


Internalization of Salmonella in Leafy Vegetables during Postharvest Conditions

Abstract
:1. Introduction
2. Materials and Methods
2.1. Fresh Produce Samples and Pathogenic Bacteria: Salmonella
2.2. Salmonella Inoculation
2.3. Removing Bacteria from the Surface of Leaves and Quantification of Internalized Bacteria
2.4. Imaging Internalized GFP-Labeled Salmonella
2.5. Quantification of Internalized GFP-Labeled Salmonella from the Digital Images
2.6. Statistical Analysis
3. Results
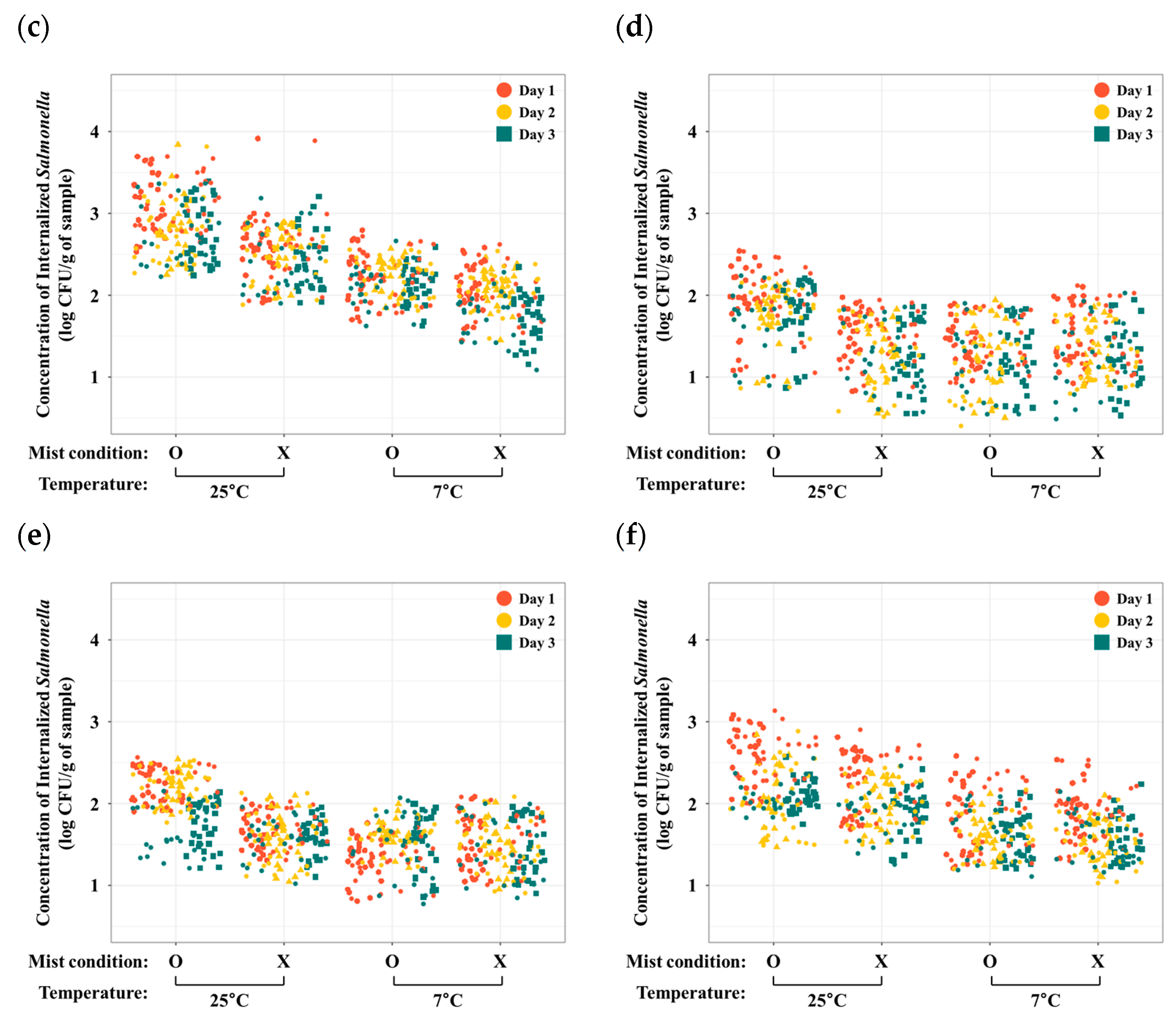
3.1. Effect of Storage Conditions on Salmonella Internalization
3.1.1. Iceberg Lettuce
3.1.2. Romaine Lettuce
3.1.3. Red Lettuce
3.1.4. Green Onion
3.1.5. Kale
3.1.6. Spinach
3.2. Effects of Temporal Variation on Salmonella Internalization
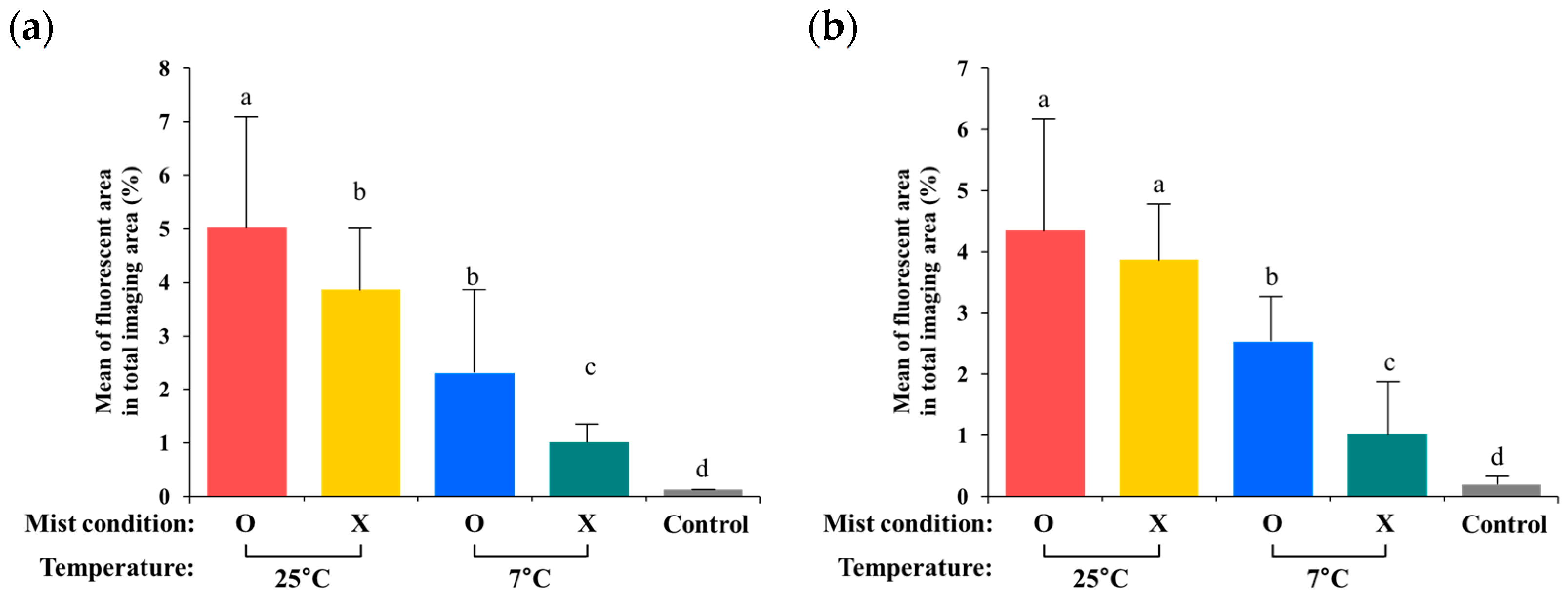
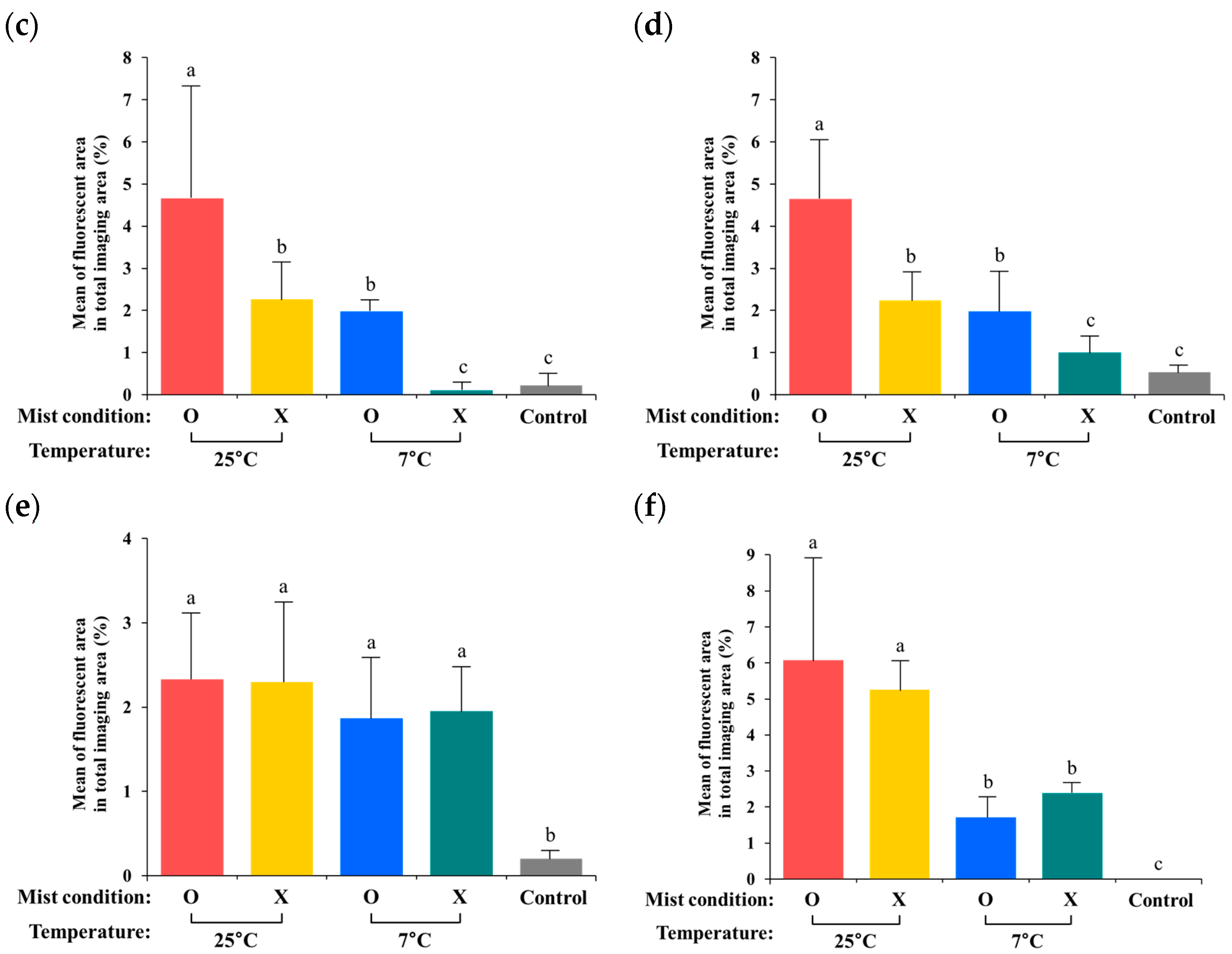
3.3. Quantification of Internalized GFP-Labeled Salmonella from the Digital Images
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Erickson, M.C.; Liao, J.Y.; Payton, A.S.; Cook, P.W.; Ortega, Y.R. Survival and internalization of Salmonella and Escherichia coli O157: H7 sprayed onto different cabbage cultivars during cultivation in growth chambers. J. Sci. Food Agric. 2019, 99, 3530–3537. [Google Scholar] [CrossRef]
- World Health Organization. Regional Office for the Eastern Mediterranean. In Healthy Diet; World Health Organization, Regional Office for the Eastern Mediterranean: Cairo, Egypt, 2019. [Google Scholar]
- Isabel, R.N.M.; Cristina, U.; Ruth, H.O.; Carmen, G.P.M. Reported Foodborne Outbreaks Due to Fresh Produce in the United States and European Union: Trends and Causes. Foodborne Pathog. Dis. 2015, 12, 32–38. [Google Scholar]
- Grivokostopoulos, N.C.; Makariti, I.P.; Tsadaris, S.; Skandamis, P.N. Impact of population density and stress adaptation on the internalization of Salmonella in leafy greens. Food Microbiol. 2022, 106, 104053. [Google Scholar] [CrossRef] [PubMed]
- Golberg, D.; Kroupitski, Y.; Belausov, E.; Pinto, R.; Sela, S. Salmonella Typhimurium internalization is variable in leafy vegetables and fresh herbs. Int. J. Food Microbiol. 2011, 145, 250–257. [Google Scholar] [CrossRef]
- Ge, C.; Lee, C.; Lee, J. The impact of extreme weather events on Salmonella internalization in lettuce and green onion. Food Res. Int. 2012, 45, 1118–1122. [Google Scholar] [CrossRef]
- Kroupitski, Y.; Gollop, R.; Belausov, E.; Pinto, R.; Sela, S. Salmonella enterica growth conditions influence lettuce leaf internalization. Front. Microbiol. 2019, 10, 639. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Chambers, H.; DiCaprio, E.; Gao, G.; Li, J. Internalization and dissemination of human norovirus and Tulane virus in fresh produce is plant dependent. Food Microbiol. 2018, 69, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Mok, C.; Lee, J. Photocatalytically Enhanced Inactivation of Internalized Pathogenic Bacteria in Fresh Produce Using UV Irradiation with Nano-Titanium Dioxide. J. Food Prot. 2021, 84, 820–826. [Google Scholar] [CrossRef]
- Eissenberger, K.; Drissner, D.; Walsh, F.; Weiss, A.; Schmidt, H. Plant variety and soil type influence Escherichia coli O104: H4 strain C227/11ϕcu adherence to and internalization into the roots of lettuce plants. Food Microbiol. 2020, 86, 103316. [Google Scholar] [CrossRef]
- Moriarty, M.J.; Semmens, K.; Bissonnette, G.K.; Jaczynski, J. Inactivation with UV-radiation and internalization assessment of coliforms and Escherichia coli in aquaponically grown lettuce. LWT 2018, 89, 624–630. [Google Scholar] [CrossRef]
- Ssemanda, J.N.; Reij, M.W.; van Middendorp, G.; Bouw, E.; van der Plaats, R.; Franz, E.; Muvunyi, C.M.; Bagabe, M.C.; Zwietering, M.H.; Joosten, H. Foodborne pathogens and their risk exposure factors associated with farm vegetables in Rwanda. Food Control 2018, 89, 86–96. [Google Scholar] [CrossRef]
- Erickson, M.C. Internalization of fresh produce by foodborne pathogens. Annu. Rev. Food Sci. Technol. 2012, 3, 283–310. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Luo, Y.; Nou, X.; Yang, Y.; Wu, Y.; Wang, Q. Effects of postharvest handling conditions on internalization and growth of Salmonella enterica in tomatoes. J. Food Prot. 2014, 77, 365–370. [Google Scholar] [CrossRef]
- Frankel, G.; Shaw, R.K.; Pink, D.; Berger, C.N.; Hand, P. Fresh produce as a potential vector for bacterial human pathogens. Microb. Biotechnol. 2009, 2, 595. [Google Scholar] [CrossRef]
- Ibarra-Sanchez, L.S.; Alvarado-Casillas, S.; Rodriguez-Garcia, M.O.; Martinez-Gonzales, N.E.; Castillo, A. Internalization of bacterial pathogens in tomatoes and their control by selected chemicals. J. Food Prot. 2004, 67, 1353–1358. [Google Scholar] [CrossRef] [PubMed]
- Eblen, B.S.; Walderhaug, M.O.; Edelson-Mammel, S.; Chirtel, S.J.; De Jesus, A.; Merker, R.I.; Buchanan, R.L.; Miller, A.J. Potential for internalization, growth, and survival of Salmonella and Escherichia coli O157: H7 in oranges. J. Food Prot. 2004, 67, 1578–1584. [Google Scholar] [CrossRef] [PubMed]
- Warriner, K.; Ibrahim, F.; Dickinson, M.; Wright, C.; Waites, W.M. Internalization of human pathogens within growing salad vegetables. Biotechnol. Genet. Eng. Rev. 2003, 20, 117–136. [Google Scholar] [CrossRef]
- Riggio, G.M.; Jones, S.L.; Gibson, K.E. Risk of human pathogen internalization in leafy vegetables during lab-scale hydroponic cultivation. Horticulturae. 2019, 5, 25. [Google Scholar] [CrossRef]
- Yoon, J.H.; Lee, S.Y. Comparison of the effectiveness of decontaminating strategies for fresh fruits and vegetables and related limitations. Crit. Rev. Food Sci. Nutr. 2018, 58, 3189–3208. [Google Scholar] [CrossRef]
- Park, S.; Navratil, S.; Gregory, A.; Bauer, A.; Srinath, I.; Jun, M.; Szonyi, B.; Nightingale, K.; Anciso, J.; Ivanek, R. Generic Escherichia coli contamination of spinach at the preharvest stage: Effects of farm management and environmental factors. Appl. Environ. Microbiol. 2013, 79, 4347–4358. [Google Scholar] [CrossRef]
- Gu, G.; Cevallos-Cevallos, J.M.; van Bruggen, A.H.C. Ingress of Salmonella enterica Typhimurium into tomato leaves through hydathodes. PLoS ONE 2013, 8, e53470. [Google Scholar] [CrossRef]
- Xia, X.; Luo, Y.; Yang, Y.; Vinyard, B.; Schneider, K.; Meng, J. Effects of tomato variety, temperature differential, and post–stem removal time on internalization of Salmonella enterica serovar Thompson in tomatoes. J. Food Prot. 2012, 75, 297–303. [Google Scholar] [CrossRef]
- Yaron, S.; Römling, U. Biofilm formation by enteric pathogens and its role in plant colonization and persistence. Microb. Biotechnol. 2014, 7, 496–516. [Google Scholar] [CrossRef]
- Iturriaga, M.H.; Tamplin, M.L.; Escartin, E.F. Colonization of tomatoes by Salmonella Montevideo is affected by relative humidity and storage temperature. J. Food Prot. 2007, 70, 30–34. [Google Scholar] [CrossRef]
- Merget, B.; Forbes, K.J.; Brennan, F.; McAteer, S.; Shepherd, T.; Strachan, N.J.; Holden, N.J. Influence of plant species, tissue type, and temperature on the capacity of Shiga-Toxigenic Escherichia coli to colonize, grow, and be internalized by plants. Appl. Environ. Microbiol. 2019, 85, e00123-19. [Google Scholar] [CrossRef]
- Warriner, K.; Huber, A.; Namvar, A.; Fan, W.; Dunfield, K. Recent advances in the microbial safety of fresh fruits and vegetables. Adv. Food Nutr. Res. 2009, 57, 155–208. [Google Scholar] [PubMed]
- Hirneisen, K.A.; Sharma, M.; Kniel, K.E. Human enteric pathogen internalization by root uptake into food crops. Foodborne Pathog. Dis. 2012, 9, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Deering, A.J.; Mauer, L.J.; Pruitt, R.E. Internalization of E. coli O157: H7 and Salmonella spp. in plants: A review. Food Res. Int. 2012, 45, 567–575. [Google Scholar] [CrossRef]
- Solomon, E.B.; Yaron, S.; Matthews, K.R. Transmission of Escherichia coli O157: H7 from contaminated manure and irrigation water to lettuce plant tissue and its subsequent internalization. Appl. Environ. Microbiol. 2002, 68, 397–400. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group 1 25 °C with Mist | Group 2 25 °C without Mist | Group 3 7 °C with Mist | Group 4 7 °C without Mist | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Day1 | Day2 | Day3 | Day1 | Day2 | Day3 | Day1 | Day2 | Day3 | Day1 | Day2 | Day3 | ||
| Iceberg lettuce | Mean | 3.2 | 2.9 | 2.9 | 2.8 | 2.8 | 2.6 | 2.5 | 2.1 | 2.0 | 1.7 | 1.7 | 1.7 |
| Max | 4.2 | 3.5 | 3.5 | 3.3 | 2.8 | 3.3 | 3.2 | 2.7 | 2.4 | 2.1 | 2.3 | 2.2 | |
| Min | 2.3 | 2.4 | 2.5 | 2.3 | 2.1 | 2.2 | 2.1 | 1.6 | 1.3 | 1.0 | 1.3 | 1.1 | |
| SE * | 0.5 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.2 | 0.3 | 0.3 | 0.3 | |
| Romaine lettuce | Mean | 3.0 | 2.7 | 2.7 | 2.6 | 2.5 | 2.4 | 2.2 | 2.0 | 1.9 | 1.9 | 1.9 | 1.8 |
| Max | 3.3 | 3.1 | 3.2 | 3.1 | 3.0 | 3.0 | 2.6 | 2.5 | 2.1 | 2.4 | 2.8 | 2.2 | |
| Min | 2.5 | 2.1 | 2.2 | 1.9 | 2.0 | 1.8 | 1.7 | 1.4 | 1.6 | 1.3 | 1.3 | 0.9 | |
| SE | 0.2 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.2 | 0.3 | 0.1 | 0.3 | 0.4 | 0.3 | |
| Red lettuce | Mean | 3.1 | 2.8 | 2.8 | 2.6 | 2.5 | 2.4 | 2.3 | 2.2 | 2.1 | 2.1 | 2.1 | 1.7 |
| Max | 3.7 | 3.8 | 3.4 | 3.9 | 2.9 | 3.2 | 2.8 | 2.6 | 2.6 | 2.6 | 2.5 | 2.3 | |
| Min | 2.5 | 2.3 | 2.2 | 1.9 | 2.0 | 1.9 | 1.7 | 1.9 | 1.4 | 1.6 | 1.5 | 1.2 | |
| SE | 0.3 | 0.4 | 0.4 | 0.4 | 0.3 | 0.3 | 0.3 | 0.2 | 0.3 | 0.2 | 0.2 | 0.3 | |
| Green onion | Mean | 1.9 | 1.8 | 1.8 | 1.5 | 1.2 | 1.2 | 1.4 | 1.2 | 1.2 | 1.5 | 1.3 | 1.2 |
| Max | 2.5 | 2.2 | 2.2 | 2.0 | 1.8 | 1.9 | 1.9 | 1.9 | 2.1 | 1.8 | 1.9 | 1.9 | |
| Min | 1.1 | 0.9 | 0.9 | 0.8 | 0.6 | 0.6 | 1.0 | 0.5 | 0.9 | 0.6 | 0.9 | 0.5 | |
| SE | 0.4 | 0.3 | 0.4 | 0.3 | 0.4 | 0.4 | 0.3 | 0.4 | 0.4 | 0.4 | 0.3 | 0.4 | |
| Kale | Mean | 2.2 | 2.2 | 1.7 | 1.7 | 1.6 | 1.6 | 1.3 | 1.6 | 1.5 | 1.5 | 1.5 | 1.5 |
| Max | 2.5 | 2.5 | 2.1 | 2.0 | 2.1 | 2.0 | 1.8 | 2.0 | 2.1 | 2.0 | 2.0 | 2.0 | |
| Min | 1.9 | 1.8 | 1.2 | 1.3 | 1.0 | 1.1 | 0.8 | 1.2 | 1.0 | 0.9 | 1.0 | 0.9 | |
| SE | 0.2 | 0.2 | 0.3 | 0.2 | 0.3 | 0.2 | 0.3 | 0.2 | 0.3 | 0.3 | 0.3 | 0.3 | |
| Spinach | Mean | 2.6 | 2.1 | 2.1 | 2.3 | 2.0 | 1.9 | 1.8 | 1.6 | 1.6 | 1.9 | 1.5 | 1.6 |
| Max | 3.1 | 2.8 | 2.6 | 2.8 | 2.4 | 2.4 | 2.6 | 2.1 | 2.5 | 2.1 | 2.1 | 2.2 | |
| Min | 2.0 | 1.5 | 1.7 | 1.7 | 1.5 | 1.3 | 1.2 | 1.2 | 1.3 | 1.2 | 1.1 | 1.2 | |
| SE | 0.3 | 0.4 | 0.2 | 0.4 | 0.3 | 0.3 | 0.4 | 0.2 | 0.3 | 0.3 | 0.2 | 0.3 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.; Park, S.; Lee, J.; Lee, S. Internalization of Salmonella in Leafy Vegetables during Postharvest Conditions. Foods 2023, 12, 3106. https://doi.org/10.3390/foods12163106
Kim J, Park S, Lee J, Lee S. Internalization of Salmonella in Leafy Vegetables during Postharvest Conditions. Foods. 2023; 12(16):3106. https://doi.org/10.3390/foods12163106
Chicago/Turabian StyleKim, Jinnam, Soeun Park, Jiyoung Lee, and Seungjun Lee. 2023. "Internalization of Salmonella in Leafy Vegetables during Postharvest Conditions" Foods 12, no. 16: 3106. https://doi.org/10.3390/foods12163106
APA StyleKim, J., Park, S., Lee, J., & Lee, S. (2023). Internalization of Salmonella in Leafy Vegetables during Postharvest Conditions. Foods, 12(16), 3106. https://doi.org/10.3390/foods12163106