P.L.V.O.; Farah, A. Headspace volatolome of peel flours from citrus fruits grown in Brazil. Food Res. Int. 2021, 150, 110801. [Google Scholar] [CrossRef] [PubMed]
- Kowalska, M.; Konopska, J.; Feszterová, M.; Zbikowska, A.; Kowalska, B. Quality assessment of natural juices and consumer preferences in the range of citrus fruit juices. Appl. Sci. 2023, 13, 765. [Google Scholar] [CrossRef]
- Lubinska-Szczygeł, M.; Polkowska, Z.; Dymerski, T.; Gorinstein, S. Comparison of the physical and sensory properties of hybrid citrus fruit Jaffa® sweetie in relation to the parent fruits. Molecules 2020, 25, 2748. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Asikin, Y.; Tamura, Y.; Aono, Y.; Kusano, M.; Shiba, H.; Yamamoto, M.; Mitsube, F.; Lin, S.-Y.; Takara, K.; Wada, K. Multivariate Profiling of Metabolites and Volatile Organic Compounds in Citrus depressa Hayata Fruits from Kagoshima, Okinawa, and Taiwan. Foods 2023, 12, 2951. https://doi.org/10.3390/foods12152951
Asikin Y, Tamura Y, Aono Y, Kusano M, Shiba H, Yamamoto M, Mitsube F, Lin S-Y, Takara K, Wada K. Multivariate Profiling of Metabolites and Volatile Organic Compounds in Citrus depressa Hayata Fruits from Kagoshima, Okinawa, and Taiwan. Foods. 2023; 12(15):2951. https://doi.org/10.3390/foods12152951
Chicago/Turabian StyleAsikin, Yonathan, Yoshio Tamura, Yusuke Aono, Miyako Kusano, Hiroshi Shiba, Masashi Yamamoto, Fumimasa Mitsube, Shu-Yen Lin, Kensaku Takara, and Koji Wada. 2023. "Multivariate Profiling of Metabolites and Volatile Organic Compounds in Citrus depressa Hayata Fruits from Kagoshima, Okinawa, and Taiwan" Foods 12, no. 15: 2951. https://doi.org/10.3390/foods12152951
APA StyleAsikin, Y., Tamura, Y., Aono, Y., Kusano, M., Shiba, H., Yamamoto, M., Mitsube, F., Lin, S. -Y., Takara, K., & Wada, K. (2023). Multivariate Profiling of Metabolites and Volatile Organic Compounds in Citrus depressa Hayata Fruits from Kagoshima, Okinawa, and Taiwan. Foods, 12(15), 2951. https://doi.org/10.3390/foods12152951