

Non-Saccharomyces Yeasts from Organic Vineyards as Spontaneous Fermentation Agents

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Yeasts from Spontaneous Fermentations
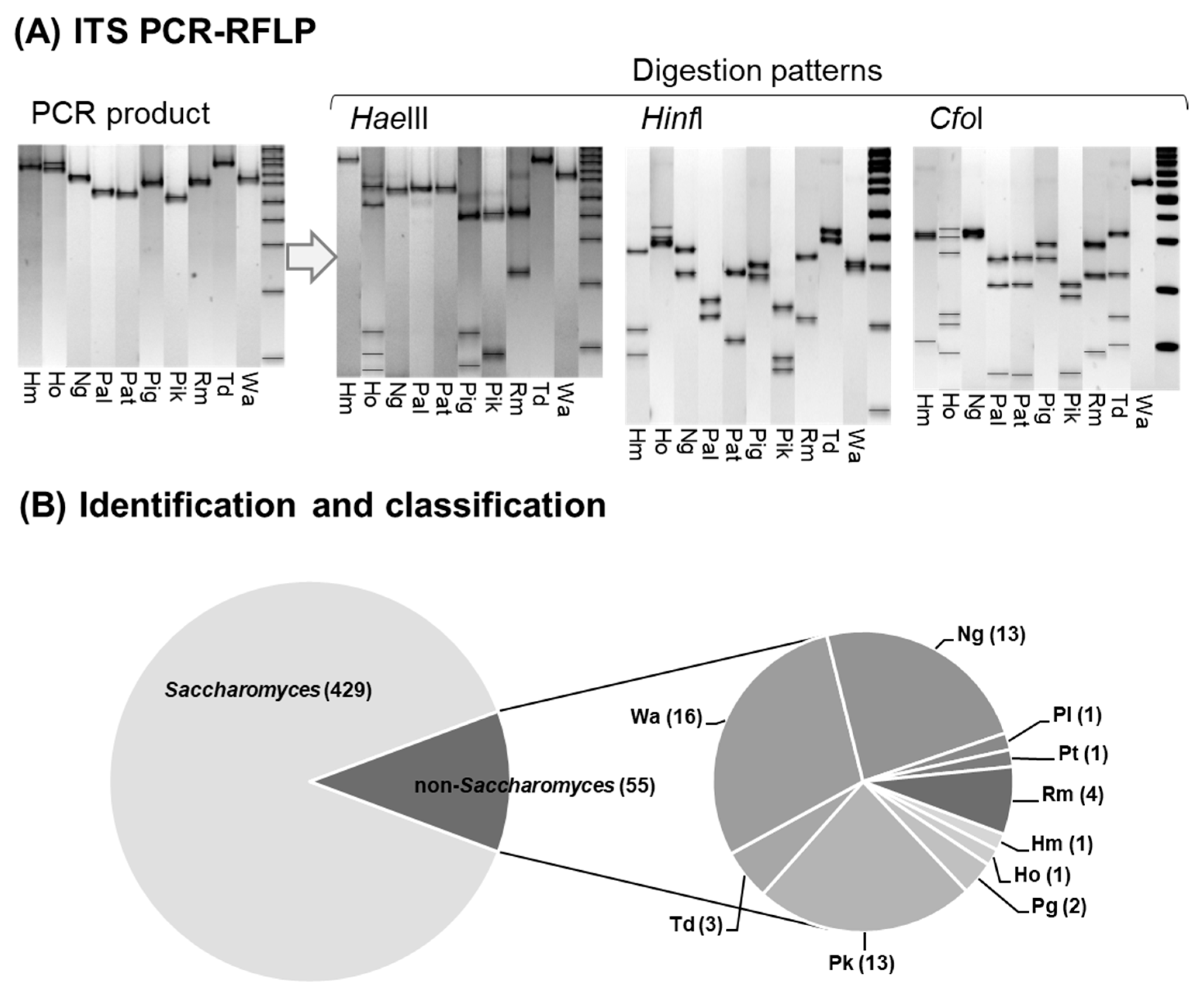
2.2. Molecular Identification of the Isolates
2.3. Enzymatic Activities
2.3.1. β-Glucosidase Activity
2.3.2. Protease Activity
2.3.3. β-Glucanase Activity
2.3.4. β-Lyase Activity
2.4. Fermentation Conditions
2.5. Statistical Analysis
3. Results
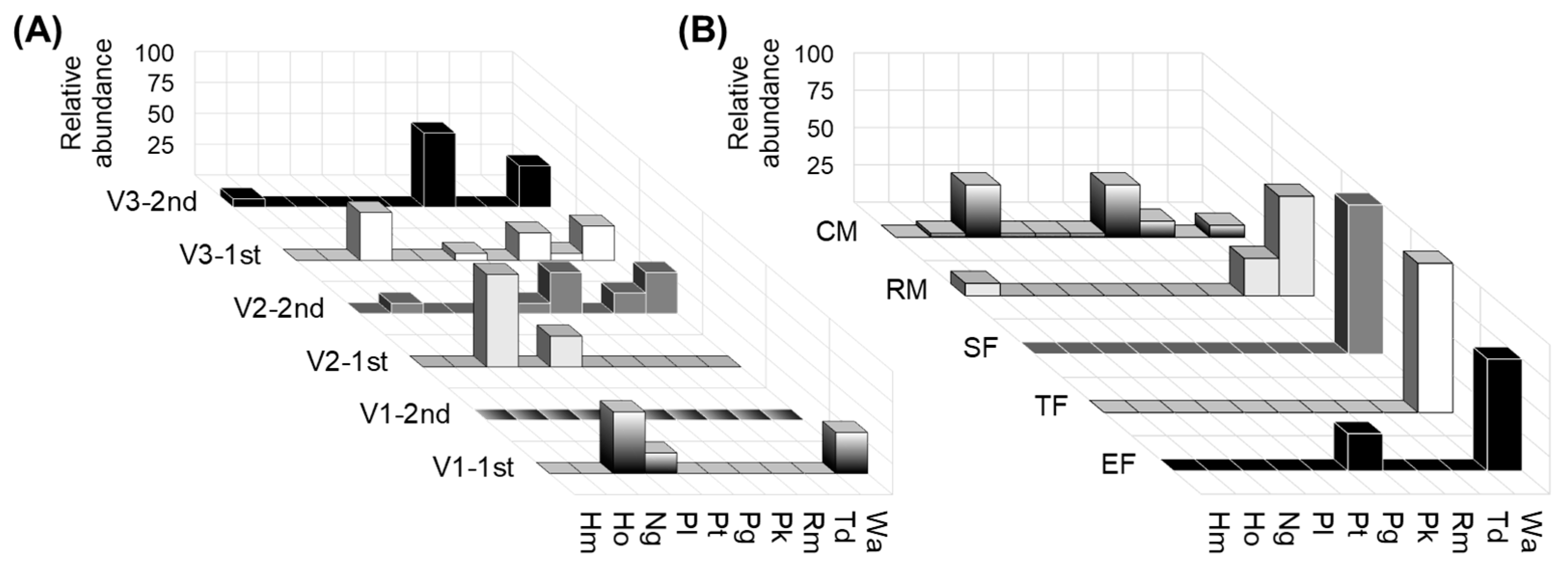
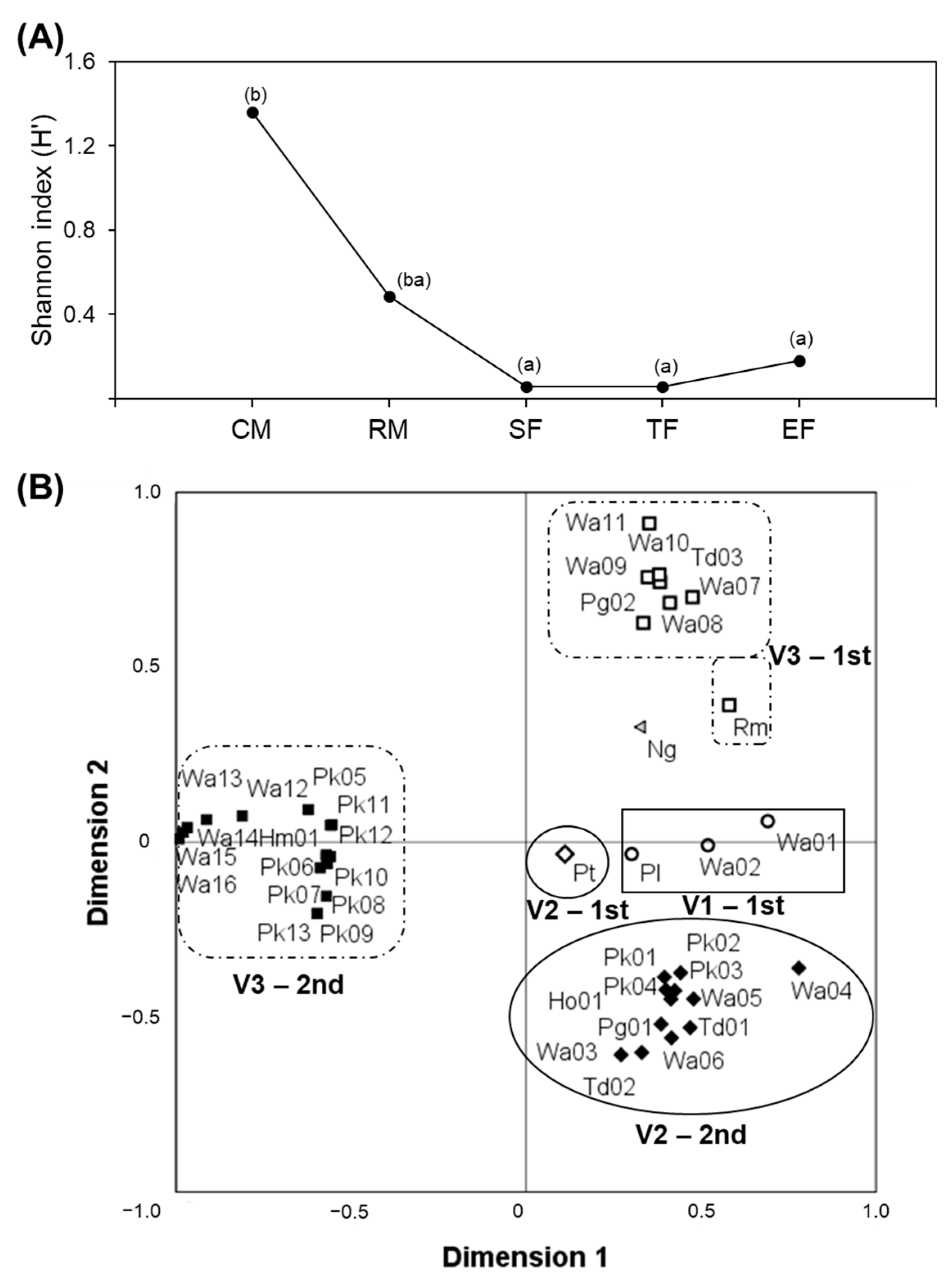
3.1. Analysis of Biodiversity of Non-Saccharomyces Populations
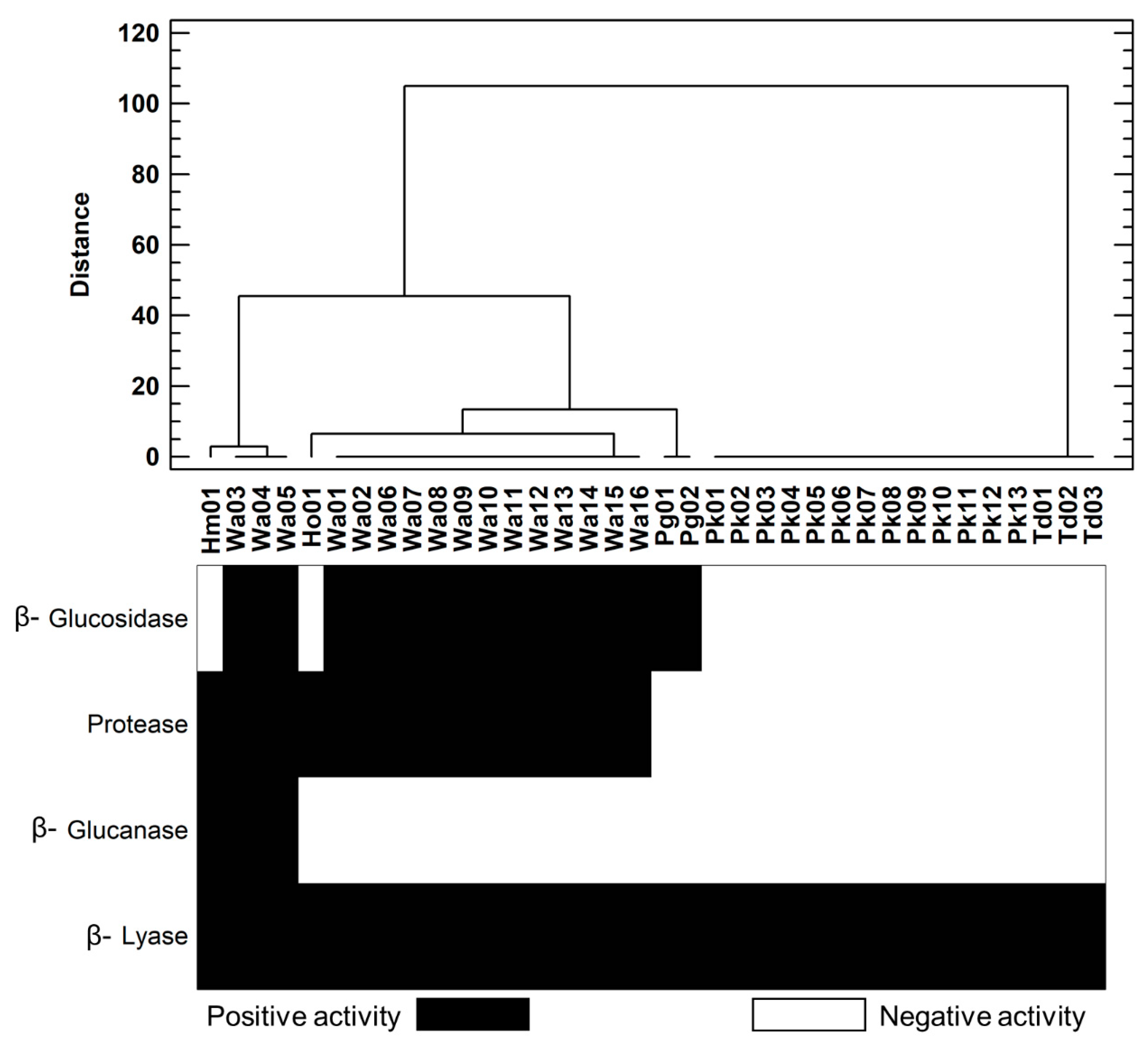
3.2. Enzymatic Profile of Yeast Population
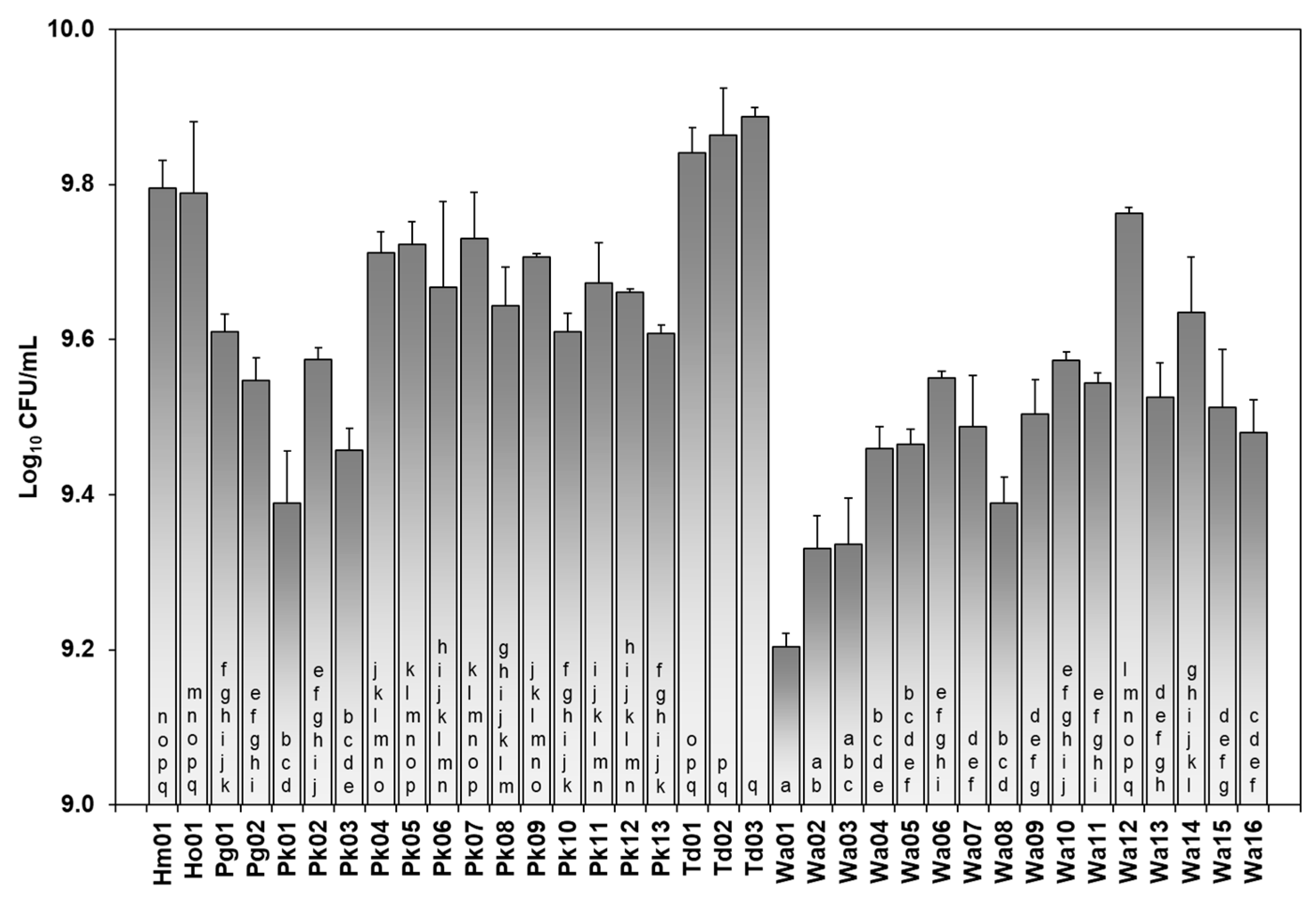
3.3. Fermentation Kinetics of Non-Saccharomyces Isolates
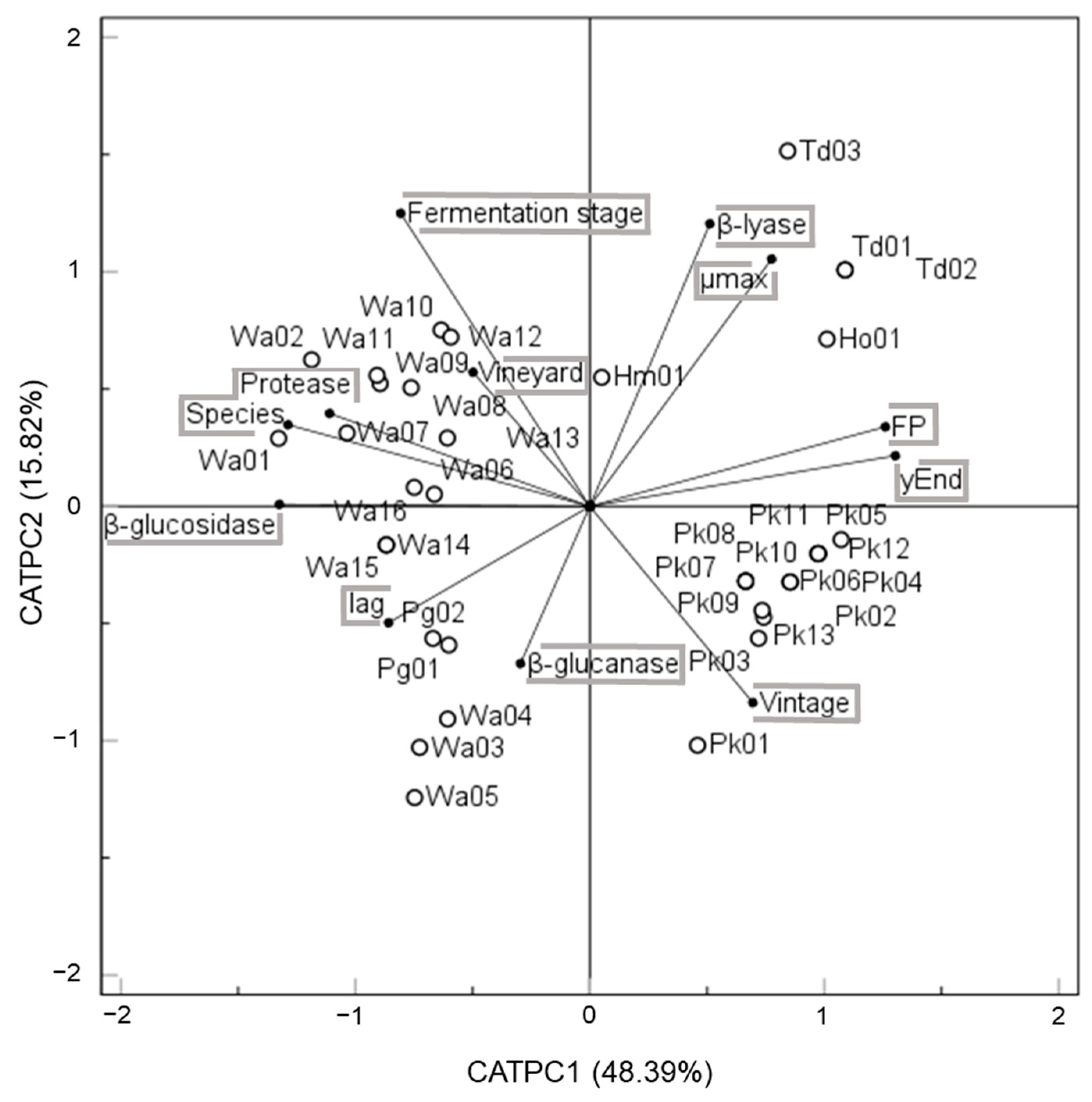
3.4. Screening of Oenological Traits
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not Your Ordinary Yeast: Non-Saccharomyces Yeasts in Wine Production Uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef]
- Borren, E.; Tian, B. The Important Contribution of Non-Saccharomyces Yeasts to the Aroma Complexity of Wine: A Review. Foods 2021, 10, 13. [Google Scholar] [CrossRef]
- Mas, A.; Portillo, M.C. Strategies for Microbiological Control of the Alcoholic Fermentation in Wines by Exploiting the Microbial Terroir Complexity: A Mini-Review. Int. J. Food Microbiol. 2022, 367, 109592. [Google Scholar] [CrossRef]
- Escribano, R.; González-Arenzana, L.; Portu, J.; Garijo, P.; López-Alfaro, I.; López, R.; Santamaría, P.; Gutiérrez, A.R. Wine Aromatic Compound Production and Fermentative Behaviour within Different Non-Saccharomyces Species and Clones. J. Appl. Microbiol. 2018, 124, 1521–1531. [Google Scholar] [CrossRef]
- Escribano, R.; González-Arenzana, L.; Garijo, P.; Berlanas, C.; López Alfaro, I.; López, R.; Gutiérrez, A.R.; Santamaría, P. Screening of Enzymatic Activities within Different Enological Non-Saccharomyces Yeasts. J. Food Sci. Technol. 2017, 54, 1555–1564. [Google Scholar] [CrossRef]
- Ciani, M.; Comitini, F. Non-Saccharomyces Wine Yeasts Have a Promising Role in Biotechnological Approaches to Winemaking. Ann. Microbiol. 2011, 61, 25–32. [Google Scholar] [CrossRef]
- Padilla, B.; Gil, J.V.; Manzanares, P. Past and Future of Non-Saccharomyces Yeasts: From Spoilage Microorganisms to Biotechnological Tools for Improving Wine Aroma Complexity. Front. Microbiol. 2016, 7, 411. [Google Scholar] [CrossRef]
- Benito, A.; Calderón, F.; Benito, S. The Influence of Non-Saccharomyces Species on Wine Fermentation Quality Parameters. Fermentation 2019, 5, 54. [Google Scholar] [CrossRef]
- Belda, I.; Ruiz, J.; Beisert, B.; Navascués, E.; Marquina, D.; Calderón, F.; Rauhut, D.; Benito, S.; Santos, A. Influence of Torulaspora Delbrueckii in Varietal Thiol (3-SH and 4-MSP) Release in Wine Sequential Fermentations. Int. J. Food Microbiol. 2017, 257, 183–191. [Google Scholar] [CrossRef]
- Binati, R.L.; Lemos Junior, W.J.F.; Luzzini, G.; Slaghenaufi, D.; Ugliano, M.; Torriani, S. Contribution of Non-Saccharomyces Yeasts to Wine Volatile and Sensory Diversity: A Study on Lachancea Thermotolerans, Metschnikowia Spp. and Starmerella Bacillaris Strains Isolated in Italy. Int. J. Food Microbiol. 2020, 318, 108470. [Google Scholar] [CrossRef]
- Canonico, L.; Comitini, F.; Ciani, M.; Solomon, M.; Varela, C. Volatile Profile of Reduced Alcohol Wines Fermented with Selected Non-Saccharomyces Yeasts under Different Aeration Conditions. Food Microbiol. 2019, 84, 103247. [Google Scholar] [CrossRef]
- Roudil, L.; Russo, P.; Berbegal, C.; Albertin, W.; Spano, G.; Capozzi, V. Non-Saccharomyces Commercial Starter Cultures: Scientific Trends, Recent Patents and Innovation in the Wine Sector. Recent Pat. Food. Nutr. Agric. 2020, 11, 27–39. [Google Scholar] [CrossRef]
- Bagheri, B.; Zambelli, P.; Vigentini, I.; Bauer, F.F.; Setati, M.E. Investigating the Effect of Selected Non-Saccharomyces Species on Wine Ecosystem Function and Major Volatiles. Front. Bioeng. Biotechnol. 2018, 6, 169. [Google Scholar] [CrossRef]
- Ciani, M.; Comitini, F.; Mannazzu, I.; Domizio, P. Controlled Mixed Culture Fermentation: A Newperspective on the Use of Non-Saccharomyces Yeasts in Winemaking. FEMS Yeast Res. 2010, 10, 123–133. [Google Scholar] [CrossRef]
- Zilelidou, E.A.; Nisiotou, A. Understanding Wine through Yeast Interactions. Microorganisms 2021, 9, 1620. [Google Scholar] [CrossRef]
- Englezos, V.; Jolly, N.P.; Di Gianvito, P.; Rantsiou, K.; Cocolin, L. Microbial Interactions in Winemaking: Ecological Aspects and Effect on Wine Quality. Trends Food Sci. Technol. 2022, 127, 99–113. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Collins, T.S.; Masarweh, C.; Allen, G.; Heymann, H.; Ebeler, S.E.; Mills, D.A. Associations among Wine Grape Microbiome, Metabolome, and Fermentation Behavior Suggest Microbial Contribution to Regional Wine Characteristics. MBio 2016, 7, e00631-16. [Google Scholar] [CrossRef]
- Liu, D.; Legras, J.L.; Zhang, P.Z.; Chen, D.L.; Howell, K. Diversity and Dynamics of Fungi during Spontaneous Fermentations and Association with Unique Aroma Profiles in Wine. Int. J. Food Microbiol. 2021, 338, 11. [Google Scholar] [CrossRef]
- López-Enríquez, L.; Vila-Crespo, J.; Rodríguez-Nogales, J.M.; Fernández-Fernández, E.; Ruipérez, V. Screening and Enzymatic Evaluation of Saccharomyces Cerevisiae Populations from Spontaneous Fermentation of Organic Verdejo Wines. Foods 2022, 11, 3448. [Google Scholar] [CrossRef]
- Querol, A.; Barrio, E.; Huerta, T.; Ramon, D. Molecular Monitoring of Wine Fermentations Conducted by Active Dry Yeast Strains. Appl. Environ. Microbiol. 1992, 58, 2948–2953. [Google Scholar] [CrossRef]
- Esteve-Zarzoso, B.; Belloch, C.; Uruburu, F.; Querol, A. Identification of Yeasts by RFLP Analysis of the 5.8S RRNA Gene and the Two Ribosomal Internal Transcribed Spacers. Int. J. Syst. Bacteriol. 1999, 49, 329–337. [Google Scholar] [CrossRef]
- Suárez-Lepe, J.A.; Íñigo, B. Microbiología Enológica: Fundamentos de Vinificación, 3rd ed.; Mundi-Prensa: Madrid, Spain, 2004; ISBN 9788484761846. [Google Scholar]
- Belda, I.; Ruiz, J.; Alastruey-Izquierdo, A.; Navascués, E.; Marquina, D.; Santos, A. Unraveling the Enzymatic Basis of Wine “Flavorome”: A Phylo-Functional Study of Wine Related Yeast Species. Front. Microbiol. 2016, 7, 12. [Google Scholar] [CrossRef]
- Baranyi, J.; Roberts, T.A. A Dynamic Approach to Predicting Bacterial Growth in Food. Int. J. Food Microbiol. 1994, 23, 277–294. [Google Scholar] [CrossRef]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Champaign, IL, USA, 1949. [Google Scholar]
- Castrillo, D.; Neira, N.; Blanco, P. Saccharomyces Cerevisiae Strain Diversity Associated with Spontaneous Fermentations in Organic Wineries from Galicia (NW Spain). Fermentation 2020, 6, 89. [Google Scholar] [CrossRef]
- Paliy, O.; Shankar, V. Application of Multivariate Statistical Techniques in Microbial Ecology. Mol. Ecol. 2016, 25, 1032–1057. [Google Scholar] [CrossRef]
- Ramette, A. Multivariate Analyses in Microbial Ecology. Fems Microbiol. Ecol. 2007, 62, 142–160. [Google Scholar] [CrossRef]
- Hammer, O.; Harper, D.; Ryan, P. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Kurtzman, C.P.; Fell, J.W.; Boekhout, T. (Eds.) Yeasts: A Taxonomic Study, 5th ed.; Elsevier Science & Technology, ProQuest Ebook Central: Oxford, UK, 2011; Volume 1–3, ISBN 978-0-123-84868-0. [Google Scholar]
- Varela, C.; Borneman, A.R. Yeasts Found in Vineyards and Wineries. Yeast 2017, 34, 111–128. [Google Scholar] [CrossRef]
- Liu, D.; Zhang, P.Z.; Chen, D.L.; Howell, K. From the Vineyard to the Winery: How Microbial Ecology Drives Regional Distinctiveness of Wine. Front. Microbiol. 2019, 10, 13. [Google Scholar] [CrossRef]
- Barata, A.; Malfeito-Ferreira, M.; Loureiro, V. The Microbial Ecology of Wine Grape Berries. Int. J. Food Microbiol. 2012, 153, 243–259. [Google Scholar] [CrossRef]
- Alexandre, H. Wine Yeast Terroir: Separating the Wheat from the Chaff—For an Open Debate. Microorganisms 2020, 8, 787. [Google Scholar] [CrossRef]
- Belda, I.; Ruiz, J.; Navascués, E.; Marquina, D.; Santos, A. Improvement of Aromatic Thiol Release through the Selection of Yeasts with Increased Beta-Lyase Activity. Int. J. Food Microbiol. 2016, 225, 1–8. [Google Scholar] [CrossRef]
- Gao, P.P.; Peng, S.; Sam, F.E.; Zhu, Y.T.; Liang, L.H.; Li, M.; Wang, J. Indigenous Non-Saccharomyces Yeasts with Beta-Glucosidase Activity in Sequential Fermentation with Saccharomyces Cerevisiae: A Strategy to Improve the Volatile Composition and Sensory Characteristics of Wines. Front. Microbiol. 2022, 13, 845837. [Google Scholar] [CrossRef]
- Lin, M.; Boss, P.; Walker, M.; Sumby, K.; Grbin, P.; Jiranek, V. Evaluation of Indigenous Non-Saccharomyces Yeasts Isolated from a South Australian Vineyard for Their Potential as Wine Starter Cultures. Int. J. Food Microbiol. 2020, 312, 108373. [Google Scholar] [CrossRef]
- Rocker, J.; Strub, S.; Ebert, K.; Grossmann, M. Usage of Different Aerobic Non-Saccharomyces Yeasts and Experimental Conditions as a Tool for Reducing the Potential Ethanol Content in Wines. Eur. Food Res. Technol. 2016, 242, 2051–2070. [Google Scholar] [CrossRef]
- Padilla, B.; Zulian, L.; Ferreres, A.; Pastor, R.; Esteve-Zarzoso, B.; Beltran, G.; Mas, A. Sequential Inoculation of Native Non-Saccharomyces and Saccharomyces Cerevisiae Strains for Wine Making. Front. Microbiol. 2017, 8, 1293. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | µmax (Days−1) | Lag (Days) | yEnd (g of Total CO2 Produced) | FP (Theoretical % vol. Ethanol) |
|---|---|---|---|---|
| Hanseniaspora meyeri | ||||
| Hm01 | 3.4 ± 0.8 | - | 39.7 ± 11.4 | 5.0 ± 1.4 |
| Hanseniaspora osmophila | ||||
| Ho01 | 9.6 ± 2.0 | - | 86.9 ± 12.5 | 10.9 ± 1.6 |
| Pichia guilliermondii | ||||
| Pg01 | 1.2 ± 0.1 a | 4.6 ± 0.2 a | 15.6 ± 1.6 a | 1.9 ± 0.2 a |
| Pg02 | 1.4 ± 0.1 a | 3.9 ± 0.7 a | 18.7 ± 1.0 a | 2.3 ± 0.1 a |
| Pichia kudriavzevii | ||||
| Pk01 | 1.8 ± 0.8 a | 1.7 ± 0.9 a | 34.3 ± 12.5 a | 4.3 ± 1.6 a |
| Pk02 | 2.4 ± 0.5 a | 0.7 ± 0.3 a | 39.4 ± 12.5 a | 4.9 ± 1.6 a |
| Pk03 | 2.8 ± 0.7 a | 0.2 ± 0.0 a | 47.4 ± 15.7 ab | 5.9 ± 2.0 ab |
| Pk04 | 4.1 ± 1.3 a | 1.9 ± 1.1 a | 64.7 ± 5.6 ab | 8.1 ± 0.7 ab |
| Pk05 | 3.2 ± 0.3 a | 1.0 ± 0.0 a | 62.2 ± 3.9 ab | 7.8 ± 0.5 ab |
| Pk06 | 3.2 ± 0.7 a | 1.6 ± 0.0 a | 68.0 ± 1.0 ab | 8.5 ± 0.0 ab |
| Pk07 | 2.6 ± 0.4 a | - | 45.9 ± 7.7 ab | 5.7 ± 1.0 ab |
| Pk08 | 3.2 ± 0.4 a | - | 51.6 ± 9.3 ab | 6.4 ± 1.2 ab |
| Pk09 | 2.7 ± 0.2 a | - | 51.3 ± 1.7 ab | 6.4 ± 0.2 ab |
| Pk10 | 3.5 ± 1.5 a | 2.0 ± 0.3 a | 75.7 ± 4.2 b | 9.5 ± 0.5 b |
| Pk11 | 3.1 ± 0.1 a | - | 54.6 ± 6.0 ab | 6.8 ± 0.8 ab |
| Pk12 | 3.3 ± 0.2 a | 0.9 ± 0.0 a | 67.9 ± 3.8 ab | 8.5 ± 0.5 ab |
| Pk13 | 3.6 ± 0.8 a | 2.8 ± 0.0 a | 63.5 ± 13.0 ab | 7.9 ± 1.6 ab |
| Torulaspora delbruckii | ||||
| Td01 | 6.1 ± 0.4 a | - | 70.6 ± 8.7 a | 8.8 ± 1.1 a |
| Td02 | 6.9 ± 1.3 a | - | 81.0 ± 8.3 a | 10.1 ± 1.0 a |
| Td03 | 6.9 ± 0.9 a | - | 86.0 ± 10.6 a | 10.8 ± 1.3 a |
| Wickerhamomyces anomalus | ||||
| Wa01 | 1.8 ± 0.1 a | 2.4 ± 0.1 a | 19.4 ± 2.0 ab | 2.4 ± 0.2 ab |
| Wa02 | 2.2 ± 0.8 a | 2.7 ± 1.4 a | 19.3 ± 2.6 ab | 2.4 ± 0.3 ab |
| Wa03 | 2. 0 ± 0.4 a | 2.4 ± 0.7 a | 17.4 ± 3.0 ab | 2.2 ± 0.4 ab |
| Wa04 | 2.3 ± 0.3 a | 1.5 ± 1.0 a | 21.1 ± 5.8 ab | 2.6 ± 0.7 ab |
| Wa05 | 1.8 ± 0.1 a | 1.5 ± 0.8 a | 14.6 ± 1.5 ab | 1.8 ± 0.2 ab |
| Wa06 | 2.4 ± 0.2 a | 1.7 ± 0.6 a | 18.3 ± 1.1 ab | 2.3 ± 0.1 ab |
| Wa07 | 2.7 ± 0.1 a | 2.2 ± 0.4 a | 17.7 ± 3.7 ab | 2.2 ± 0.5 ab |
| Wa08 | 2.6 ± 0.2 a | 1.3 ± 1.0 a | 24.7 ± 3.6 b | 3.1 ± 0.5 b |
| Wa09 | 2.1 ± 0.9 a | 2.0 ± 0.7 a | 14.6 ± 1.8 ab | 1.8 ± 0.2 ab |
| Wa10 | 2.3 ± 0.7 a | 0.4 ± 0.2 a | 24.7 ± 4.7 b | 3.1 ± 0.6 b |
| Wa11 | 2.0 ± 0.6 a | 1.8 ± 0.4 a | 17.5 ± 4.8 ab | 2.2 ± 0.6 ab |
| Wa12 | 2.2 ± 0.7 a | 1.2 ± 1.7 a | 22.2 ± 7.0 ab | 2.8 ± 0.9 ab |
| Wa13 | 2.0 ± 0.6 a | 0.6 ± 0.8 a | 21.6 ± 8.0 ab | 2.7 ± 1.0 ab |
| Wa14 | 1.8 ± 0.1 a | 1.1 ± 1.0 a | 16.3 ± 3.8 ab | 2.0 ± 0.5 ab |
| Wa15 | 1.5 ± 0.5 a | 1.5 ± 1.0 a | 11.2 ± 2.4 a | 1.4 ± 0.3 a |
| Wa16 | 2.0 ± 0.1 a | 1.5 ± 0.8 a | 13.1 ± 1.4 ab | 1.6 ± 0.2 ab |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Enríquez, L.; Vila-Crespo, J.; Rodríguez-Nogales, J.M.; Fernández-Fernández, E.; Ruipérez, V. Non-Saccharomyces Yeasts from Organic Vineyards as Spontaneous Fermentation Agents. Foods 2023, 12, 3644. https://doi.org/10.3390/foods12193644
López-Enríquez L, Vila-Crespo J, Rodríguez-Nogales JM, Fernández-Fernández E, Ruipérez V. Non-Saccharomyces Yeasts from Organic Vineyards as Spontaneous Fermentation Agents. Foods. 2023; 12(19):3644. https://doi.org/10.3390/foods12193644
Chicago/Turabian StyleLópez-Enríquez, Lorena, Josefina Vila-Crespo, José Manuel Rodríguez-Nogales, Encarnación Fernández-Fernández, and Violeta Ruipérez. 2023. "Non-Saccharomyces Yeasts from Organic Vineyards as Spontaneous Fermentation Agents" Foods 12, no. 19: 3644. https://doi.org/10.3390/foods12193644
APA StyleLópez-Enríquez, L., Vila-Crespo, J., Rodríguez-Nogales, J. M., Fernández-Fernández, E., & Ruipérez, V. (2023). Non-Saccharomyces Yeasts from Organic Vineyards as Spontaneous Fermentation Agents. Foods, 12(19), 3644. https://doi.org/10.3390/foods12193644