
Antitumor Effects of Poplar Propolis on DLBCL SU-DHL-2 Cells

Abstract
:1. Introduction
2. Materials and Methods
2.1. Propolis Extraction and Components Determination
2.2. Determination of IC50 of Propolis Extract on SU-DHL-2 Cell
2.3. Different Proteins of Different Group Cells by Proteomics
2.4. Detection of Relative Gene Expression
2.5. ROS Staining
2.6. Statistical Analysis
3. Results
3.1. Chemical Composition of Ethanol Extracts of Propolis
3.2. The Inhibition Effects of EEP on SU-DHL-2 Cell
3.3. Differential Proteins of SU-DHL-2 Cells Treated by EEP and Control
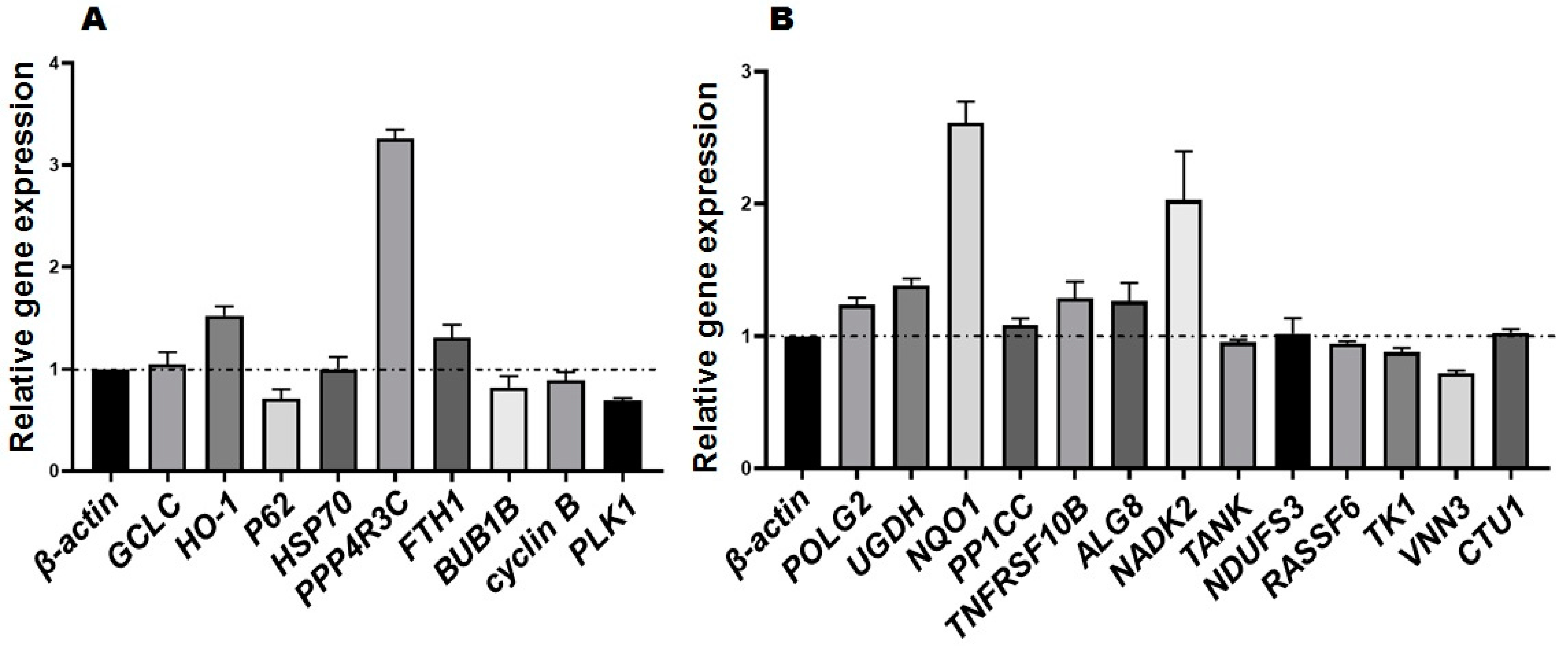
3.4. RT-PCR Identification
3.5. ROS Staining of SU-DHL-2 Cell
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kasote, D.; Bankova, V.; Viljoen, A.M. Propolis: Chemical diversity and challenges in quality control. Phytochem. Rev. 2022, 21, 1887–1911. [Google Scholar] [CrossRef] [PubMed]
- Salatino, A.; Salatino, M.L.F. Scientific note: Often quoted, but not factual data about propolis composition. Apidologie 2021, 52, 312–314. [Google Scholar] [CrossRef]
- Šturm, L.; Ulrih, N.P. Advances in the propolis chemical composition between 2013 and 2018: A review. eFood 2020, 1, 24–37. [Google Scholar] [CrossRef] [Green Version]
- El-Guendouz, S.; Lyoussi, B.; Miguel, M.G. Insight on propolis from mediterranean countries: Chemical composition, biological activities and application fields. Chem. Biodivers 2019, 16, e1900094. [Google Scholar] [CrossRef] [PubMed]
- Braga, V.N.L.; Juanes, C.D.C.; Peres, H.D.S.; Sousa, J.R.D.; Cavalcanti, B.C.; Jamacaru, F.V.F.; de Lemos, T.L.G.; Dornelas, C.A. Gum arabic and red propolis protecteting colorectal preneoplastic lesions in a rat model of azoxymethane1. Acta Cir. Bras. 2019, 34, e201900207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sameni, H.R.; Yosefi, S.; Alipour, M.; Pakdel, A.; Torabizadeh, N.; Semnani, V.; Bandegi, A.R. Co-administration of 5FU and propolis on AOM/DSS induced colorectal cancer in BALB-c mice. Life Sci. 2021, 276, 119390. [Google Scholar] [CrossRef]
- Demir, S.; Aliyazicioglu, Y.; Turan, I.; Misir, S.; Mentese, A.; Yaman, S.O.; Akbulut, K.; Deger, O. Antiproliferative and proapoptotic activity of Turkish propolis on human lung cancer cell line. Nutr. Cancer 2016, 68, 165–172. [Google Scholar] [CrossRef]
- Xuan, H.; Li, Z.; Yan, H.; Sang, Q.; Wang, K.; He, Q.; Wang, Y.; Hu, F. Antitumor activity of Chinese propolis in human breast cancer MCF-7 and MDA-MB-231 Cells. Evid-Based Compl. Alt. 2014, 2014, e280120. [Google Scholar] [CrossRef] [Green Version]
- Prasad, S.B.; Turnia, I. Antitumor activity with no toxicity of propolis from Meghalaya, India in ascites Dalton’s lymphoma-bearing mice. Indian J. Nat. Prod. Resour. 2021, 11, 267–279. [Google Scholar] [CrossRef]
- Alday, E.; Valencia, D.; Carreño, A.; Picerno, P.; Piccinelli, A.L.; Rastrelli, L.; Robles-Zepeda, R.; Hernandez, J.; Velazquez, C. Apoptotic induction by pinobanksin and some of its ester derivatives from Sonoran propolis in a B-cell lymphoma cell line. Chem.-Biol. Interact. 2015, 242, 35–44. [Google Scholar] [CrossRef]
- Benguedouar, L.; Lahouel, M.; Gangloff, S.; Durlach, A.; Grange, F.; Bernard, P.; Antonicelli, F. Algerian ethanolic extract of propolis and galangin decreased melanoma tumour progression in C57BL6 mice. Ann. Dermatol. Syphiligr. 2015, 142, S294. [Google Scholar] [CrossRef]
- Kimoto, T.; Arai, S.; Kohguchi, M.; Aga, M.; Nomura, Y.; Micallef, M.J.; Kurimoto, M.; Mito, K. Apoptosis and suppression of tumor growth by artepillin C extracted from Brazilian propolis. Cancer Detect. Prev. 1998, 22, 506–515. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, K.; Ashida, H.; Matsuura, Y.; Kanazawa, K. Antioxidative bioavailability of artepillin C in Brazilian propolis. Arch. Biochem. Biophys. 2004, 424, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Messerli, S.M.; Ahn, M.R.; Kunimasa, K.; Yanagihara, M.; Tatefuji, T.; Hashimoto, K.; Mautner, V.; Uto, Y.; Hori, H.; Kumazawa, S.; et al. Artepillin C (ARC) in Brazilian green propolis selectively blocks oncogenic PAK1 signaling and suppresses the growth of NF tumors in mice. Phytother. Res. 2009, 23, 423–427. [Google Scholar] [CrossRef]
- Grunberger, D.; Banerjee, R.; Eisinger, K.; Oltz, E.M.; Efros, L.; Caldwell, M.; Estevez, V.; Nakanishi, K. Preferential cytotoxicity on tumor cells by caffeic acid phenethyl ester isolated from propolis. Experientia 1988, 44, 230–232. [Google Scholar] [CrossRef]
- Jayakumar, A.; Venugopal, R.; Zielinski, R.; Rusin, A.; Fokt, I.; Skora, S.; Ni, X.; Duvis, M.; Priebe, W. High sensitivity of cutaneous T-cell lymphoma (CTCL) to CABE, a component of propolis. Cancer Res. 2015, 75, 2096. [Google Scholar] [CrossRef]
- Arber, D.A.; Orazi, A.; Hasserjian, R.; Thiele, J.; Borowitz, M.J.; Le Beau, M.M.; Bloomfield, C.D.; Cazzola, M.; Vardiman, J.W. The 2016 revision of the World Health Organization classification of lymphoid neoplasms. Blood 2016, 127, 2391–2405. [Google Scholar] [CrossRef]
- Li, S.; Young, K.H.; Medeiros, L.J. Diffuse large B-cell lymphoma. Pathology 2018, 50, 74–87. [Google Scholar] [CrossRef] [Green Version]
- Yao, S.; Xu, F.; Chen, Y.; Ge, Y.; Zhang, F.; Huang, H.; Li, L.; Lin, D.; Luo, X.; Xu, J.; et al. Fbw7 regulates apoptosis in activated B-cell like diffuse large B-cell lymphoma by targeting Stat3 for ubiquitylation and degradation. J. Exp. Clin. Cancer Res. 2017, 36, e10. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Sun, Y.; Wang, J.; He, Q.; Chen, X.; Lan, X.; Chen, J.; Dou, Q.; Shi, X.; Liu, J. Proteasomal cysteine deubiquitinase inhibitor b-AP15 suppresses migration and induces apoptosis in diffuse large B cell lymphoma. J. Exp. Clin. Cancer Res. 2019, 38, 1756–1766. [Google Scholar] [CrossRef]
- Shi, L.; Zhu, H.; Zhang, M.; Liu, Y.; Wang, Y.; Zhao, J.; Lei, F.; He, P. Effect of realgar on induction of apoptosis in DLBCL cell line SU-DHL-4 and its possible mechanisms. J. Exp. Hematol. 2014, 22, 729–734. [Google Scholar] [CrossRef]
- Li, J.; Zhang, J.; Wang, X.; Sun, Z. Effects of 17-DMAG on diffuse large B-cell lymphoma cell apoptosis. Exp. Ther. Med. 2017, 14, 3727–3731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, B.; Yan, F.; Wu, C. Overexpressed miR-195 attenuated immune escape of diffuse large B-cell lymphoma by targeting PD-L1. Biomed. Pharmacother. 2018, 98, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Pu, R.; Li, Y.; Wu, Z.; Li, C.; Miao, X.; Yang, W. Chemical compositions of propolis from China and the United States and their antimicrobial activities against Penicillium notatum. Molecules 2019, 24, 3576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kachuk, C.; Stephen, K.; Doucette, A. Comparison of sodium dodecyl sulfate depletion techniques for proteome analysis by mass spectrometry. J. Chromatogr. A 2015, 1418, 158–166. [Google Scholar] [CrossRef]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef]
- Noaman, N.; Coorssen, J.R. Coomassie does it (better): A Robin Hood approach to total protein quantification. Anal. Biochem. 2018, 556, 53–56. [Google Scholar] [CrossRef]
- Liu, M.; Jin, L.; Sun, S.; Liu, P.; Feng, X.; Cheng, Z.; Liu, W.; Guan, K.; Shi, Y.; Yuan, H. Metabolic reprogramming by PCK1 promotes TCA cataplerosis, oxidative stress and apoptosis in liver cancer cells and suppresses hepatocellular carcinoma. Oncogene 2018, 37, 1637–1653. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Sun, Y.; Rao, G. Studies on chemical constituents of Yunnan propolis. J. Chin. Med. Mater. 2016, 39, 2247–2250. [Google Scholar] [CrossRef]
- Yao, J.; Sun, J.; Wu, L.; Wang, H.; Wang, G.; Ye, W. Identification of major constituents from propolis by LC-IT-TOF-MS. J. China Pharm. Univ. 2017, 48, 178–183. [Google Scholar] [CrossRef]
- de Groot, A.C. Propolis: A review of properties, applications, chemical composition, contact allergy, and other adverse effects. Dermatitis 2013, 24, 263–282. [Google Scholar] [CrossRef] [PubMed]
- Wezgowiec, J.; Wieczynska, A.; Wieckiewicz, W.; Kulbacka, J.; Saczko, J.; Pachura, N.; Wieckiewicz, M.; Gancarz, R.; Wilk, K.A. Polish propolis-chemical composition and biological effects in tongue cancer cells and macrophages. Molecules 2020, 25, 2426. [Google Scholar] [CrossRef] [PubMed]
- Ilhan-Ayisigi, E.; Ulucan, F.; Saygili, E.; Saglam-Metiner, P.; Gulce-Iz, S.; Yesil-Celiktas, O. Nano-vesicular formulation of propolis and cytotoxic effects in a 3D spheroid model of lung cancer. J. Sci. Food Agric. 2020, 100, 3525–3535. [Google Scholar] [CrossRef] [PubMed]
- Khacha-Ananda, S.; Tragoolpua, K.; Chantawannakul, P.; Tragoolpua, Y. Propolis extracts from the northern region of Thailand suppress cancer cell growth through induction of apoptosis pathways. Investig. New Drugs 2016, 34, 707–722. [Google Scholar] [CrossRef]
- da Silva Frozza, C.O.; Santos, D.A.; Rufatto, L.C.; Minetto, L.; Scariot, F.J.; Echeverrigaray, S.; Pich, C.T.; Moura, S.; Padilha, F.F.; Borsuk, S. Antitumor activity of Brazilian red propolis fractions against Hep-2 cancer cell line. Biomed. Pharmacother. 2017, 91, 951–963. [Google Scholar] [CrossRef]
- Xuan, H.; Wang, Y.; Li, A.; Fu, C.; Wang, Y.; Peng, W. Bioactive components of Chinese propolis water extract on antitumor activity and quality control. Evid-Based Compl. Alt. 2016, 2016, 9641965. [Google Scholar] [CrossRef] [Green Version]
- Avcı, Ç.B.; Gündüz, C.; Baran, Y.; Şahin, F.; Yılmaz, S.; Dogan, Z.O.; Saydam, G. Caffeic acid phenethyl ester triggers apoptosis through induction of loss of mitochondrial membrane potential in CCRF-CEM cells. J. Cancer Res. Clin. Oncol. 2011, 137, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Xu, T.; Ding, W.; Ji, X.; Ao, X.; Liu, Y.; Yu, W.; Wang, J. Molecular mechanisms of ferroptosis and its role in cancer therapy. J. Cell. Mol. Med. 2019, 23, 4900–4912. [Google Scholar] [CrossRef] [Green Version]
- Cortes-Wanstreet, M.M.; Giedzinski, E.; Limoli, C.L.; Luderer, U. Overexpression of glutamate–cysteine ligase protects human COV434 granulosa tumour cells against oxidative and γ-radiation-induced cell death. Mutagenesis 2009, 24, 211–224. [Google Scholar] [CrossRef]
- Bracht, K.; Grünert, R.; Bednarski, P.J. Correlations between the activities of 19 anti-tumor agents and the intracellular glutathione concentrations in a panel of 14 human cancer cell lines: Comparisons with the National Cancer Institute data. Anti-Cancer Drugs 2006, 17, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Biamonte, F.; Battaglia, A.M.; Zolea, F.; Oliveira, D.M.; Aversa, I.; Santamaria, G.; Giovannone, E.D.; Rocco, G.; Viglietto, G.; Costanzo, F. Ferritin heavy subunit enhances apoptosis of non-small cell lung cancer cells through modulation of miR-125b/p53 axis. Cell Death Dis. 2018, 9, 1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, A.; Shafarin, J.; Abu Jabal, R.; Aljabi, N.; Hamad, M.; Sualeh Muhammad, J.; Unnikannan, H.; Hamad, M. Ferritin heavy chain (FTH1) exerts significant antigrowth effects in breast cancer cells by inhibiting the expression of c-MYC. FEBS Open Bio. 2021, 11, 3101–3114. [Google Scholar] [CrossRef] [PubMed]
- Lobello, N.; Biamonte, F.; Pisanu, M.E.; Faniello, M.C.; Jakopin, Ž.; Chiarella, E.; Giovannone, E.D.; Mancini, R.; Ciliberto, G.; Cuda, G.; et al. Ferritin heavy chain is a negative regulator of ovarian cancer stem cell expansion and epithelial to mesenchymal transition. Oncotarget 2016, 7, 62019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Podkalicka, P.; Mucha, O.; Józkowicz, A.; Dulak, J.; Łoboda, A. Heme oxygenase inhibition in cancers: Possible tools and targets. Contemp. Oncol. (Pozn) 2018, 22, 23–32. [Google Scholar] [CrossRef]
- Kwon, M.Y.; Park, E.; Lee, S.J.; Chung, S.W. Heme oxygenase-1 accelerates erastin-induced ferroptotic cell death. Oncotarget 2015, 6, 24393–24403. [Google Scholar] [CrossRef] [Green Version]
- Chang, L.; Chiang, S.; Chen, S.; Yu, Y.; Chou, R.; Chang, W. Heme oxygenase-1 mediates BAY 11–7085 induced ferroptosis. Cancer Lett. 2018, 416, 124–137. [Google Scholar] [CrossRef]
- Malfa, G.A.; Tomasello, B.; Acquaviva, R.; Genovese, C.; La Mantia, A.; Cammarata, F.P.; Ragusa, M.; Renis, M.; Di Giacomo, C. Betula etnensis Raf. (Betulaceae) extract induced HO-1 expression and ferroptosis cell death in human colon cancer cells. Int. J. Mol. Sci. 2019, 20, 2723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fest, S.; Soldati, R.; Christiansen, N.M.; Zenclussen, M.L.; Kilz, J.; Berger, E.; Starke, S.; Lode, H.N.; Engel, C.; Zenclussen, A.C.; et al. Targeting of heme oxygenase-1 as a novel immune regulator of neuroblastoma. Int. J. Cancer 2016, 138, 2030–2042. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.; Chen, F.; Jiang, J.; Zhang, H.; Zhou, M. Screening key genes and signaling pathways in colorectal cancer by integrated bioinformatics analysis. Mol. Med. Rep. 2019, 20, 1259–1269. [Google Scholar] [CrossRef]
- Zeng, H.; Lazarova, D.L.; Bordonaro, M. Mechanisms linking dietary fiber, gut microbiota and colon cancer prevention. World J. Gastrointest Oncol. 2014, 6, 41. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Duan, X.; Zeng, G. Identification of potential biomarkers and pivotal biological pathways for prostate cancer using bioinformatics analysis methods. PeerJ 2019, 7, e7872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thai, S.F.; Jones, C.P.; Robinette, B.L.; Ren, H.; Vallant, B.; Fisher, A.; Kitchin, K.T. Effects of copper nanoparticles on mRNA and small RNA expression in human hepatocellular carcinoma (HepG2) cells. J. Nanosci. Nanotechnol. 2021, 21, 5083–5098. [Google Scholar] [CrossRef] [PubMed]
- Denk, H.; Stumptner, C.; Abuja, P.M.; Zatloukal, K. Sequestosome 1/p62-related pathways as therapeutic targets in hepatocellular carcinoma. Expert Opin. Ther. Targets 2019, 23, 393–406. [Google Scholar] [CrossRef] [PubMed]
- Katsuragi, Y.; Ichimura, Y.; Komatsu, M. Regulation of the Keap1–Nrf2 pathway by p62/SQSTM1. Curr. Opin. Toxicol. 2016, 1, 54–61. [Google Scholar] [CrossRef] [Green Version]
- Patiño-Morales, C.C.; Soto-Reyes, E.; Arechaga-Ocampo, E.; Ortiz-Sánchez, E.; Antonio-Véjar, V.; Pedraza-Chaverri, J.; García-Carrancá, A. Curcumin stabilizes p53 by interaction with NAD (P) H: Quinone oxidoreductase 1 in tumor-derived cell lines. Redox Biol. 2020, 28, 101320. [Google Scholar] [CrossRef]
- Sheikhshabani, S.H.; Amini-Farsani, Z.; Rahmati, S.; Jazaeri, A.; Mohammadi-Samani, M.; Asgharzade, S. Oleuropein reduces cisplatin resistance in ovarian cancer by targeting apoptotic pathway regulators. Life Sci. 2021, 278, 119525. [Google Scholar] [CrossRef]
- Pitts, T.M.; Davis, S.L.; Eckhardt, S.G.; Bradshaw-Pierce, E.L. Targeting nuclear kinases in cancer: Development of cell cycle kinase inhibitors. Pharmacol. Ther. 2014, 142, 258–269. [Google Scholar] [CrossRef]
- Blagden, S.; Bono, J. Drugging cell cycle kinases in cancer therapy. Curr. Drug Targets 2005, 6, 325–335. [Google Scholar] [CrossRef]
- Mohajan, S.; Jaiswal, P.K.; Vatanmakarian, M.; Yousefi, H.; Sankaralingam, S.; Alahari, S.K.; Alahari, S.K.; Koul, S.; Koul, H.K. Hippo pathway: Regulation, deregulation and potential therapeutic targets in cancer. Cancer Lett. 2021, 507, 112–123. [Google Scholar] [CrossRef]
- Kang, G.Y.; Lee, E.R.; Kim, J.H.; Jung, J.W.; Lim, J.; Kim, S.K.; CHO, S.G.; Kim, K.P. Downregulation of PLK-1 expression in kaempferol-induced apoptosis of MCF-7 cells. Eur. J. Pharmacol. 2009, 611, 17–21. [Google Scholar] [CrossRef]
- Dimri, M.; Cho, J.H.; Kang, M.; Dimri, G.P. PLK1 inhibition down-regulates polycomb group protein BMI1 via modulation of the miR-200c/141 cluster. J. Biol. Chem. 2015, 290, 3033–3044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spänkuch, B.; Heim, S.; Kurunci-Csacsko, E.; Lindenau, C.; Yuan, J.; Kaufmann, M.; Strebhardt, K. Down-regulation of Polo-like kinase 1 elevates drug sensitivity of breast cancer cells in vitro and in vivo. Cancer Res. 2006, 66, 5836–5846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, C.P.; Gomes-da-Silva, L.C.; Ramalho, J.S.; de Lima, M.C.; Simoes, S.; Moreira, J.N. Impact of PLK-1 silencing on endothelial cells and cancer cells of diverse histological origin. Curr. Gene Ther. 2013, 13, 189–201. [Google Scholar] [CrossRef]
- Nihal, M.; Stutz, N.; Schmit, T.; Ahmad, N.; Wood, G.S. Polo-like kinase 1 (Plk1) is expressed by cutaneous T-cell lymphomas (CTCLs), and its downregulation promotes cell cycle arrest and apoptosis. Cell Cycle 2011, 10, 1303–1311. [Google Scholar] [CrossRef]
- Shao, C.; Ahmad, N.; Hodges, K.; Kuang, S.; Ratliff, T.; Liu, X. Inhibition of polo-like kinase 1 (Plk1) enhances the antineoplastic activity of metformin in prostate cancer. J. Biol. Chem. 2015, 290, 2024–2033. [Google Scholar] [CrossRef] [Green Version]
- Spänkuch-Schmitt, B.; Bereiter-Hahn, J.; Kaufmann, M.; Strebhardt, K. Effect of RNA silencing of polo-like kinase-1 (PLK1) on apoptosis and spindle formation in human cancer cells. J. Natl. Cancer Inst. 2002, 94, 1863–1877. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Erikson, R.L. Polo-like kinase (Plk) 1 depletion induces apoptosis in cancer cells. Proc. Natl. Acad. Sci. USA 2003, 100, 5789–5794. [Google Scholar] [CrossRef] [Green Version]
- Raab, C.A.; Raab, M.; Becker, S.; Strebhardt, K. Non-mitotic functions of polo-like kinases in cancer cells. Biochim. Biophys. Acta Rev. Cancer 2021, 1875, 188467. [Google Scholar] [CrossRef]
- Kato, T.; Wada, H.; Patel, P.; Hu, H.P.; Lee, D.; Ujiie, H.; Hirohashi, K.; Nakajima, T.; Sato, M.; Kaji, M.; et al. Overexpression of KIF23 predicts clinical outcome in primary lung cancer patients. Lung Cancer 2016, 92, 53–61. [Google Scholar] [CrossRef]
- Iltzsche, F.; Simon, K.; Stopp, S.; Pattschull, G.; Francke, S.; Wolter, P.; Hauser, S.; Murphy, D.J.; Garcia, P.; Rosenwald, A.; et al. An important role for Myb-MuvB and its target gene KIF23 in a mouse model of lung adenocarcinoma. Oncogene 2017, 36, 110–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jian, W.; Deng, X.; Munankarmy, A.; Borkhuu, O.; Ji, C.; Wang, X.; Zheng, W.; Yu, Y.; Zhou, X.; Fang, L. KIF23 promotes triple negative breast cancer through activating epithelial-mesenchymal transition. Gland. Surg. 2021, 10, 1941. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Fusaki, N.; Ohta, S.; Iwahori, Y.; Iizuka, Y.; Inagawa, K.; Kawakami, Y.; Yoshida, K.; Toda, M. Downregulation of KIF23 suppresses glioma proliferation. J. Neuro-Oncol. 2012, 106, 519–529. [Google Scholar] [CrossRef]
- Liang, W.; Liu, X.; Huang, H.; Gao, Z.; Li, K. Prognostic significance of KIF23 expression in gastric cancer. World J. Gastro Oncol. 2020, 12, 1104. [Google Scholar] [CrossRef] [PubMed]
- Vostakolaei, M.A.; Hatami-Baroogh, L.; Babaei, G.; Molavi, O.; Kordi, S.; Abdolalizadeh, J. Hsp70 in cancer: A double agent in the battle between survival and death. J. Cell. Physiol. 2021, 236, 3420–3444. [Google Scholar] [CrossRef]
- Gonçalves, E.; Fragoulis, A.; Garcia-Alonso, L.; Cramer, T.; Saez-Rodriguez, J.; Beltrao, P. Widespread post-transcriptional attenuation of genomic copy-number variation in cancer. Cell Syst. 2017, 5, 386–398. [Google Scholar] [CrossRef] [Green Version]
- Akbani, R.; Ng, P.K.S.; Werner, H.M.; Shahmoradgoli, M.; Zhang, F.; Ju, Z.; Liu, W.; Yang, J.; Yoshihara, K.; Li, J.; et al. A pan-cancer proteomic perspective on The Cancer Genome Atlas. Nat. Commun. 2014, 5, 3887. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Liu, T.; Zhang, Z.; Payne, S.H.; Zhang, B.; McDermott, J.E.; Zhou, J.; Petyuk, V.A.; Chen, L.; Ray, D.; et al. Integrated proteogenomic characterization of human high-grade serous ovarian cancer. Cell 2016, 166, 755–765. [Google Scholar] [CrossRef] [Green Version]
- Johansson, H.J.; Socciarelli, F.; Vacanti, N.M.; Haugen, M.H.; Zhu, Y.; Siavelis, I.; Fernandez-Woodbridge, A.; Aure, M.R.; Sennblad, B.; Vesterlund, M.; et al. Breast cancer quantitative proteome and proteogenomic landscape. Nat. Commun. 2019, 10, 1600. [Google Scholar] [CrossRef]
- Falcão, S.I.; Vale, N.; Gomes, P.; Domingues, M.R.; Freire, C.; Cardoso, S.M.; Vilas-Boas, M. Phenolic profiling of Portuguese propolis by LC–MS spectrometry: Uncommon propolis rich in flavonoid glycosides. Phytochem. Anal. 2013, 24, 309–318. [Google Scholar]
- Nagaoka, T.; Banskota, A.H.; Tezuka, Y.; Midorikawa, K.; Matsushige, K.; Kadota, S. Caffeic acid phenethyl ester (CAPE) analogues: Potent nitric oxide inhibitors from the Netherlands propolis. Biol. Pharm. Bull. 2003, 26, 487–491. [Google Scholar]
- Bilikova, K.; Popova, M.; Trusheva, B.; Bankova, V.A. New anti-Paenibacillus larvae substances purified from propolis. Apidologie 2013, 44, 278–285. [Google Scholar]
- Bloor, S.; Catchpole, O.; Mitchell, K.; Webby, R.; Davis, P. Antiproliferative acylated glycerols from New Zealand propolis. J. Nat. Prod. 2019, 82, 2359–2367. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathway | p-Value | Description of Proteins |
|---|---|---|
| Ferroptosis | 0.0016 | Glutamate–cysteine ligase regulatory subunit, Ferritin, Heme oxygenase |
| Mineral absorption | 0.0076 | Ferritin, Heme oxygenase |
| Fluid shear stress and atherosclerosis | 0.0108 | Sequestosome-1, NAD(P)H dehydrogenase [quinone] 1, Heme oxygenase |
| Ubiquinone and other terpenoid–quinone biosynthesis | 0.0247 | NAD(P)H dehydrogenase [quinone] 1 |
| Necroptosis | 0.0321 | Sequestosome-1, Ferritin, Tumor necrosis factor receptor superfamily member 10A |
| Influenza A | 0.0414 | DnaJ homolog subfamily C member 3, Epididymis secretory sperm binding protein, Tumor necrosis factor receptor superfamily member 10A |
| Ubiquitin-mediated proteolysis | 0.0431 | NEDD8-conjugating enzyme UBE2F, Ubiquitin-conjugating enzyme E2 J1, cDNA, FLJ92255, highly similar to Homo sapiens ring finger protein 7 (RNF7), mRNA |
| Ascorbate and aldarate metabolism | 0.0488 | UDP-glucose 6-dehydrogenase |
| Antigen processing and presentation | 0.0491 | Epididymis secretory sperm binding protein, cDNA FLJ78235 |
| Cell cycle | 0.0201 | Mitotic checkpoint serine/threonine-protein kinase BUB1 beta, Serine/threonine-protein kinase PLK, G2/mitotic-specific cyclin-B2 |
| Progesterone-mediated oocyte maturation | 0.0387 | Serine/threonine-protein kinase PLK,G2/mitotic-specific cyclin-B2 |
| FoxO signaling pathway | 0.0430 | Serine/threonine-protein kinase PLK,G2/mitotic-specific cyclin-B2 |
| Hippo signaling pathway—multiple species | 0.0457 | Ras association domain-containing protein 6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Tian, Y.; Yang, A.; Zhang, C.; Miao, X.; Yang, W. Antitumor Effects of Poplar Propolis on DLBCL SU-DHL-2 Cells. Foods 2023, 12, 283. https://doi.org/10.3390/foods12020283
Liu X, Tian Y, Yang A, Zhang C, Miao X, Yang W. Antitumor Effects of Poplar Propolis on DLBCL SU-DHL-2 Cells. Foods. 2023; 12(2):283. https://doi.org/10.3390/foods12020283
Chicago/Turabian StyleLiu, Xiaoqing, Yuanyuan Tian, Ao Yang, Chuang Zhang, Xiaoqing Miao, and Wenchao Yang. 2023. "Antitumor Effects of Poplar Propolis on DLBCL SU-DHL-2 Cells" Foods 12, no. 2: 283. https://doi.org/10.3390/foods12020283
APA StyleLiu, X., Tian, Y., Yang, A., Zhang, C., Miao, X., & Yang, W. (2023). Antitumor Effects of Poplar Propolis on DLBCL SU-DHL-2 Cells. Foods, 12(2), 283. https://doi.org/10.3390/foods12020283