


Comparative Assessment of Functional Components and Antioxidant Activities between Hippophae rhamnoides ssp. sinensis and H. tibetana Berries in Qinghai–Tibet Plateau


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Biochemical Sampling
2.3. Determination of Soluble Sugar Content
2.4. Determination of Organic Acid Content
2.5. Determination of Phenolic and Flavonoid Content
2.6. Determination of VC and VE Content
2.7. Determination of Carotenoid Content
2.8. In Vitro Antioxidant Activity Assays
2.9. Cell Cytotoxicity Assay
2.10. Cellular Antioxidant Activity (CAA) Assay
2.11. Data Statistics
3. Results and Discussion
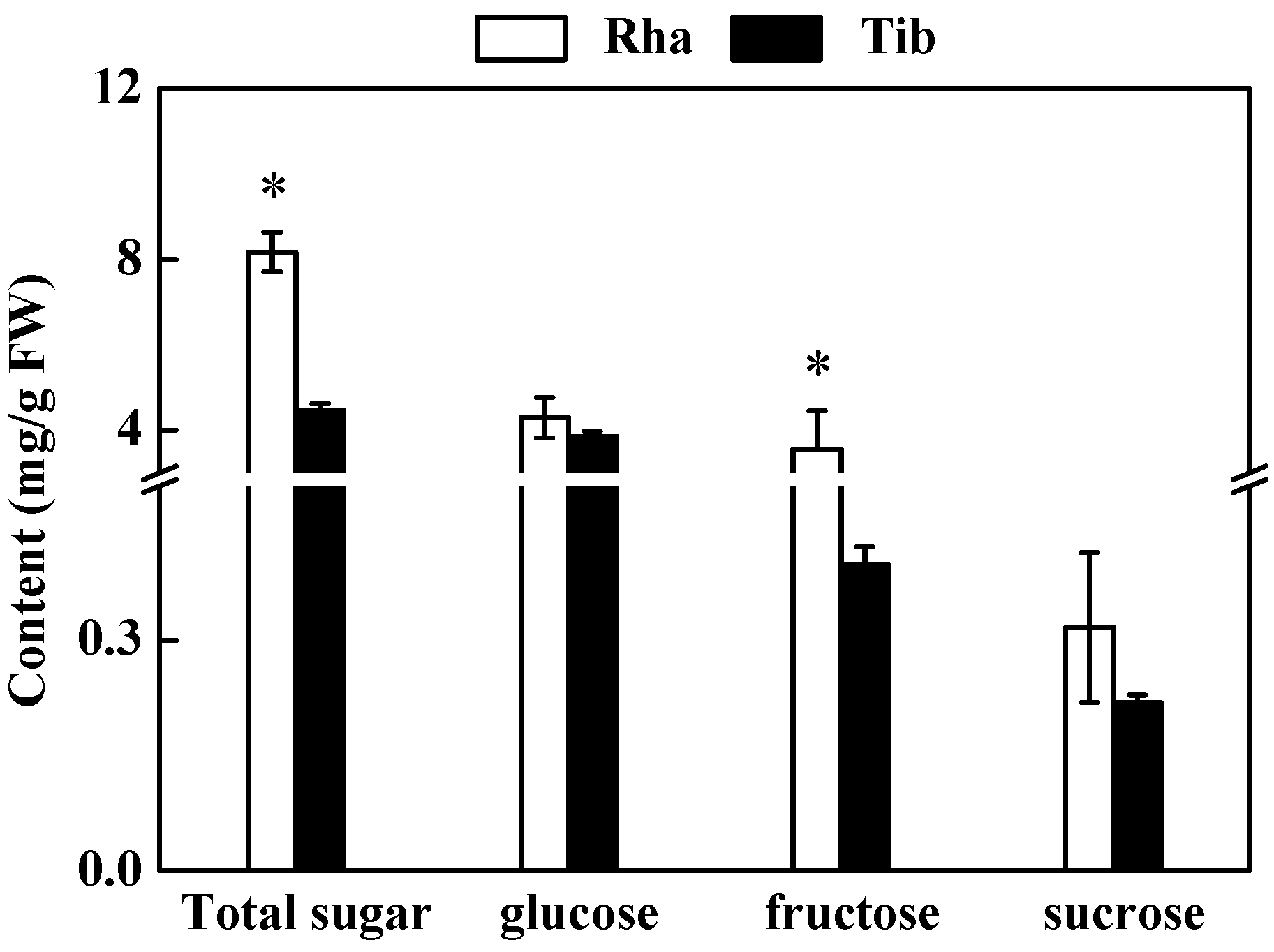
3.1. The Content of Soluble Sugars in Rha and Tib Berries
3.2. The Content of Organic Acids in Rha and Tib Berries
3.3. The Content of Phenolics, Flavonoids and Twelve Phenolics in Rha and Tib Berries
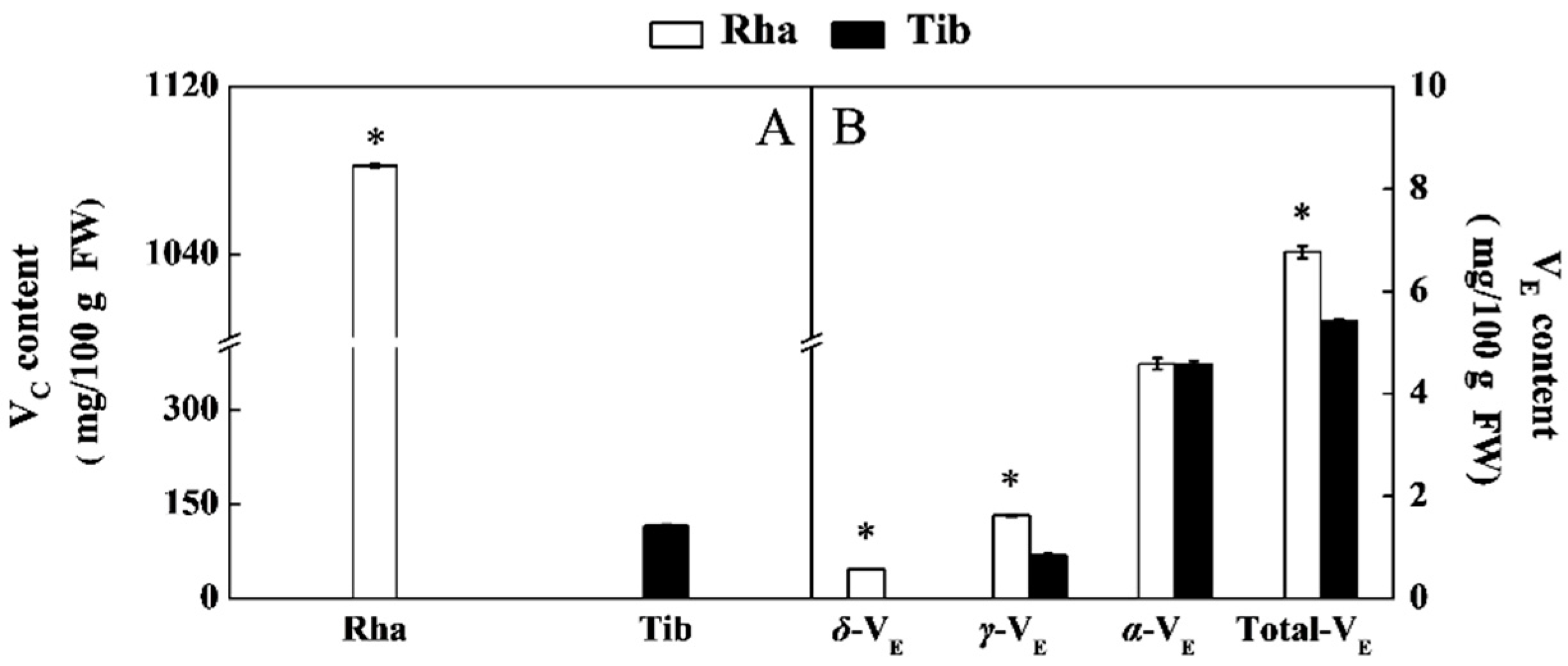
3.4. The Content of VC and VE in Rha and Tib Berries
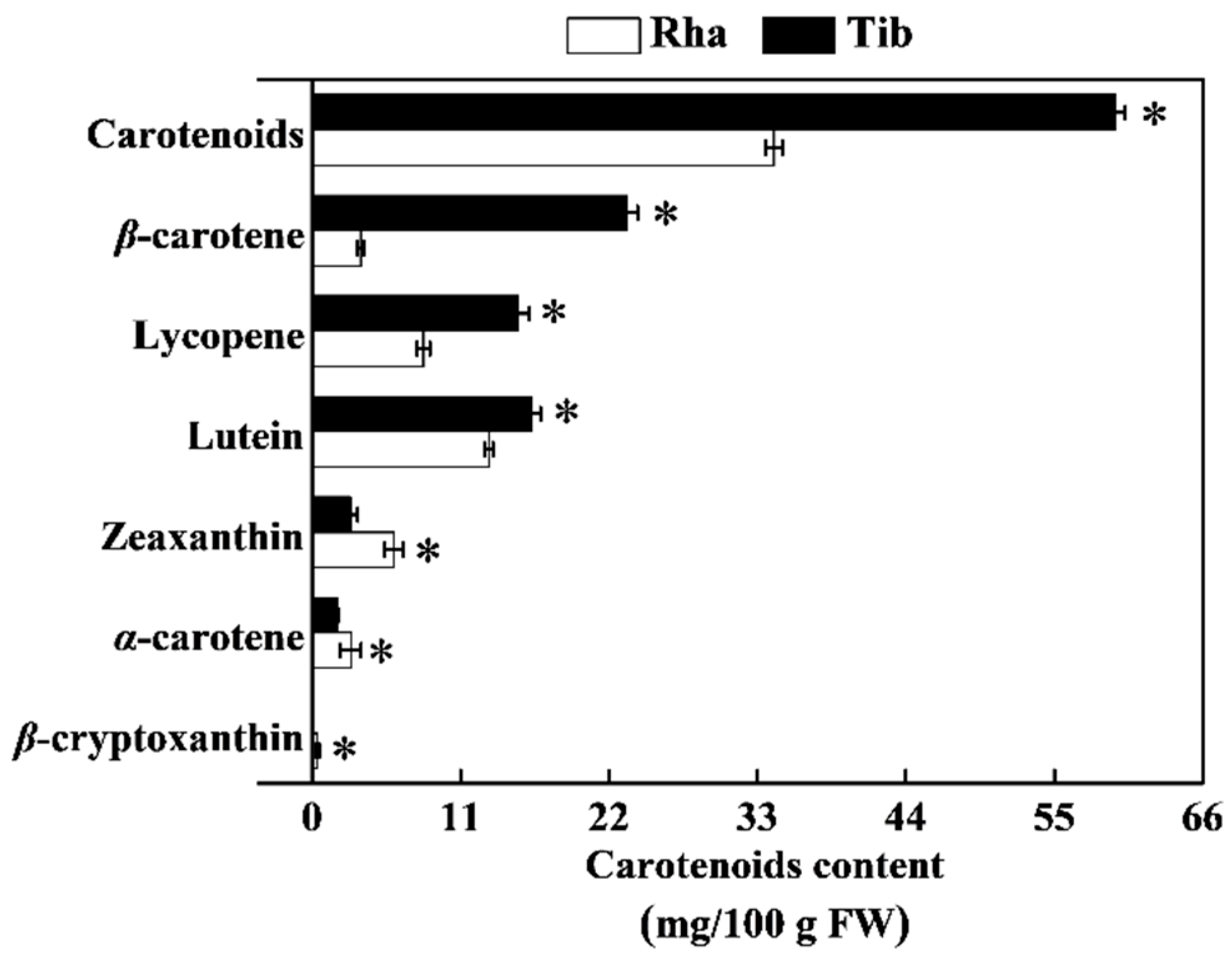
3.5. The Content of Carotenoids in Rha and Tib Berries
3.6. In Vitro Antioxidant Activity of Rha and Tib Berries
3.7. Cell Cytotoxicity and CAA of Rha and Tib Berries
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wang, N.N.; Wen, X.F.; Gao, Y.; Lu, S.G.; Li, Y.M.; Shi, Y.B.; Yang, Z.G. Identification and characterization of the bioactive polyphenols and volatile compounds in sea buckthorn leaves tea together with antioxidant and α-glucosidase inhibitory activities. Front. Nutr. 2022, 9, 890486. [Google Scholar] [CrossRef] [PubMed]
- Stobdan, T.; Korekar, G.; Srivastava, R.B. Nutritional attributes and health application of sea buckthorn (Hippophae rhamnoides L.)—A review. Curr. Nutr. Food Sci. 2013, 9, 151–165. [Google Scholar] [CrossRef]
- Olas, B. Sea buckthorn as a source of important bioactive compounds in cardiovascular diseases. Food Chem. Toxicol. 2016, 197, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.W.; Xu, Z.Z.; Liao, X.J. Bioactive compounds, health benefits and functional food products of sea buckthorn: A review. Crit. Rev. Food Sci. Nutr. 2021, 5, 6761–6782. [Google Scholar] [CrossRef]
- Wu, Z.J.; Ruan, C.J.; Li, H.; Yang, M.L.; Xie, M.M.; Xiao, T.T. Compositions of soluble sugars and organic acids in berries of 12 sea buckthorn cultivars. J. Northwest For. Univ. 2016, 31, 106–112, (In Chinese with English abstract). [Google Scholar]
- Wei, J.; Li, S.; Su, T.T.; Zhao, J.M.; Jiang, Y.M.; Zubarev, Y.A.; Bi, Y. Phenolic compositions and antioxidant activities of Hippophae tibetana and H. rhamnoides ssp. sinensis berries produced in Qinghai-Tibet Plateau. Food Chem. X. 2022, 15, 100397. [Google Scholar] [CrossRef] [PubMed]
- Kallio, H.; Yang, B.; Peippo, P. Effects of different origins and harvesting time on vitamin C, tocopherols, and tocotrienols in sea buckthorn (Hippophaë rhamnoides) berries. J. Agric. Food Chem. 2002, 50, 6136–6142. [Google Scholar] [CrossRef]
- Kallio, H.; Yang, B.; Peippo, P.; Tahvonen, R.; Pan, R. Triacylglycerols, glycerophospholipids, tocopherols, and tocotrienols in berries and seeds of two subspecies (ssp. sinensis and mongolica) of sea buckthorn (Hippophaë rhamnoides). J. Agric. Food Chem. 2002, 50, 3004–3009. [Google Scholar] [CrossRef] [PubMed]
- Raffo, A.; Paoletti, F.; Antonelli, M. Changes in sugar, organic acid, flavonol and carotenoid composition during ripening of berries of three seabuckthorn (Hippophae rhamnoides L.) cultivars. Eur. Food. Res. Technol. 2004, 219, 360–368. [Google Scholar] [CrossRef]
- Dąbrowski, G.; Czaplicki, S.; Szustak, M.; Cichońska, E.; Gendaszewska-Darmach, E.; Konopka, I. Composition of flesh lipids and oleosome yield optimization of selected sea buckthorn (Hippophae rhamnoides L.) cultivars grown in Poland. Food Chem. 2022, 369, 130921. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.X.; Guo, X.B.; Li, T.; Fu, X.; Liu, R.H. Comparative assessment of phytochemical profiles, antioxidant and antiproliferative activities in sea buckthorn (Hippophaë rhamnoides L.) berries. Food Chem. 2017, 221, 997–1003. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.Y.; Laaksonen, O.; Zheng, J.; Yang, W.; Trépanier, M.; Kallio, H.; Yang, B. Flavonol glycosides in berries of two major subspecies of sea buckthorn (Hippophaë rhamnoides L.) and influence of growth sites. Food Chem. 2016, 200, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Yang, B.; Trépanier, M.; Kallio, H. Effects of genotype, latitude, and weather conditions on the composition of sugars, sugar alcohols, fruit acids, and ascorbic acid in sea buckthorn (Hippophaë rhamnoides ssp. mongolica) berry juice. J. Agric. Food Chem. 2012, 60, 3180–3189. [Google Scholar] [CrossRef]
- Petreikov, M.; Yeselson, L.; Shen, S.; Levin, I.; Schaffer, A.A.; Efrati, A.; Bar, M. Carbohydrate balance and accumulation during development of near-isogenic tomato lines differing in the AGPase-L1 allele. J. Am. Soc. Hortic. Sci. 2009, 134, 1070–1076. [Google Scholar] [CrossRef] [Green Version]
- Coelho, E.M.; Miskinis, C.V.S.P.; de Sá, A.G.B.; Pereira, G.E.; Azevêdoa, L.C.; Limaa, M.S. Simultaneous analysis of sugars and organic acids in wine and grape juices by HPLC: Method validation and characterization of products from northeast Brazil. J. Food Compos. Anal. 2017, 66, 160–167. [Google Scholar] [CrossRef] [Green Version]
- Sytařová, I.; Orsavová, J.; Snopek, L.; Mlček, J.; Byczyński, Ł.; Ladislava, M. Impact of phenolic compounds and vitamins C and E on antioxidant activity of sea buckthorn (Hippophaë rhamnoides L.) berries and leaves of diverse ripening times. Food Chem. 2019, 310, 125784. [Google Scholar] [CrossRef]
- Tkacz, K.; Wojdyo, A.; Turkiewicz, I.P.; Bobak, L.; Nowicka, P. Anti-oxidant and anti-enzymatic activities of sea buckthorn (Hippopha rhamnoides L.) fruits modulated by chemical components. Antioxidants 2019, 8, 618. [Google Scholar] [CrossRef] [Green Version]
- Meléndez-Martínez, A.J.; Escudero-Gilete, M.L.; Vicario, I.M.; Heredia, F.J. Study of the influence of carotenoid structure and individual carotenoids in the qualitative and quantitative attributes of orange juice colour. Food Res. Int. 2010, 43, 1289–1296. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Bao, J.S.; Cai, Y.Z.; Sun, M.; Wang, G.Y.; Corke, H. Anthocyanins, flavonols and free radical scavenging activity of Chinese bayberry (Myrica rubra) extracts and their color properties and stability. J. Agric. Food Chem. 2005, 53, 2327–2332. [Google Scholar] [CrossRef]
- Hsu, B.; Coupar, I.M.; Ng, K. Antioxidant activity of hot water extract from the fruit of the Doum palm, Hyphaene thebaica. Food Chem. 2005, 98, 317–328. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kellett, M.E.; Greenspan, P.; Pegg, R.B. Modification of the cellular antioxidant activity (CAA) assay to study phenolic antioxidants in a Caco-2 cell line. Food Chem. 2017, 244, 359–363. [Google Scholar] [CrossRef] [PubMed]
- Tiitinen, K.M.; Yang, B.; Haraldsson, G.G.; Jonsdottir, S.; Kallio, H.P. Fast analysis of sugars, fruit acids, and vitamin C in sea buckthorn (Hippophae rhamnoides L.) varieties. J. Agric. Food Chem. 2006, 54, 2508–2513. [Google Scholar] [CrossRef] [PubMed]
- Fatima, T.; Kesari, V.; Watt, I.; Wishart, D.; Todd, J.F.; Schroeder, W.R.; Paliyath, G.; Krishna, P. Metabolite profiling and expression analysis of flavonoid, vitamin C and tocopherol biosynthesis genes in the antioxidant-rich sea buckthorn (Hippophae rhamnoides L.). Phytochemistry 2015, 118, 181–191. [Google Scholar] [CrossRef]
- Kumar, V.; Sharma, A.; Bhardwaj, R.; Thukral, A.K. Analysis of organic acids of tricarboxylic acid cycle in plants using GC-MS, and system modeling. J. Anal. Sci. Technol. 2017, 8, 20. [Google Scholar] [CrossRef]
- Carrington, Y.; Guo, J.; Le, C.H.; Fillo, A.; Kwon, J.; Tran, L.; Ehlting, J. Evolution of a secondary metabolic pathway from primary metabolism: Shikimate and quinate biosynthesis in plants. Plant J. 2018, 95, 823–833. [Google Scholar] [CrossRef]
- Acosta-Estrada, B.A.; Gutiérrez-Uribe, J.A.; Serna-Saldívar, S.O. Bound phenolics in foods, a review. Food Chem. 2014, 152, 46–55. [Google Scholar] [CrossRef]
- Vogt, T. Phenylpropanoid biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, C.Y.; Zhang, G.Y.; Zhang, J.G.; Zeng, Y.F.; Liu, J.Y. Integrated analysis of multiomic data reveals the role of the antioxidant network in the quality of sea buckthorn berry. FASEB J. 2017, 31, 1929–1938. [Google Scholar] [CrossRef] [Green Version]
- Fan, H.L.; Bai, S.W.; Zhang, F.Q.; Wang, J. Main active components and ROS scavenging effects of five Hippophae rhamnoides L. cultivars from Hexi corridor. Nat. Prod. Res. Dev. 2016, 28, 107–111, 163. Available online: http://www.trcw.ac.cn/CN/10.16333/j.1001-6880.2016.1.019 (accessed on 15 January 2016). (In Chinese with English abstract).
- Mène-Saffrané, L. Vitamin E biosynthesis and its regulation in plants. Antioxidants 2018, 7, 2. [Google Scholar] [CrossRef]
- Pop, R.M.; Weesepoel, Y.; Socaciu, C.; Pintea, A.; Vincken, J.P.; Gruppen, H. Carotenoid composition of berries and leaves from six Romanian sea buckthorn (Hippophae rhamnoides L.) varieties. Food Chem. 2014, 147, 1460–1466. [Google Scholar] [CrossRef]
- Huang, H.Z.; Sun, Y.J.; Lou, S.T.; Li, H.; Ye, X.Q. In vitro digestion combined with cellular assay to determine the antioxidant activity in Chinese bayberry (Myrica rubra Sieb. et Zucc.) fruits: A comparison with traditional methods. Food Chem. 2014, 146, 363–370. [Google Scholar] [CrossRef]
- Ji, M.Y.; Gong, X.; Li, X.; Wang, C.C.; Li, M.H. Advanced research on the antioxidant activity and mechanism of polyphenols from Hippophae species—A review. Molecules 2020, 25, 917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagarajan, S.; Nagarajan, R.; Kumar, J.; Salemme, A.; Togna, A.R.; Saso, L.; Bruno, F. Antioxidant activity of synthetic polymers of phenolic compounds. Polymers 2020, 12, 1646. [Google Scholar] [CrossRef] [PubMed]
- Paciolla, C.; Fortunato, S.; Dipierro, N.; Paradiso, A.; Leonardis, S.D.; Mastropasqua, L.; Pinto, M.C.D. Vitamin C in plants: From functions to biofortification. Antioxidants 2019, 8, 519. [Google Scholar] [CrossRef] [Green Version]
- Irakli, K.N.; Samanidou, V.F.; Papadoyannis, I.N. Development and validation of an HPLC method for the simultaneous determination of tocopherols, tocotrienols and carotenoids in cereals after solid-phase extraction. J. Sep. Sci. 2011, 34, 1375–1382. [Google Scholar] [CrossRef]
- Rop, O.; Ercişli, S.; Mlcek, J.; Jurikova, T.; Hoza, I. Antioxidant and radical scavenging activities in fruits of 6 sea buckthorn (Hippophae rhamnoides L.) cultivars. Turk. J. Agric. For. 2014, 38, 224–232. [Google Scholar] [CrossRef]
- Chen, C.; Xu, X.M.; Chen, Y.; You, M.Y.; Wen, F.Y.; Zhan, Y. Identification, quantification, and antioxidant activity of acylated flavonol glycosides from sea buckthorn (Hippophae rhamnoides ssp. sinensis). Food Chem. 2013, 141, 1573–1579. [Google Scholar] [CrossRef]
- Guo, X.B.; Guo, R.X.; Chang, X.X.; Brennan, C.; Li, T.; Fu, X.; Liu, R.H. Phenolic compounds, antioxidant activity, antiproliferative activity and bioaccessibility of sea buckthorn (Hippophaë rhamnoides L.) berries as affected by in vitro digestion. Food Funct. 2017, 8, 4229–4240. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, K.L.; Liu, R.H. Cellular antioxidant activity (CAA) assay for assessing antioxidants, foods, and dietary supplements. J. Agric. Food Chem. 2007, 55, 8896–8907. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenolic Compounds (mg/g FW) | Free Phenolics | Bound Phenolics | Total Phenolics |
|---|---|---|---|
| Benzoic acid derivatives | |||
| Gallic acid | |||
| Rha | 0.0081 ± 0.0004 a (51.59) | 0.0076 ± 0.0003 a (48.41) | 0.0157 ± 0.0007 a |
| Tib | 0.0091 ± 0.0004 a (62.32) | 0.0054 ± 0.0002 a (37.68) | 0.0146 ± 0.0006 a |
| Protocatechuic acid | |||
| Rha | 0.0012 ± 0.0001 a (54.54) | 0.0011 ± 0.0000 a (45.46) | 0.0023 ± 0.0002 a |
| Tib | 0.0016 ± 0.0004 a (69.56) | 0.0007 ± 0.0002 a (30.44) | 0.0023 ± 0.0006 a |
| Sum | |||
| Rha | 0.0093 ± 0.0005 b (51.66) | 0.0087 ± 0.0003 a (48.34) | 0.0180 ± 0.0009 a |
| Tib | 0.0107 ± 0.0008 a (63.31) | 0.0061 ± 0.0004 b (36.69) | 0.0169 ± 0.0012 b |
| Flavanols | |||
| Catechin | |||
| Rha | 0.1083 ± 0.0064 a (80.58) | 0.0262 ± 0.0021 a (19.42) | 0.1345 ± 0.0085 a |
| Tib | 0.0472 ± 0.0069 b (76.99) | 0.0141 ± 0.0038 b (23.01) | 0.0613 ± 0.0108 b |
| Epicatechin | |||
| Rha | 0.0093 ± 0.0012 a (67.39) | 0.0044 ± 0.0000 a (32.61) | 0.0137 ± 0.0013 a |
| Tib | 0.0052 ± 0.0001 b (54.16) | 0.0043 ± 0.0000 a (45.84) | 0.0096 ± 0.0001 b |
| Sum | |||
| Rha | 0.1176 ± 0.0076 a (79.35) | 0.0306 ± 0.0021 a (20.65) | 0.1482 ± 0.0098 a |
| Tib | 0.0524 ± 0.0070 b (73.90) | 0.0184 ± 0.0038 b (26.10) | 0.0709 ± 0.0109 b |
| Flavonols | |||
| Isorhamnetin | |||
| Rha | 0.0966 ± 0.0132 a (89.69) | 0.0111 ± 0.0009 a (10.31) | 0.1077 ± 0.0142 a |
| Tib | 0.0397 ± 0.0063 b (76.19) | 0.0124 ± 0.0022 a (23.81) | 0.0521 ± 0.0085 b |
| Kaempferol | |||
| Rha | 0.0039 ± 0.0001 a (54.16) | 0.0033 ± 0.0000 a (45.84) | 0.0072 ± 0.0001 a |
| Tib | 0.0034 ± 0.0000 a (50.00) | 0.0034 ± 0.0000 a (50.00) | 0.0068 ± 0.0000 a |
| Myricetin | |||
| Rha | 0.0197 ± 0.0012 a (94.71) | 0.0011 ± 0.0000 a (5.29) | 0.0208 ± 0.0012 a |
| Tib | 0.0095 ± 0.0006 b (85.58) | 0.0015 ± 0.0002 a (14.42) | 0.0111 ± 0.0009 b |
| Quercetin | |||
| Rha | 0.0612 ± 0.0107 a (92.31) | 0.0050 ± 0.0007 a (7.69) | 0.0662 ± 0.0115 a |
| Tib | 0.0143 ± 0.0034 b (74.09) | 0.0049 ± 0.0003 a (25.91) | 0.0193 ± 0.0038 b |
| Naringenin | |||
| Rha | 0.0024 ± 0.0000 a (48.97) | 0.0025 ± 0.0000 a (51.03) | 0.0049 ± 0.0000 a |
| Tib | 0.0025 ± 0.0000 a (50.00) | 0.0025 ± 0.0000 a (50.00) | 0.0050 ± 0.0000 a |
| Sum | |||
| Rha | 0.1838 ± 0.0350 a (88.83) | 0.0230 ± 0.0022 a (16.17) | 0.2069 ± 0.0375 a |
| Tib | 0.0694 ± 0.0397 b (73.75) | 0.0247 ± 0.0081 a (26.25) | 0.0941 ± 0.0481 a |
| Phenylpropanoids | |||
| Ferulic acid | |||
| Rha | 0.0073 ± 0.0003 a (53.67) | 0.0064 ± 0.0005 a (46.33) | 0.0136 ± 0.0008 a |
| Tib | 0.0046 ± 0.0008 b (40.35) | 0.0068 ± 0.0015 a (59.65) | 0.0114 ± 0.0023 b |
| Chlorogenic acid | |||
| Rha | 0.0615 ± 0.0031 a (99.51) | 0.0003 ± 0.0001 a (0.49) | 0.0618 ± 0.0007 a |
| Tib | 0.0286 ± 0.0031 b (98.96) | 0.0003 ± 0.0000 a (1.04) | 0.0289 ± 0.0031 b |
| P-coumaric acid | |||
| Rha | 0.0329 ± 0.0009 a (80.04) | 0.0073 ± 0.0007 a (19.96) | 0.0402 ± 0.0016 a |
| Tib | 0.0079 ± 0.0015 b (48.76) | 0.0082 ± 0.0012 a (51.24) | 0.0162 ± 0.0028 b |
| Sum | |||
| Rha | 0.1016 ± 0.0043 a (87.96) | 0.0139 ± 0.0012 b (12.04) | 0.1155 ± 0.0031 a |
| Tib | 0.0411 ± 0.0054 b (72.61) | 0.0153 ± 0.0027 a (27.39) | 0.0564 ± 0.0082 b |
| Total | |||
| Rha | 0.4124 ± 0.0147 a (84.40) | 0.0762 ± 0.0033 a (15.6) | 0.4886 ± 0.0142 a |
| Tib | 0.1736 ± 0.0288 b (72.94) | 0.0645 ± 0.0032 a (27.06) | 0.2383 ± 0.0037 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, T.; Wei, J.; Zhao, J.; Jiang, Y.; Bi, Y.; George, G. Comparative Assessment of Functional Components and Antioxidant Activities between Hippophae rhamnoides ssp. sinensis and H. tibetana Berries in Qinghai–Tibet Plateau. Foods 2023, 12, 341. https://doi.org/10.3390/foods12020341
Su T, Wei J, Zhao J, Jiang Y, Bi Y, George G. Comparative Assessment of Functional Components and Antioxidant Activities between Hippophae rhamnoides ssp. sinensis and H. tibetana Berries in Qinghai–Tibet Plateau. Foods. 2023; 12(2):341. https://doi.org/10.3390/foods12020341
Chicago/Turabian StyleSu, Tingting, Juan Wei, Jinmei Zhao, Yumei Jiang, Yang Bi, and Galitsyn George. 2023. "Comparative Assessment of Functional Components and Antioxidant Activities between Hippophae rhamnoides ssp. sinensis and H. tibetana Berries in Qinghai–Tibet Plateau" Foods 12, no. 2: 341. https://doi.org/10.3390/foods12020341
APA StyleSu, T., Wei, J., Zhao, J., Jiang, Y., Bi, Y., & George, G. (2023). Comparative Assessment of Functional Components and Antioxidant Activities between Hippophae rhamnoides ssp. sinensis and H. tibetana Berries in Qinghai–Tibet Plateau. Foods, 12(2), 341. https://doi.org/10.3390/foods12020341