
Physicochemical Quality, Polyphenol Profiles, and Postharvest Performance of Florida Pearl® ‘FL 16.78-109’ White Strawberries Compared to the Red Cultivar ‘Florida Brilliance’ †


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Weather Conditions
2.3. Storage Conditions
2.4. Instrumental Color and Texture Analysis
2.5. Weight Loss and Dry Weight
2.6. Acidity and Soluble Solids Content
2.7. Sugar Analysis
2.8. Ascorbic Acid Analysis
2.9. Total Phenolics and Anthocyanin Analysis
2.10. Polyphenol Extraction
2.11. Individual Polyphenol Analysis
2.12. Statistical Analysis
3. Results and Discussion
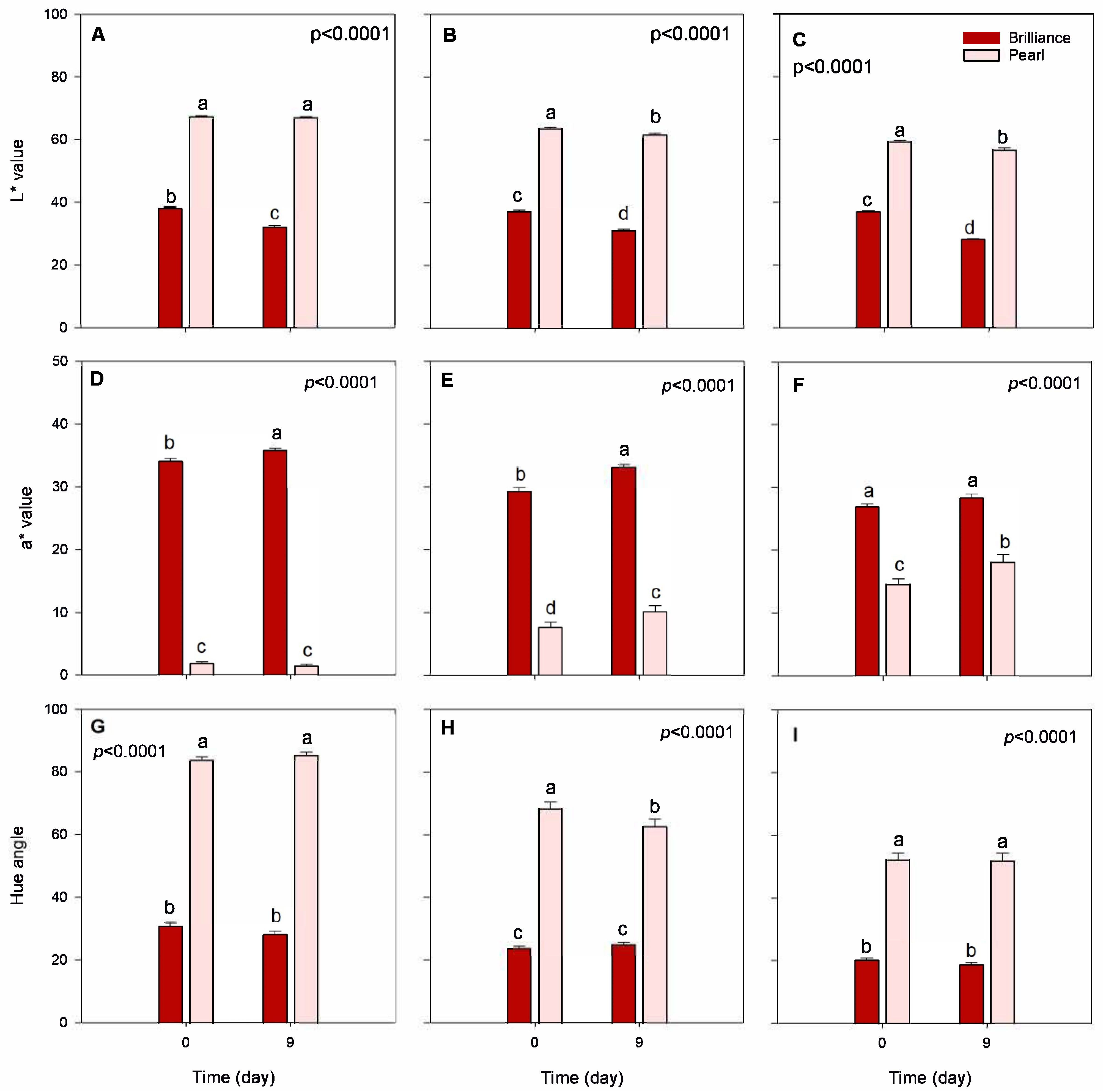
3.1. Color: Differences between Pearl and Brilliance, Seasonal Variability, and Impact of Storage
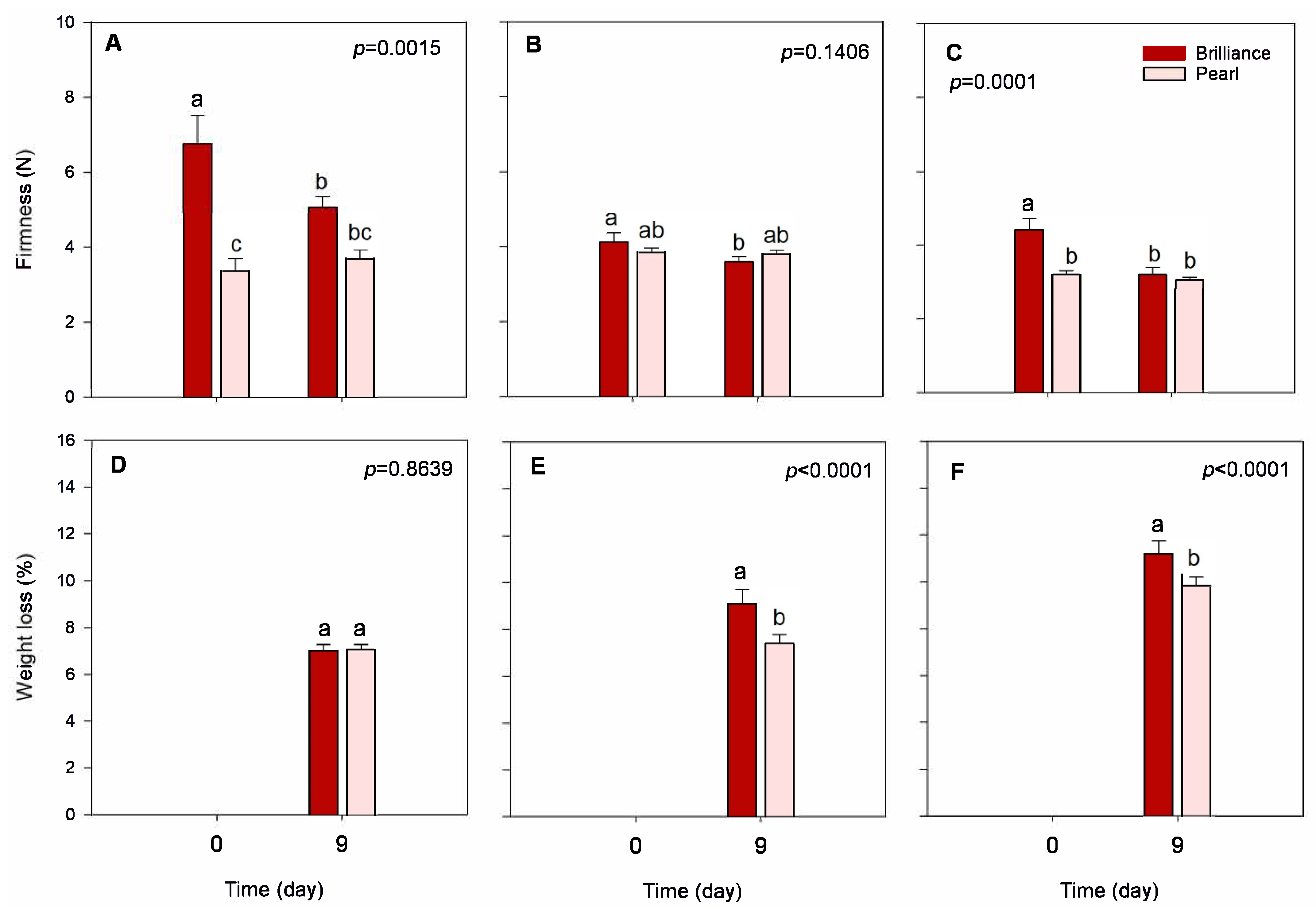
3.2. Firmness: Differences between Pearl and Brilliance, Seasonal Variability, and Impact of Storage
3.3. Chemical Composition: Differences between Pearl and Brilliance, Seasonal Variability, and Impact of Storage
3.3.1. Weight Loss
3.3.2. Acidity and Soluble Solids Content
3.3.3. Sugar Profiles
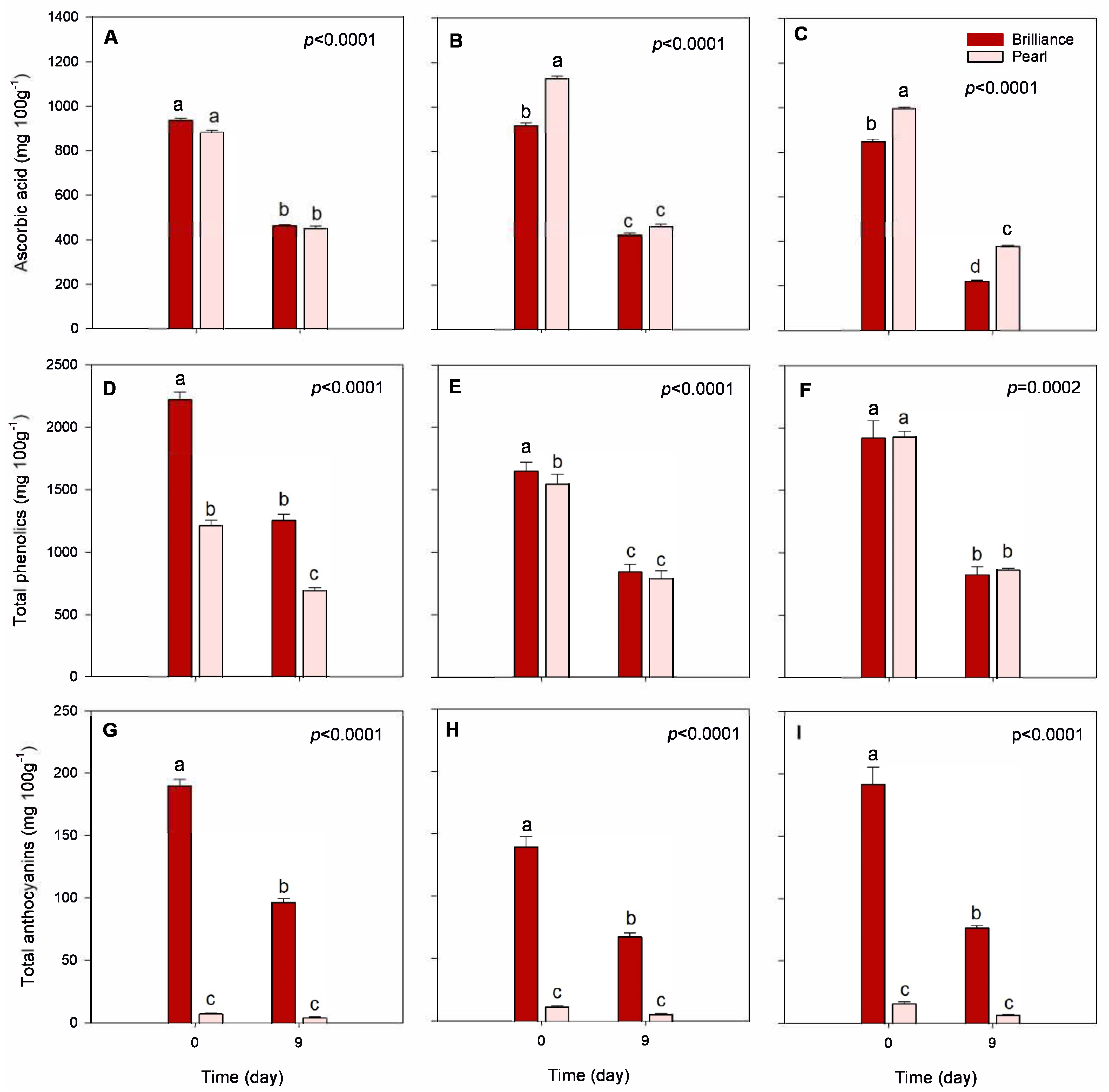
3.3.4. Ascorbic Acid Content
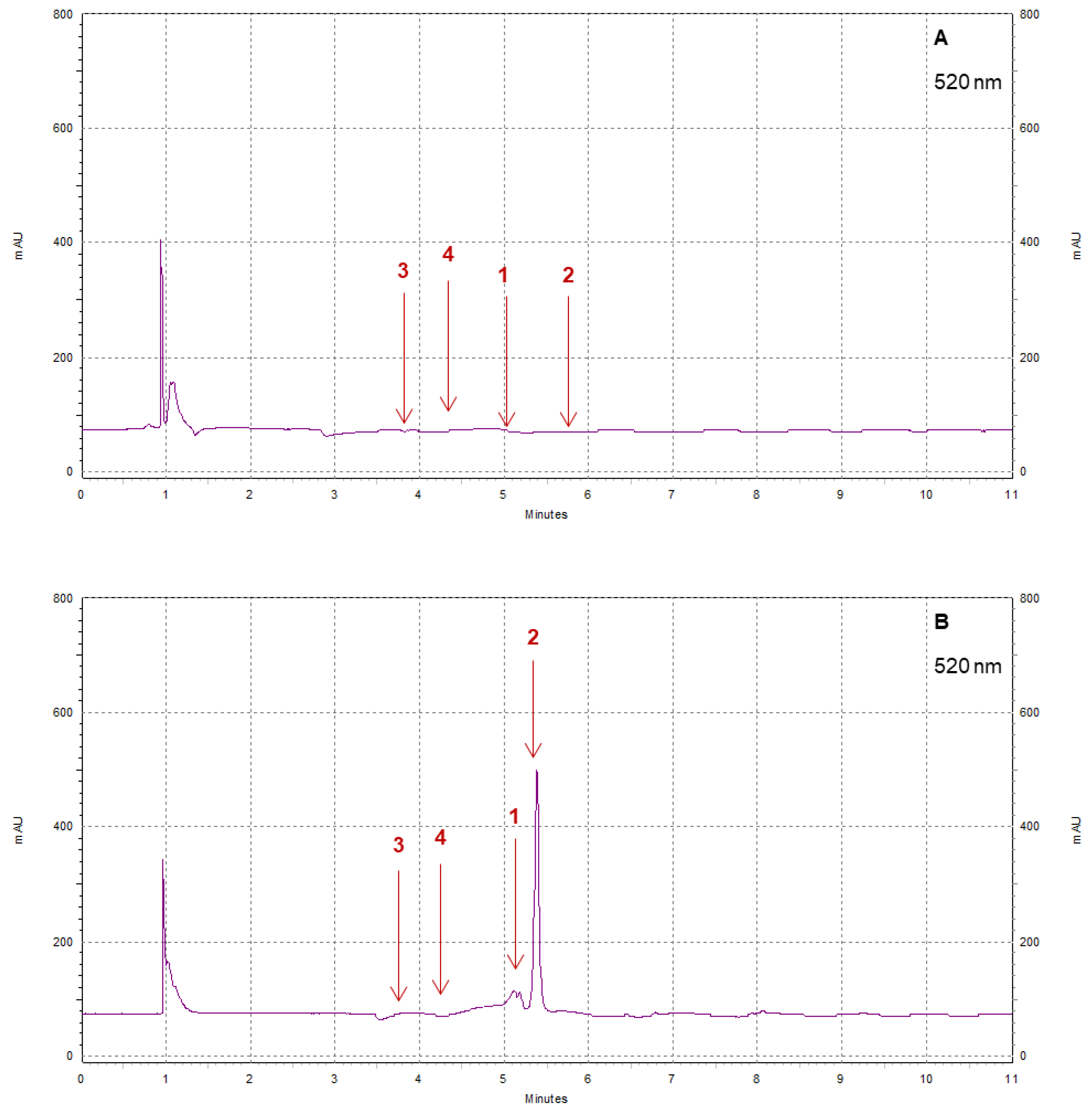
3.3.5. Total Phenolic and Anthocyanin Contents
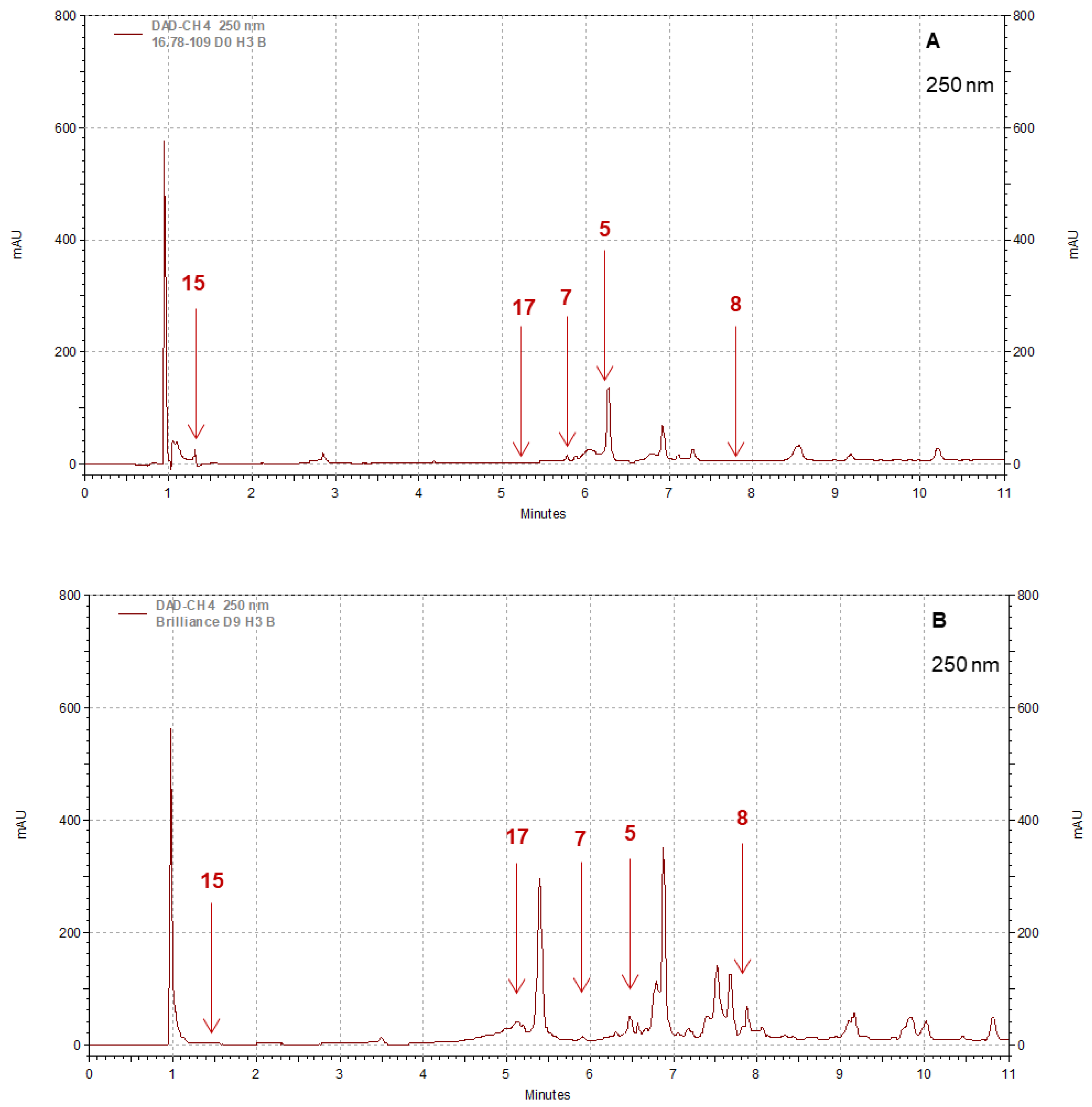
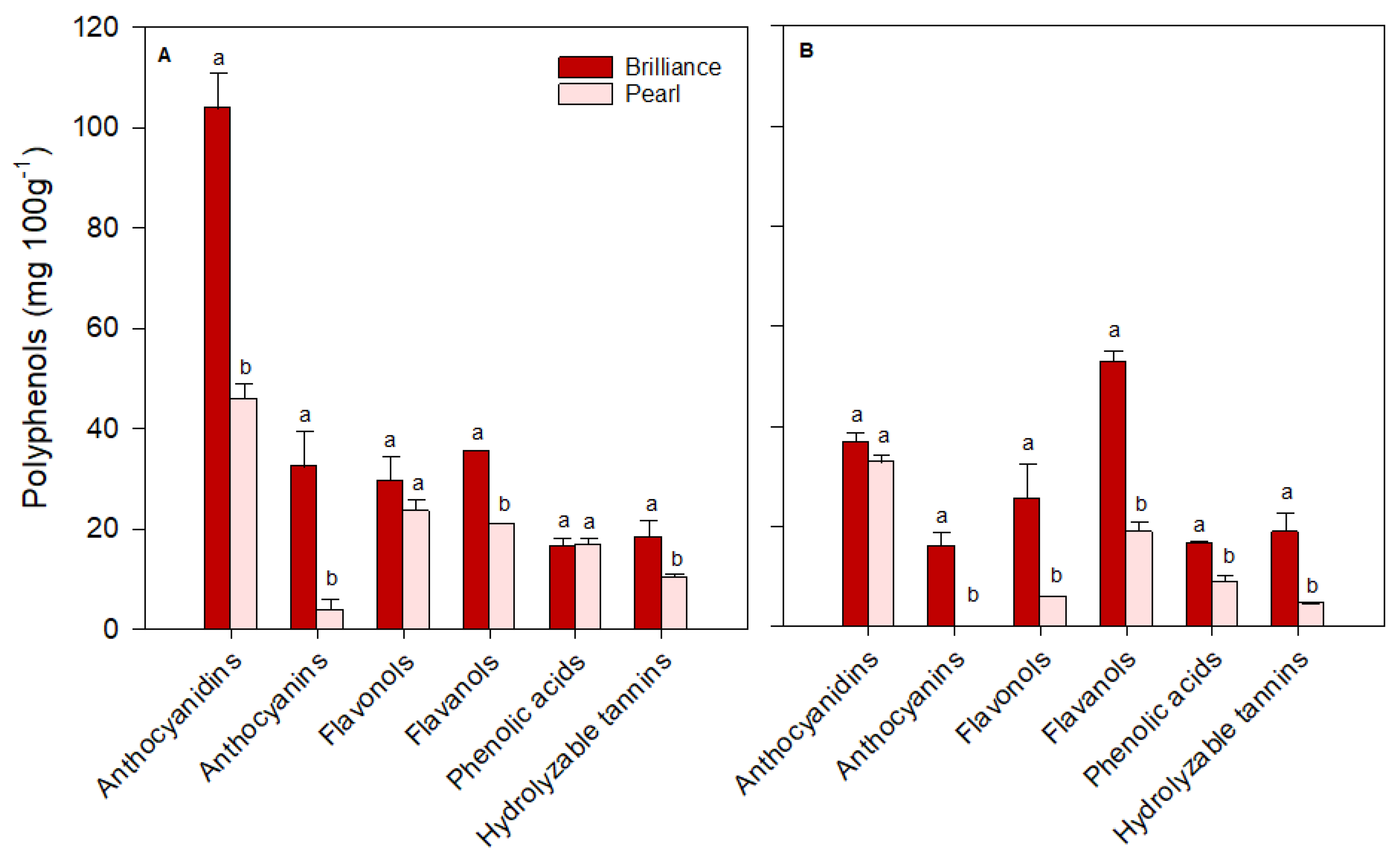
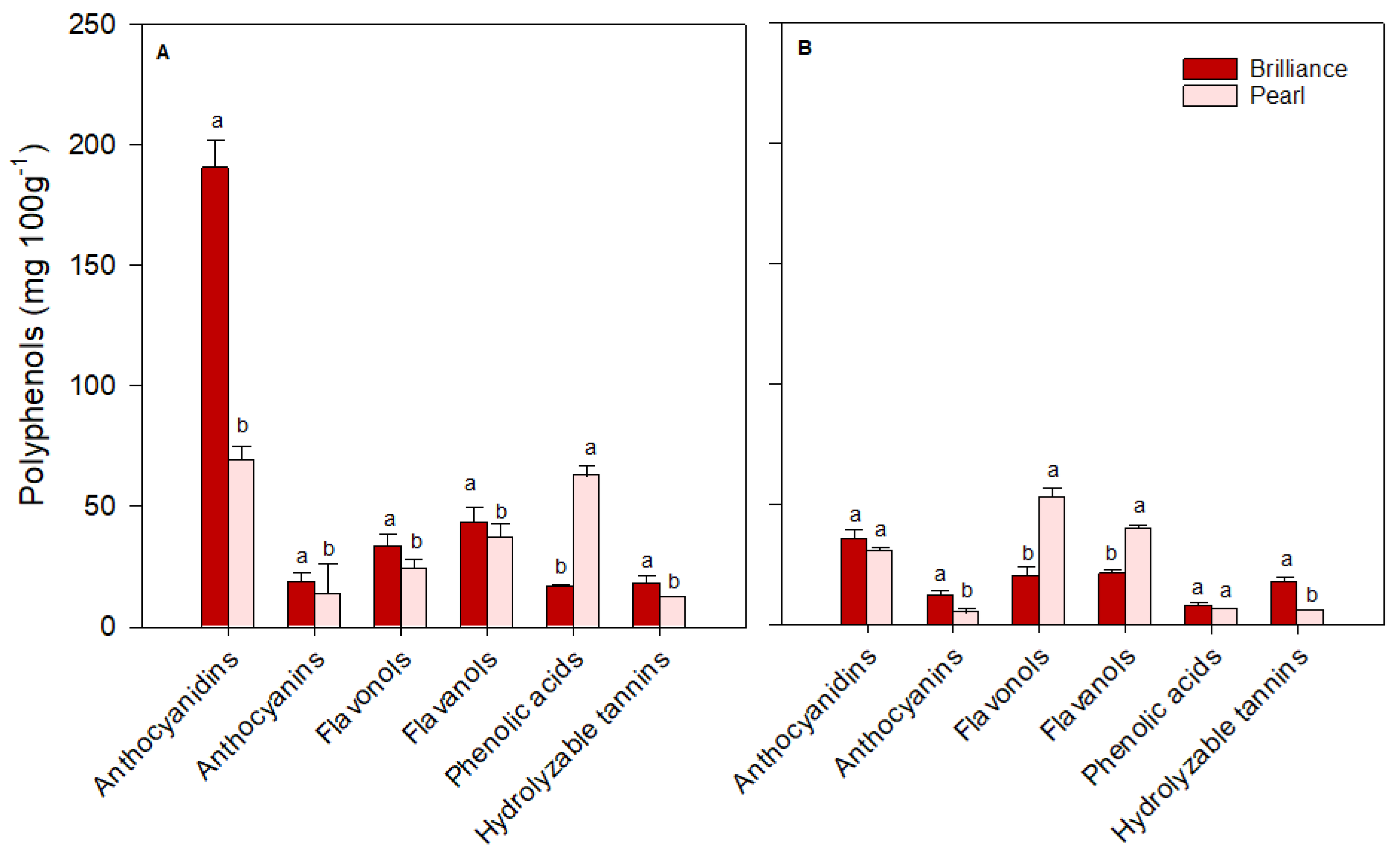
3.4. Polyphenol Profiles: Differences between Pearl and Brilliance, Seasonal Variability, and Impact of Storage
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Finn, C.E.; Retamales, J.B.; Lobos, G.A.; Hancock, J.F. The Chilean strawberry (Fragaria chiloensis): Over 1000 years of domestication. HortScience 2013, 48, 418–421. [Google Scholar] [CrossRef]
- Carrasco, B.; Garcés, M.; Rojas, P.; Saud, G.; Herrera, R.; Retamales, J.B.; Caligari, P.D. The Chilean strawberry [Fragaria chiloensis (L.) Duch.]: Genetic diversity and structure. J. Am. Soc. Hortic. Sci. 2007, 132, 501–506. [Google Scholar] [CrossRef]
- Morales-Quintana, L.; Ramos, P. Chilean strawberry (Fragaria chiloensis): An integrative and comprehensive review. Food Res. Int. 2019, 119, 769–776. [Google Scholar] [CrossRef]
- Prat, L.; Espinoza, M.I.; Agosin, E.; Silva, H. Identification of volatile compounds associated with the aroma of white strawberries (Fragaria chiloensis). J. Sci. Food Agric. 2014, 94, 752–759. [Google Scholar] [CrossRef] [PubMed]
- Whitaker, V.M.; Dalid, C.; Osorio, L.F.; Peres, N.A.; Verma, S.; Lee, S.; Plotto, A. Florida Pearl®‘FL 16.78-109’Pineberry. HortScience 2023, 58, 143–146. [Google Scholar] [CrossRef]
- Aaby, K.; Mazur, S.; Nes, A.; Skrede, G. Phenolic compounds in strawberry (Fragaria × ananassa Duch.) fruits: Composition in 27 cultivars and changes during ripening. Food Chem. 2012, 132, 86–97. [Google Scholar] [CrossRef]
- Steyn, W.; Wand, S.J.E.; Holcraft, D.M.; Jacobs, G. Anthocyanins in vegetative tissues:A proposed unified function in photoprotection. New Phytol. 2002, 155, 349–361. [Google Scholar] [CrossRef]
- Smillie, R.M.; Hetherington, S.E. Photoabatement by anthocyanin shields photosynthetic systems from light stress. Photosynthetica 1999, 36, 451–463. [Google Scholar] [CrossRef]
- Cominelli, E.; Gusmaroli, G.; Allegra, D.; Galbiati, M.; Wade, H.K.; Jenkins, G.I.; Tonelli, C. Expression analysis of anthocyanin regulatory genes in response to different light qualities in Arabidopsis thaliana. J. Plant Physiol. 2008, 165, 886–894. [Google Scholar] [CrossRef]
- Palmieri, L.; Masuero, D.; Martinatti, P.; Baratto, G.; Martens, S.; Vrhovsek, U. Genotype-by-environment effect on bioactive compounds in strawberry (Fragaria × ananassa Duch.). J. Sci. Food Agric. 2017, 97, 4180–4189. [Google Scholar] [CrossRef]
- Anttonen, M.J.; Hoppula, K.I.; Netstby, R.; Verheul, M.J.; Karjalainen, R.O. Influence of fertilization, mulch color, early forcing, fruit order, planting date, shading, growing environment, and genotype on the contents of selected phenolics in strawberry (Fragaria × ananassa Duch.) fruits. J. Agric. Food Chem. 2006, 54, 2614–2620. [Google Scholar] [CrossRef] [PubMed]
- Hjernø, K.; Alm, R.; Canbäck, B.; Matthiesen, R.; Trajkovski, K.; Björk, L.; Roepstorff, P.; Emanuelsson, C. Down-regulation of the strawberry Bet v 1-homologous allergen in concert with the flavonoid biosynthesis pathway in colorless strawberry mutant. Proteomics 2006, 6, 1574–1587. [Google Scholar] [CrossRef] [PubMed]
- Salvatierra, A.; Pimentel, P.; Moya-León, M.A.; Herrera, R. Increased accumulation of anthocyanins in Fragaria chiloensis fruits by transient suppression of FcMYB1 gene. Phytochemistry 2013, 90, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Saud, G.; Carbone, F.; Perrotta, G.; Figueroa, C.R.; Moya, M.; Herrera, R.; Retamales, J.B.; Carrasco, B.; Cheel, J.; Schmeda-Hirschmann, G.; et al. Transcript profiling suggests transcriptional repression of the flavonoid pathway in the white-fruited Chilean strawberry, Fragaria chiloensis (L.) Mill. Genet. Resour. Crop Evol. 2009, 56, 895–903. [Google Scholar] [CrossRef]
- Simirgiotis, M.J.; Schmeda-Hirschmann, G. Determination of phenolic composition and antioxidant activity in fruits, rhizomes and leaves of the white strawberry (Fragaria chiloensis spp. chiloensis form chiloensis) using HPLC-DAD–ESI-MS and free radical quenching techniques. J. Food Compos. Anal. 2010, 23, 545–553. [Google Scholar] [CrossRef]
- Simirgiotis, M.J.; Theoduloz, C.; Caligari, P.D.S.; Schmeda-Hirschmann, G. Comparison of phenolic composition and antioxidant properties of two native Chilean and one domestic strawberry genotypes. Food Chem. 2009, 113, 377–385. [Google Scholar] [CrossRef]
- Schmeda Hirschmann, G.; Simirgiotis, M.J.; Cheel, J. Chemistry of the Chilean Strawberry (Fragaria chiloensis spp. chiloensis). Genes Genomes Genom. 2011, 5, 85–90. [Google Scholar]
- Salvatierra, A.; Pimentel, P.; Moya-Leon, M.A.; Caligari, P.D.S.; Herrera, R. Comparison of transcriptional profiles of flavonoid genes and anthocyanin contents during fruit development of two botanical forms of Fragaria chiloensis ssp. chiloensis. Phytochemistry 2010, 71, 1839–1847. [Google Scholar] [CrossRef]
- Noriega, F.; Mardones, C.; Fischer, S.; García-Viguera, C.; Moreno, D.A.; López, M.D. Seasonal changes in white strawberry: Effect on aroma, phenolic compounds and its biological activity. J. Berry Res. 2021, 11, 103–118. [Google Scholar] [CrossRef]
- Barrios, S.; Lema, P.; Lareo, C. Modeling respiration rate of strawberry (cv. San Andreas) for modified atmosphere packaging design. Int. J. Food Prop. 2014, 17, 2039–2051. [Google Scholar] [CrossRef]
- Wu, X.; Han, W.; Yang, Z.Q.; Zhang, Y.X.; Zheng, Y.J. The difference in temperature between day and night affects the strawberry soluble sugar content by influencing the photosynthesis, respiration and sucrose phosphatase synthase. Hortic. Sci. 2021, 48, 174–182. [Google Scholar] [CrossRef]
- Wang, S.Y.; Camp, M.J. Temperatures after bloom affect plant growth and fruit quality of strawberry. Sci. Hortic. 2000, 85, 183–199. [Google Scholar] [CrossRef]
- Kim, S.-K.; Bae, R.N.; Na, H.; Ko, K.D.; Chun, C. Changes in fruit physicochemical characteristics by fruit clusters in June-bearing strawberry cultivars. Hortic. Sci. Technol. 2012, 30, 378–384. [Google Scholar] [CrossRef]
- FAWN. Florida Automated Weather Network. 2021. Available online: https://fawn.ifas.ufl.edu (accessed on 12 July 2021).
- Nunes, M.C.d.N. Color Atlas of Postharvest Quality of Fruits and Vegetables; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- Kelly, K.; do Nascimento Nunes, C.; Whitaker, V.M. A comparison of physical and chemical attributes of strawberry cultivars and advanced breeding selections from the University of Florida. Proc. Fla. State Hortic. Soc. 2016, 129, 185–189. [Google Scholar]
- Whitaker, V.M.; Chandler, C.K.; Santos, B.M.; Peres, N.; Nunes, M.C.d.N.; Plotto, A.; Sims, C.A. Winterstar™(‘FL 05-107’) strawberry. HortScience 2012, 47, 296–298. [Google Scholar] [CrossRef]
- Kelly, K.; Whitaker, V.M.; Nunes, M.C.d.N. Physicochemical characterization and postharvest performance of the new Sensation ® ‘Florida127’ strawberry compared to commercial standards. Sci. Hortic. 2016, 211, 283–294. [Google Scholar] [CrossRef]
- Nunes, M.; Brecht, J.K.; Morais, A.M.M.B.; Sargent, S.A. Physical and chemical quality characteristics of strawberries after storage are reduced by a short delay to cooling. Postharvest Biol. Technol. 1995, 6, 17–28. [Google Scholar] [CrossRef]
- Nunes, M.C.N.; Brecht, J.K.; Morais, A.M.B.; Sargent, S.A. Possible influences of water loss and polyphenol oxidase activity on anthocyanin content and discoloration in fresh ripe strawberry (cv. Oso Grande) during storage at 1 C. J. Food Sci. 2005, 70, S79–S84. [Google Scholar] [CrossRef]
- Abountiolas, M.; Kelly, K.; Yagiz, Y.; Li, Z.; Mahnken, G.; Borejsza-Wysocki, W.; Marshall, M.; Sims, C.A.; Peres, N.; Nunes, M.C.d.N. Sensory Quality, Physicochemical Attributes, Polyphenol Profiles, and Residual Fungicides in Strawberries from Different Disease-Control Treatments. J. Agric. Food Chem. 2018, 66, 6986–6996. [Google Scholar] [CrossRef]
- Whitaker, V.M.; Peres, N.A.; Osorio, L.F.; Fan, Z.; do Nascimento Nunes, M.C.; Plotto, A.; Sims, C.A. ‘Florida Brilliance’Strawberry. HortScience 2019, 54, 2073–2077. [Google Scholar] [CrossRef]
- Molinett, S.A.; Alfaro, J.F.; Sáez, F.A.; Elgueta, S.; Moya-León, M.A.; Figueroa, C.R. Postharvest treatment of hydrogen sulfide delays the softening of chilean strawberry fruit by downregulating the expression of key genes involved in pectin catabolism. Int. J. Mol. Sci. 2021, 22, 10008. [Google Scholar] [CrossRef] [PubMed]
- Moya-León, M.A.; Mattus-Araya, E.; Herrera, R. Molecular events occurring during softening of strawberry fruit. Front. Plant Sci. 2019, 10, 615. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, C.R.; Pimentel, P.; Gaete-Eastman, C.; Moya, M.; Herrera, R.; Caligari, P.D.S.; Moya-León, M.A. Softening rate of the Chilean strawberry (Fragaria chiloensis) fruit reflects the expression of polygalacturonase and pectate lyase genes. Postharvest Biol. Technol. 2008, 49, 210–220. [Google Scholar] [CrossRef]
- Figueroa, C.R.; Pimentel, P.; Dotto, M.C.; Civello, P.M.; Martínez, G.A.; Herrera, R.; Moya-León, M.A. Expression of five expansin genes during softening of Fragaria chiloensis fruit: Effect of auxin treatment. Postharvest Biol. Technol. 2009, 53, 51–57. [Google Scholar] [CrossRef]
- Figueroa, C.R.; Rosli, H.G.; Civello, P.M.; Martínez, G.A.; Herrera, R.; Moya-León, M.A. Changes in cell wall polysaccharides and cell wall degrading enzymes during ripening of Fragaria chiloensis and Fragaria× ananassa fruits. Sci. Hortic. 2010, 124, 454–462. [Google Scholar] [CrossRef]
- Nunes, M.C.N.; Brecht, J.K.; Morais, A.M.M.B.; Sargent, S.A. Physicochemical changes during strawberry development in the field compared with those that occur in harvested fruit during storage. J. Sci. Food Agric. 2006, 86, 180–190. [Google Scholar] [CrossRef]
- Nunes, M.; Morais, A.M.M.B.; Brecht, J.K.; Sargent, S.A. Fruit maturity and storage temperature influence response of strawberries to controlled atmospheres. J. Am. Soc. Hortic. Sci. 2002, 127, 836–842. [Google Scholar] [CrossRef]
- Singh, R.; Sharma, R.; Tyagi, S. Pre-harvest foliar application of calcium and boron influences physiological disorders, fruit yield and quality of strawberry (Fragaria× ananassa Duch.). Sci. Hortic. 2007, 112, 215–220. [Google Scholar] [CrossRef]
- Saavedra, G.M.; Figueroa, N.E.; Poblete, L.A.; Cherian, S.; Figueroa, C.R. Effects of preharvest applications of methyl jasmonate and chitosan on postharvest decay, quality and chemical attributes of Fragaria chiloensis fruit. Food Chem. 2016, 190, 448–453. [Google Scholar] [CrossRef]
- Figueroa, C.R.; Opazo, M.C.; Vera, P.; Arrigada, O.; Díaz, M.; Moya-León, M.A. Effect of postharvest treatment of calcium and auxin on cell wall composition and expression of cell wall-modifying genes in the Chilean strawberry (Fragaria chiloensis) fruit. Food Chem. 2012, 132, 2014–2022. [Google Scholar] [CrossRef]
- Cayo, Y.P.; Sargent, S.; Nunes, C.d.N.; Whitaker, V. Composition of Commercial Strawberry Cultivars and Advanced Selections as Affected by Season, Harvest, and Postharvest Storage. HortScience 2016, 51, 1134–1143. [Google Scholar] [CrossRef]
- Cordenunsi, B.R.; Oliveira do Nascimento, J.R.; Genovese, M.I.; Lajolo, F.M. Influence of cultivar on quality parameters and chemical composition of strawberry fruits grown in Brazil. J. Agric. Food Chem. 2002, 50, 2581–2586. [Google Scholar] [CrossRef] [PubMed]
- Forney, C.F.; Breen, P.J. Collection and characterization of phloem exudate from strawberry pedicels. HortScience 1985, 20, 413–414. [Google Scholar] [CrossRef]
- Kelly, K.; Yagiz, Y.; Li, Z.; Mahnken, G.; Borejsza-Wysocki, W.; Marshall, M.; Sims, C.A.; Peres, N.; Nunes, M.C.d.N. Sensory and Physicochemical Quality, Residual Fungicide Levels and Microbial Load in ‘Florida Radiance’Strawberries from Different Disease Control Treatments Exposed to Simulated Supply Chain Conditions. Foods 2021, 10, 1442. [Google Scholar] [CrossRef]
- Kim, S.K.; Kim, D.S.; Kim, D.Y.; Chun, C. Variation of bioactive compounds content of 14 oriental strawberry cultivars. Food Chem. 2015, 184, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Cervantes, L.; Ariza, M.T.; Gómez-Mora, J.A.; Miranda, L.; Medina, J.J.; Soria, C.; Martínez-Ferri, E. Light exposure affects fruit quality in different strawberry cultivars under field conditions. Sci. Hortic. 2019, 252, 291–297. [Google Scholar] [CrossRef]
- Gasperotti, M.; Masuero, D.; Guella, G.; Palmieri, L.; Martinatti, P.; Pojer, E.; Mattivi, F.; Vrhovsek, U. Evolution of ellagitannin content and profile during fruit ripening in Fragaria spp. J. Agric. Food Chem. 2013, 61, 8597–8607. [Google Scholar] [CrossRef]
- Cheel, J.; Theoduloz, C.; Rodríguez Caligari, P.D.S.; Schmeda-Hirschmann, G. Free radical scavenging activity and phenolic content in achenes and thalamus from Fragaria chiloensis ssp. chiloensis, F. vesca and F. x ananassa cv. Chandler. Food Chem. 2007, 102, 36–44. [Google Scholar] [CrossRef]
- Cheel, J.; Theoduloz, C.; Rodríguez, J.; Saud, G.; Caligari, P.D.S.; Schmeda-Hirschmann, G. E-cinnamic acid derivatives and phenolics from Chilean strawberry fruits, Fragaria chiloensis ssp. chiloensis. J. Agric. Food Chem. 2005, 53, 8512–8518. [Google Scholar] [CrossRef]
- Wang, X.; Wu, L.; Qiu, J.; Qian, Y.; Wang, M. Comparative Metabolomic Analysis of the Nutritional Aspects from Ten Cultivars of the Strawberry Fruit. Foods 2023, 12, 1153. [Google Scholar] [CrossRef]
- Thomas-Valdés, S.; Marzban, G.; Herndl, A.; Laimer, M.; Mezzetti, B.; Battino, M. Changes in polyphenol composition and bioactivity of the native Chilean white strawberry (Fragaria chiloensis spp. chiloensis f. chiloensis) after in vitro gastrointestinal digestion. Food Res. Int. 2018, 105, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Tulipani, S.; Marzban, G.; Herndl, A.; Laimer, M.; Mezzetti, B.; Battino, M. Influence of environmental and genetic factors on health-related compounds in strawberry. Food Chem. 2011, 124, 906–913. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, H.; Yang, Y.; Li, M.; Zhang, Y.; Liu, J.; Dong, J.; Li, J.; Butelli, E.; Xue, Z.; et al. The control of red colour by a family of MYB transcription factors in octoploid strawberry (Fragaria× ananassa) fruits. Plant Biotechnol. J. 2020, 18, 1169–1184. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Song, P.; Zhang, X.; Li, G.; Hu, P.; Aslam, A.; Zhao, X.; Zhou, H. Identification of candidate genes influencing anthocyanin biosynthesis during the development and ripening of red and white strawberry fruits via comparative transcriptome analysis. Peer J. 2021, 9, e10739. [Google Scholar] [CrossRef]
- Nour, V.; Trandafir, I.; Cosmulescu, S. Antioxidant Compounds, Nutritional Quality and Colour of Two Strawberry Genotypes from Fragaria × Ananassa. Erwerbs Obstbau 2017, 59, 123–131. [Google Scholar] [CrossRef]
- Zhao, J.; Liu, J.; Wang, F.; Wang, S.; Feng, H.; Xie, X.; Hao, F.; Zhang, L.; Fang, C. Volatile constituents and ellagic acid formation in strawberry fruits of selected cultivars. Food Res. Int. 2020, 138, 109767. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Harvest Date | Temperature (°C) | Rainfall (mm) | Daily UV Index (Max AVG) | ||
|---|---|---|---|---|---|
| Max | Min | AVG | |||
| 18 January | 17.8 | 6.1 | 12.5 | 0 | 4 |
| 10 February | 27.2 | 18.9 | 21.1 | 0 | 5 |
| 1 March | 28.3 | 21.7 | 24.3 | 0 | 6 |
| Compound | Brilliance | Pearl | ||
|---|---|---|---|---|
| Mean FW ± SE | Mean DW ± SE | Mean FW ± SE | Mean DW ± SE | |
| Acidity (%) | 0.78 ± 0.04 | 11.36 ± 0.40 | 0.65 ± 0.06 | 8.39 ± 0.33 |
| SSC (%) | 7.17 ± 0.22 | 95.74 ± 2.13 | 7.97 ± 0.53 | 96.47 ± 1.82 |
| Sugars (g 100 g−1) | ||||
| Sucrose | 0.26 ± 0.05 | 3.92 ± 1.10 | 0.04 ± 0.01 | 0.64 ± 0.21 |
| Glucose | 2.03 ± 0.11 | 29.45 ± 2.15 | 2.67 ± 0.17 | 34.19 ± 1.33 |
| Fructose | 2.10 ± 0.14 | 30.44 ± 2.67 | 2.96 ± 0.18 | 38.18 ± 1.28 |
| Total Sugars | 4.39 ± 0.20 | 63.81 ± 3.87 | 5.67 ± 0.34 | 73.02 ± 2.45 |
| Bioactive compounds (mg 100 g−1) | ||||
| Total ascorbic acid | 61.78 ± 1.54 | 900.53 ± 21.69 | 74.63 ± 2.27 | 1015.83 ± 40.25 |
| Total phenolics | 132.97 ± 4.49 | 1931.06 ± 95.10 | 114.51 ± 1.83 | 1605.66 ± 110.33 |
| Total anthocyanins | 11.91 ± 0.44 | 173.67 ± 9.77 | 0.83 ± 0.03 | 12.10 ± 1.37 |
| Polyphenols (mg 100 g−1) | ||||
| Cyanidin | 0.98 ± 0.09 | 14.34 ± 1.17 | 0.82 ± 0.02 | 11.46 ± 0.75 |
| Pelargonidin | 7.18 ± 1.24 | 105.94 ± 19.37 | 3.40 ± 0.18 | 47.32 ± 3.73 |
| Cyanidin-3-glucoside | 0.12 ± 0.02 | 1.72 ± 0.51 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Pelargonidin 3-glucoside | 5.13 ± 2.45 | 76.11 ± 36.30 | 1.81 ± 0.33 | 21.85 ± 3.08 |
| Kaempferol 3-glucoside | 1.27 ± 0.17 | 17.80 ± 2.52 | 7.32 ± 0.28 | 102.21 ± 6.91 |
| Kaempferol | 1.06 ± 0.09 | 15.61 ± 1.34 | 0.93 ± 0.04 | 12.66 ± 0.55 |
| Quercetin | 3.51 ± 0.19 | 7.74 ± 2.99 | 4.62 ± 0.02 | 0.87 ± 0.27 |
| Quercetin 3-glucoside | 0.52 ± 0.31 | 41.39 ± 4.46 | 7.86 ± 2.23 | 86.22 ± 31.76 |
| Myricetin | 1.39 ± 0.14 | 20.10 ± 1.79 | 1.70 ± 0.16 | 22.58 ± 1.85 |
| Catechin | 2.01 ± 0.59 | 29.63 ± 8.82 | 2.03 ± 0.67 | 30.43 ± 9.53 |
| Epicatechin | 8.31 ± 3.46 | 122.83 ± 51.23 | 1.19 ± 0.17 | 17.33 ± 2.88 |
| Caffeic acid | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| p-Coumaric acid | 1.16 ± 0.14 | 15.65 ± 1.89 | 0.04 ± 0.02 | 0.66 ± 0.32 |
| Ferulic acid | 3.60 ± 0.48 | 53.28 ± 6.25 | 1.82 ± 0.60 | 25.89 ± 8.54 |
| Gallic acid | 2.32 ± 0.70 | 34.09 ± 10.33 | 3.90 ± 0.78 | 60.89 ± 11.52 |
| Chlorogenic acid | 0.19 ± 0.00 | 2.71 ± 0.04 | 0.24 ± 0.04 | 3.47 ± 0.65 |
| Ellagic acid | 1.15 ± 0.11 | 16.71 ± 1.51 | 1.03 ± 0.13 | 14.34 ± 1.96 |
| Polyphenols | Day 0 | Day 9 | ||
|---|---|---|---|---|
| Brilliance | Pearl | Brilliance | Pearl | |
| Anthocyanidins | ||||
| Cyanidin | 15.77 ± 3.52 a | 8.77 ± 0.10 b | 12.16 ± 1.66 a | 4.85 ± 0.13 b |
| Pelargonidin | 88.11 ± 5.12 a | 37.09 ± 3.50 b | 24.68 ± 2.98 a | 28.07 ± 1.18 a |
| Anthocyanins | ||||
| Cyanidin 3-glucoside | 3.81 ± 2.85 a | 0.00 ± 0.00 b | 5.20 ± 2.60 a | 0.00 ± 0.00 b |
| Pelargonidin 3-glucoside | 32.42 ± 0.29 a | 0.00 ± 0.00 b | 10.11 ± 0.20 a | 0.00 ± 0.00 b |
| Flavonols | ||||
| Kaempferol 3-glucoside | 16.01 ± 2.49 b | 79.105 ± 9.79 a | 12.73 ± 1.60 b | 24.75 ± 1.63 a |
| Kaempferol | 12.31 ± 0.28 a | 10.81 ± 1.34 a | 11.59 ± 0.16 a | 5.18 ± 0.22 b |
| Quercetin 3-glucoside | 41.06 ± 2.91 a | 16.78 ± 5.85 b | 9.25 ± 0.14 a | 5.01 ± 1.06 a |
| Quercetin | 5.21 ± 1.44 a | 0.82 ± 0.81 b | 4.29 ± 0.32 a | 0.23 ± 0.02 b |
| Myricetin | 24.35 ± 2.40 a | 22.76 ± 2.10 a | 21.35 ± 7.22 a | 5.84 ± 0.07 b |
| Flavanols | ||||
| Catechin | 13.29 ± 0.01 a | 10.46 ± 0.02 a | 6.76 ± 0.12 a | 8.18 ± 0.59 a |
| Epicatechin | 22.23 ± 0.09 a | 10.58 ± 0.16 b | 46.25 ± 2.16 a | 10.73 ±1.47 b |
| Phenolic acids | ||||
| Caffeic acid | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| p-Coumaric acid | 22.18 ± 0.91 a | 0.00 ± 0.00 b | 26.39 ± 4.60 a | 0.00 ± 0.00 b |
| Ferulic acid | 73.11 ± 4.23 a | 13.96 ± 1.94 b | 7.92 ± 0.10 a | 3.85 ± 0.54 b |
| Gallic acid | 14.10 ± 0.13 a | 14.82 ± 1.68 a | 6.42 ± 0.22 a | 5.50 ± 0.08 b |
| Chlorogenic acid | 2.55 ± 0.01 a | 2.01 ± 0.01 a | 10.26 ± 0.22 a | 3.38 ± 1.39 b |
| Hydrolyzable tannins | ||||
| Ellagic acid | 18.46 ± 4.47 a | 10.42 ± 0.74 b | 18.95 ± 3.69 a | 4.65 ± 0.05 b |
| Polyphenols | Day 0 | Day 9 | ||
|---|---|---|---|---|
| Brilliance | Pearl | Brilliance | Pearl | |
| Anthocyanidins | ||||
| Cyanidin | 11.83 ± 0.01 a | 11.44 ± 0.13 a | 8.15 ± 1.25 a | 6.76 ± 0.57 a |
| Pelargonidin | 54.55 ± 4.23 a | 45.54 ± 5.97 b | 152.30 ± 18.48 a | 27.89 ± 4.31 a |
| Anthocyanins | ||||
| Cyanidin 3-glucoside | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| Pelargonidin 3-glucoside | 215.63 ± 15.54 a | 17.66 ± 0.23 b | 0.62 ± 0.02 a | 1.22 ± 0.71 a |
| Flavonols | ||||
| Kaempferol 3-glucoside | 23.07 ± 4.24 b | 100.15 ± 8.43 a | 6.72 ± 0.69 a | 5.00 ± 0.86 a |
| Kaempferol | 20.44 ± 1.06 a | 13.48 ± 0.19 b | 7.92 ± 0.94 a | 6.32 ± 0.45 a |
| Quercetin 3-glucoside | 50.56 ± 4.70 b | 194.35 ± 0.38 a | 81.98 ± 9.62 a | 4.42 ± 0.84 b |
| Quercetin | 0.56 ± 0.06 a | 0.83 ± 0.28 a | 13.24 ± 1.89 a | 4.18 ± 1.01 b |
| Myricetin | 19.75 ± 1.58 a | 21.90 ± 0.36 a | 7.94 ± 0.75 a | 6.59 ± 0.32 a |
| Flavanols | ||||
| Catechin | 60.90 ± 10.91 a | 59.04 ± 9.70 a | 6.08 ± 0.23 a | 8.61 ± 1.88 a |
| Epicatechin | 317.50 ± 33.58 a | 17.02 ± 3.02 b | 5.69 ± 1.52 a | 6.49 ± 1.01 a |
| Phenolic acids | ||||
| Caffeic acid | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| p-Coumaric acid | 10.62 ± 3.27 a | 1.30 ± 0.55 b | 16.0 ± 8.34 a | 2.47 ± 0.40 b |
| Ferulic acid | 37.58 ± 6.01 b | 53.53 ± 10.62 a | 48.99 ± 5.71 a | 4.85 ± 0.51 b |
| Gallic acid | 74.10 ± 0.93 b | 92.61 ± 2.37 a | 6.68 ± 0.72 a | 7.42 ± 1.10 a |
| Chlorogenic acid | 2.74 ± 0.01 a | 4.99 ± 1.00 a | 1.17 ± 0.04 b | 4.85 ± 0.84 a |
| Hydrolyzable tannins | ||||
| Ellagic acid | 13.64 ± 0.27 a | 18.57 ± 5.38 a | 7.24 ± 0.89 a | 5.44 ± 0.06 a |
| Polyphenols | Day 0 | Day 9 | ||
|---|---|---|---|---|
| Brilliance | Pearl | Brilliance | Pearl | |
| Anthocyanidins | ||||
| Cyanidin | 15.4 ± 1.37 a | 13.26 ± 1.44 a | 14.28 ± 5.08 a | 6.46 ± 0.61 b |
| Pelargonidin | 175.15 ± 7.34 a | 55.91 ± 3.66 b | 21.42 ± 3.18 a | 24.11 ± 3.33 a |
| Anthocyanins | ||||
| Cyanidin 3-glucoside | 1.36 ± 0.66 a | 0.01 ± 0.00 b | 1.19 ± 0.19 b | 3.42 ± 0.76 a |
| Pelargonidin 3-glucoside | 12.70 ± 0.56 a | 18.99 ± 2.58 a | 10.93 ± 0.93 a | 1.68 ± 0.99 b |
| Flavonols | ||||
| Kaempferol 3-glucoside | 14.33 ± 1.33 b | 119.68 ± 4.78 a | 20.22 ± 3.58 b | 54.27 ± 1.54 a |
| Kaempferol | 14.08 ± 0.20 a | 13.08 ± 0.03 a | 7.40 ± 1.68 a | 5.65 ± 0.42 a |
| Quercetin 3-glucoside | 32.54 ± 11.60 a | 24.37 ± 5.32 b | 5.84 ± 1.18 b | 9.85 ± 1.92 a |
| Quercetin | 17.44 ± 3.81 a | 0.93 ± 0.36 b | 14.05 ± 3.61 a | 2.82 ± 0.02 b |
| Myricetin | 16.20 ± 1.49 b | 23.14 ± 1.34 a | 6.23 ± 0.18 b | 49.87 ± 3.28 a |
| Flavanols | ||||
| Catechin | 14.71 ± 0.14 a | 15.14 ± 2.52 a | 6.28 ± 0.93 a | 8.59 ± 0.01 a |
| Epicatechin | 28.78 ± 6.39 a | 22.14 ± 7.28 a | 15.02 ± 0.06 b | 31.22 ± 9.48 a |
| Phenolic acids | ||||
| Caffeic acid | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| p-Coumaric acid | 14.16 ± 1.80 a | 0.46 ± 0.01 b | 15.56 ± 1.40 a | 0.57 ± 0.14 b |
| Ferulic acid | 49.16 ± 4.73 a | 6.22 ± 0.94 b | 74.06 ± 11.33 a | 5.96 ± 1.05 b |
| Gallic acid | 14.11 ± 0.55 b | 59.89 ± 5.46 a | 5.05 ± 0.15 a | 5.53 ± 0.13 a |
| Chlorogenic acid | 2.83 ± 0.02 a | 2.91 ± 0.01 a | 2.80 ± 1.14 a | 1.14 ± 0.03 a |
| Hydrolyzable tannins | ||||
| Ellagic acid | 18.04 ± 2.36 a | 12.73 ± 0.25 a | 17.73 ± 1.82 a | 6.20 ± 0.17 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smith, A.N.; do Nascimento Nunes, M.C. Physicochemical Quality, Polyphenol Profiles, and Postharvest Performance of Florida Pearl® ‘FL 16.78-109’ White Strawberries Compared to the Red Cultivar ‘Florida Brilliance’. Foods 2023, 12, 3143. https://doi.org/10.3390/foods12173143
Smith AN, do Nascimento Nunes MC. Physicochemical Quality, Polyphenol Profiles, and Postharvest Performance of Florida Pearl® ‘FL 16.78-109’ White Strawberries Compared to the Red Cultivar ‘Florida Brilliance’. Foods. 2023; 12(17):3143. https://doi.org/10.3390/foods12173143
Chicago/Turabian StyleSmith, Alyssa Nicole, and Maria Cecilia do Nascimento Nunes. 2023. "Physicochemical Quality, Polyphenol Profiles, and Postharvest Performance of Florida Pearl® ‘FL 16.78-109’ White Strawberries Compared to the Red Cultivar ‘Florida Brilliance’" Foods 12, no. 17: 3143. https://doi.org/10.3390/foods12173143
APA StyleSmith, A. N., & do Nascimento Nunes, M. C. (2023). Physicochemical Quality, Polyphenol Profiles, and Postharvest Performance of Florida Pearl® ‘FL 16.78-109’ White Strawberries Compared to the Red Cultivar ‘Florida Brilliance’. Foods, 12(17), 3143. https://doi.org/10.3390/foods12173143