
Grapevine Shoot Extract Rich in Trans-Resveratrol and Trans-ε-Viniferin: Evaluation of Their Potential Use for Cardiac Health

 ,
, 
 ,
,  , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Raw Biomass and Extraction
2.2. Characterization of the Extract
2.2.1. Phenolic Content, Composition, and Antioxidant Assays
2.2.2. Other Components
2.3. In Vivo Evaluation of the Cardiac Protective Activity
2.3.1. Animals
2.3.2. Acute Preliminary Toxicity Effect
2.3.3. Experimental In Vivo Assay
2.3.4. Electrocardiogram
2.3.5. Biological Sample Collection
2.3.6. Analysis of Cardiac Parameters in Plasma
2.3.7. Determination of Cardiac Pro-Antioxidant and Antioxidants Levels
2.3.8. DNA Fragmentation Analysis
2.3.9. Determination of Heart Pro-Apoptotic Genes
2.3.10. Infarct Size Determination
2.3.11. Histopathological Analysis
2.4. Statistical Analysis
3. Results
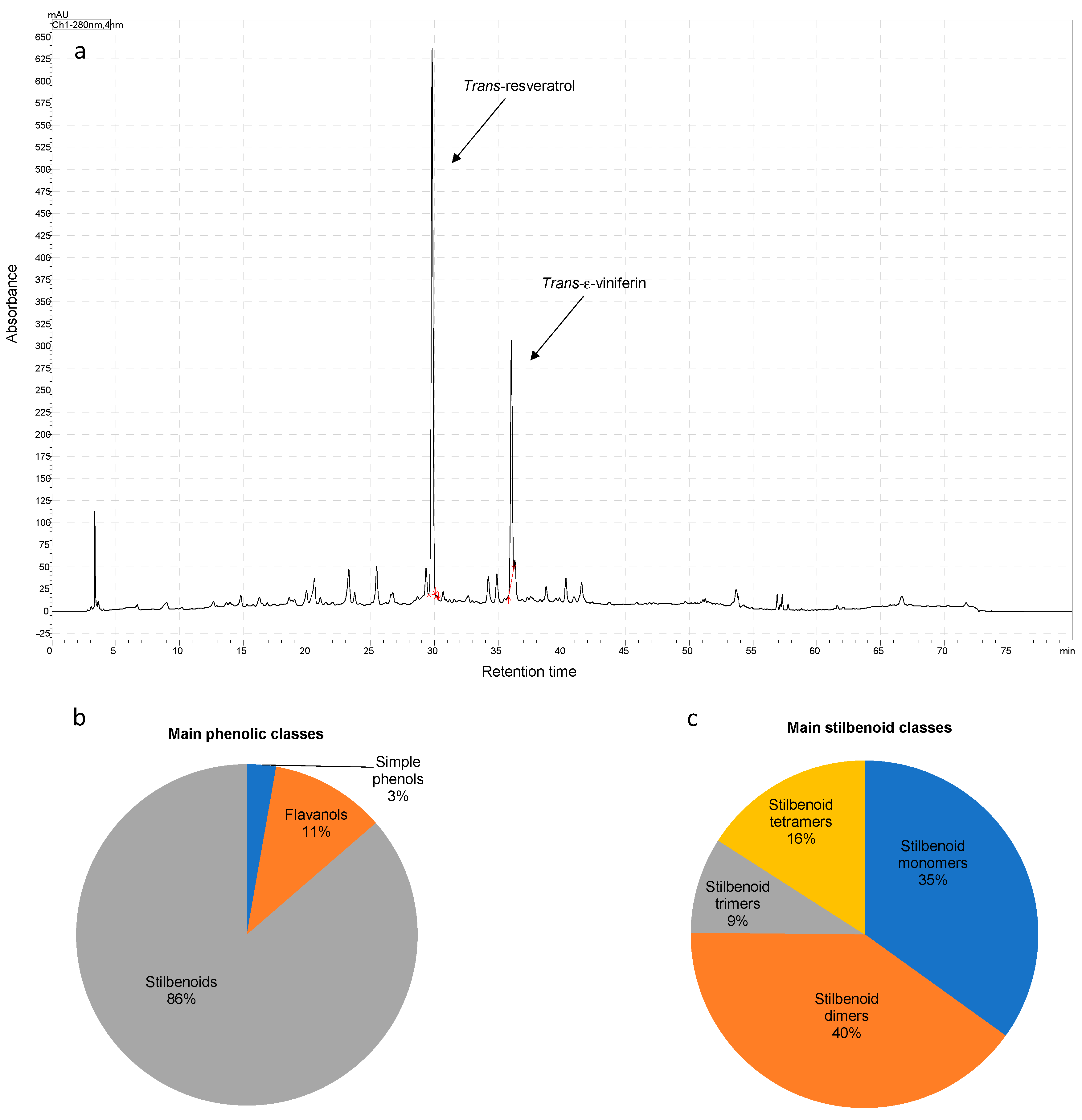
3.1. Phenolic Composition and Antioxidant Potency
3.2. Other Components of GSE
3.3. Preliminary Acute Toxicity Evaluation
3.4. Effect of GSE on Rats’ Weight
3.5. Effect of GSE on ST-Segment
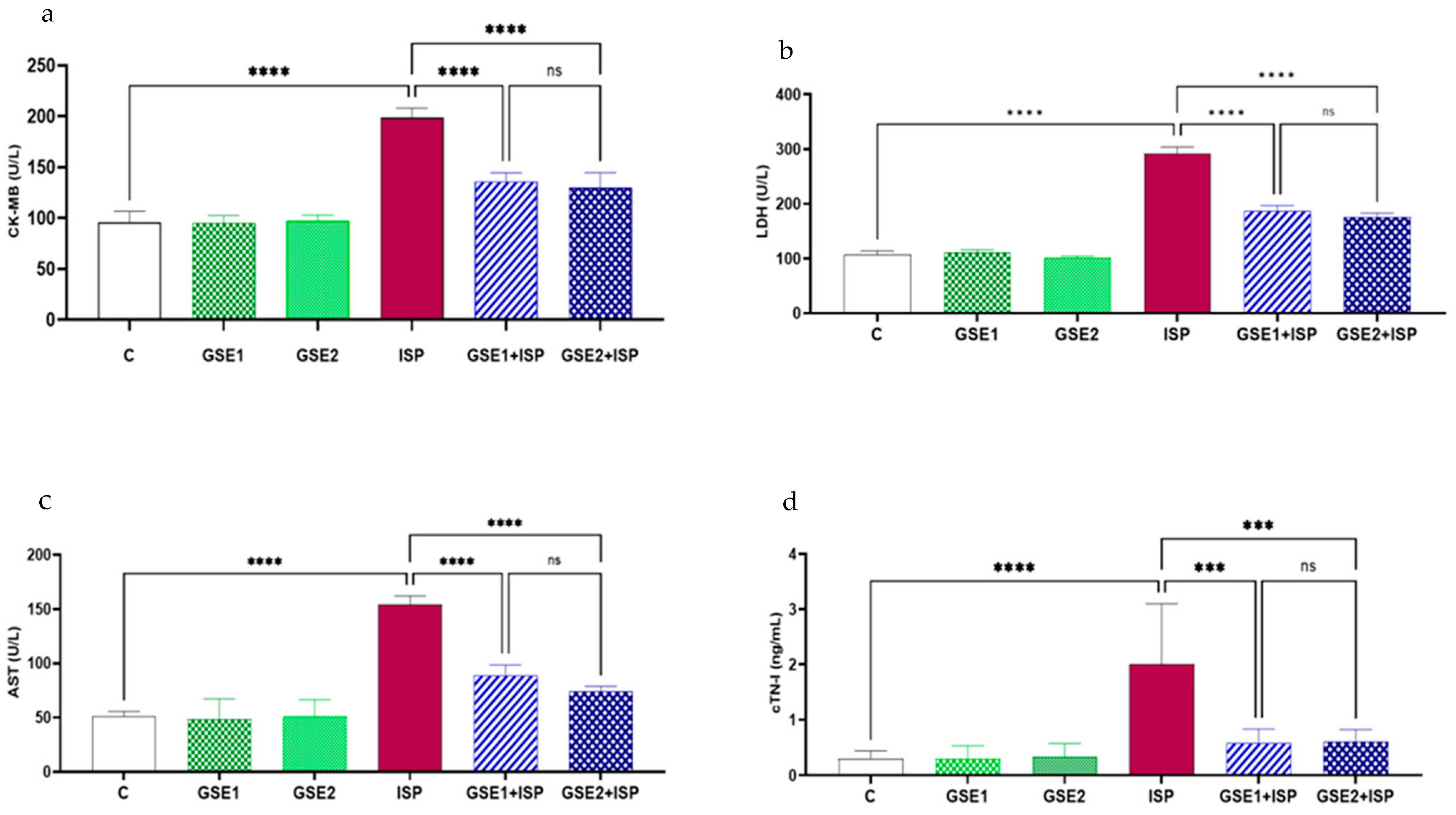
3.6. Effect of GSE on the Cardiac Indicators of Injury
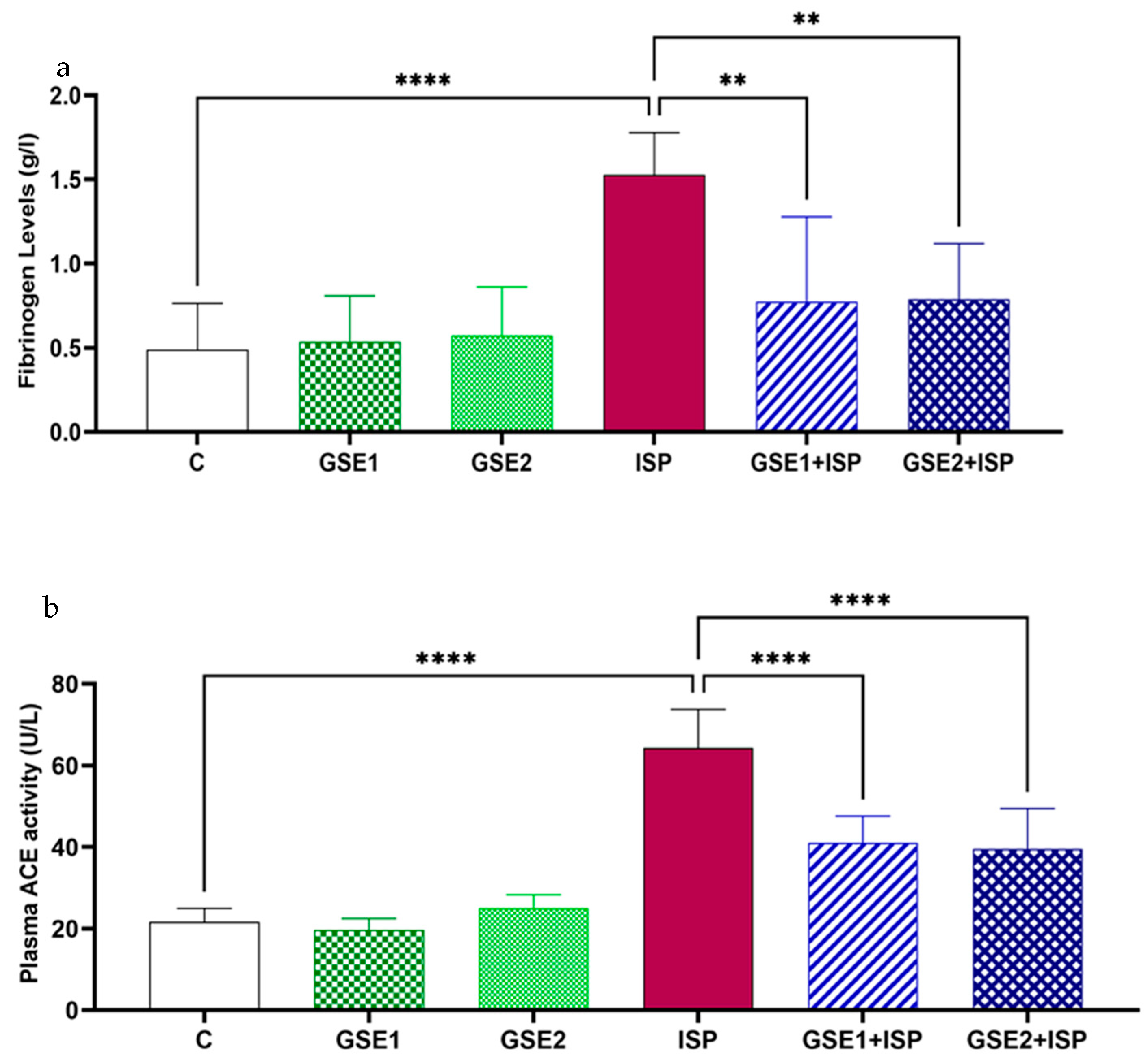
3.7. Effect of GSE on Fibrinogen Concentration
3.8. Effect of GSE on ACE
3.9. Effect of GSE on Plasmatic Lipids
3.10. Effect of GSE on Major Plasma Electrolytes
3.11. Effect of GSE on MDA, CAT, SOD, and GSH Levels
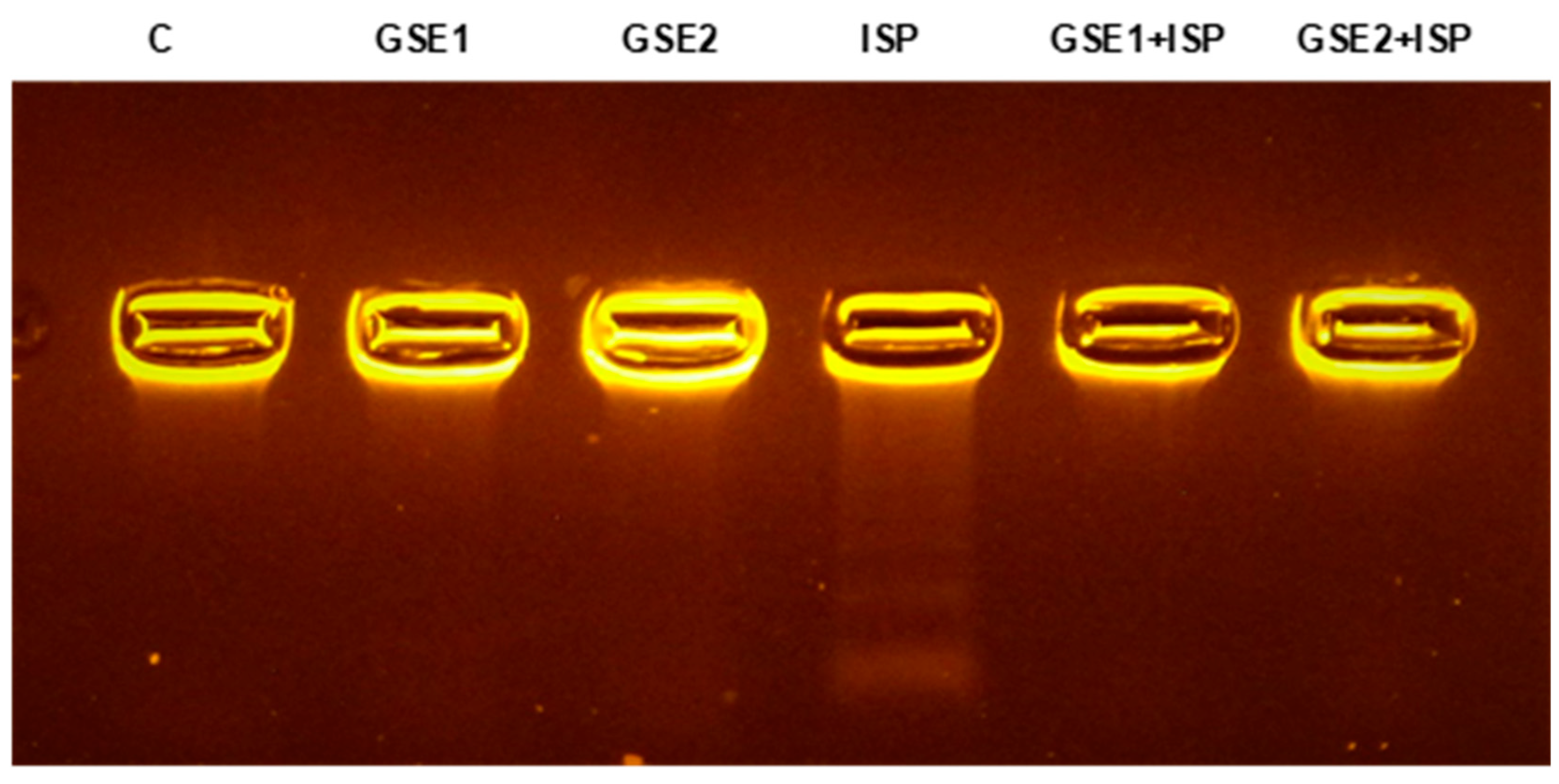
3.12. Effect of GSE on the DNA from Cardiac Tissue
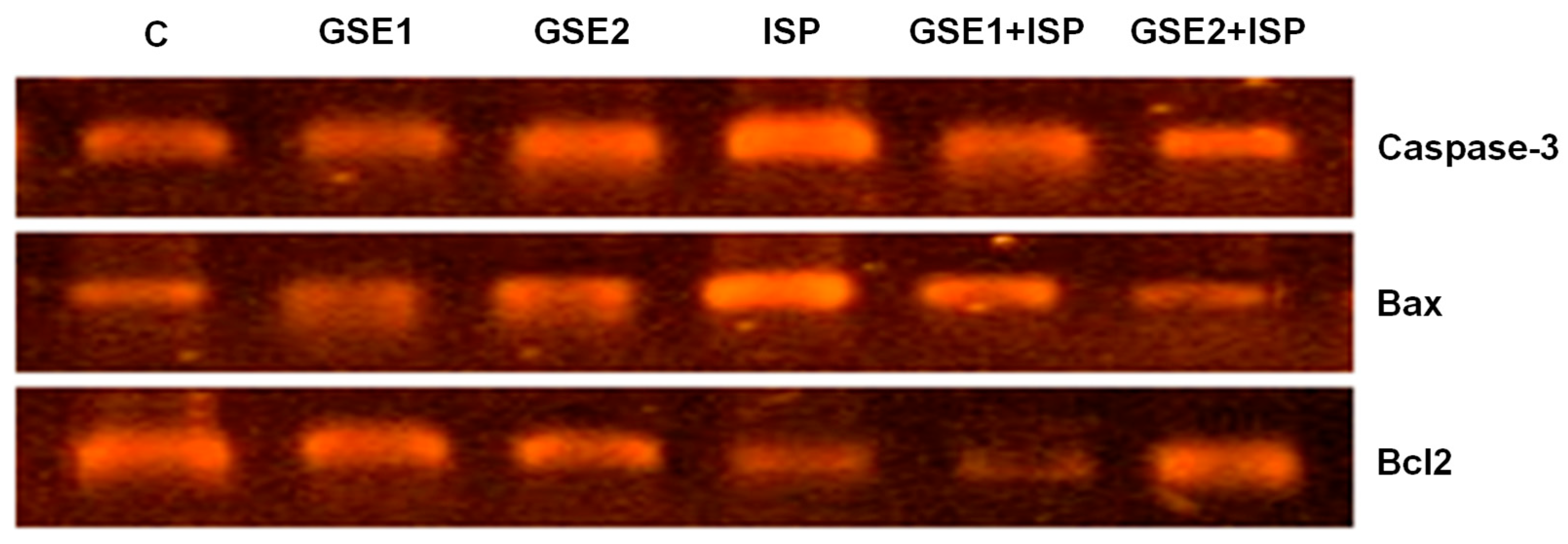
3.13. Effect of GSE on Pro-Apoptotic Genes
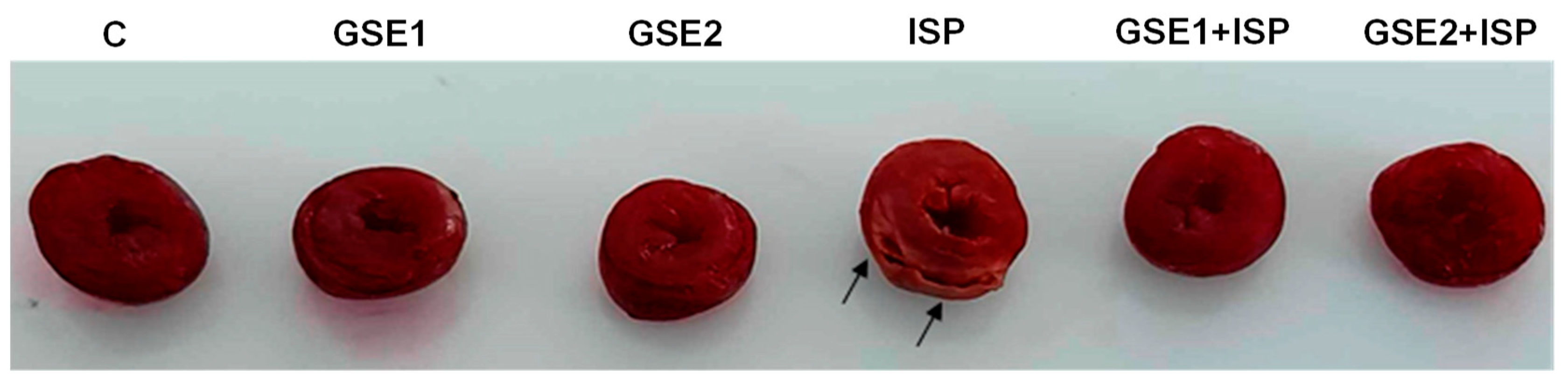
3.14. Infarct Size Staining
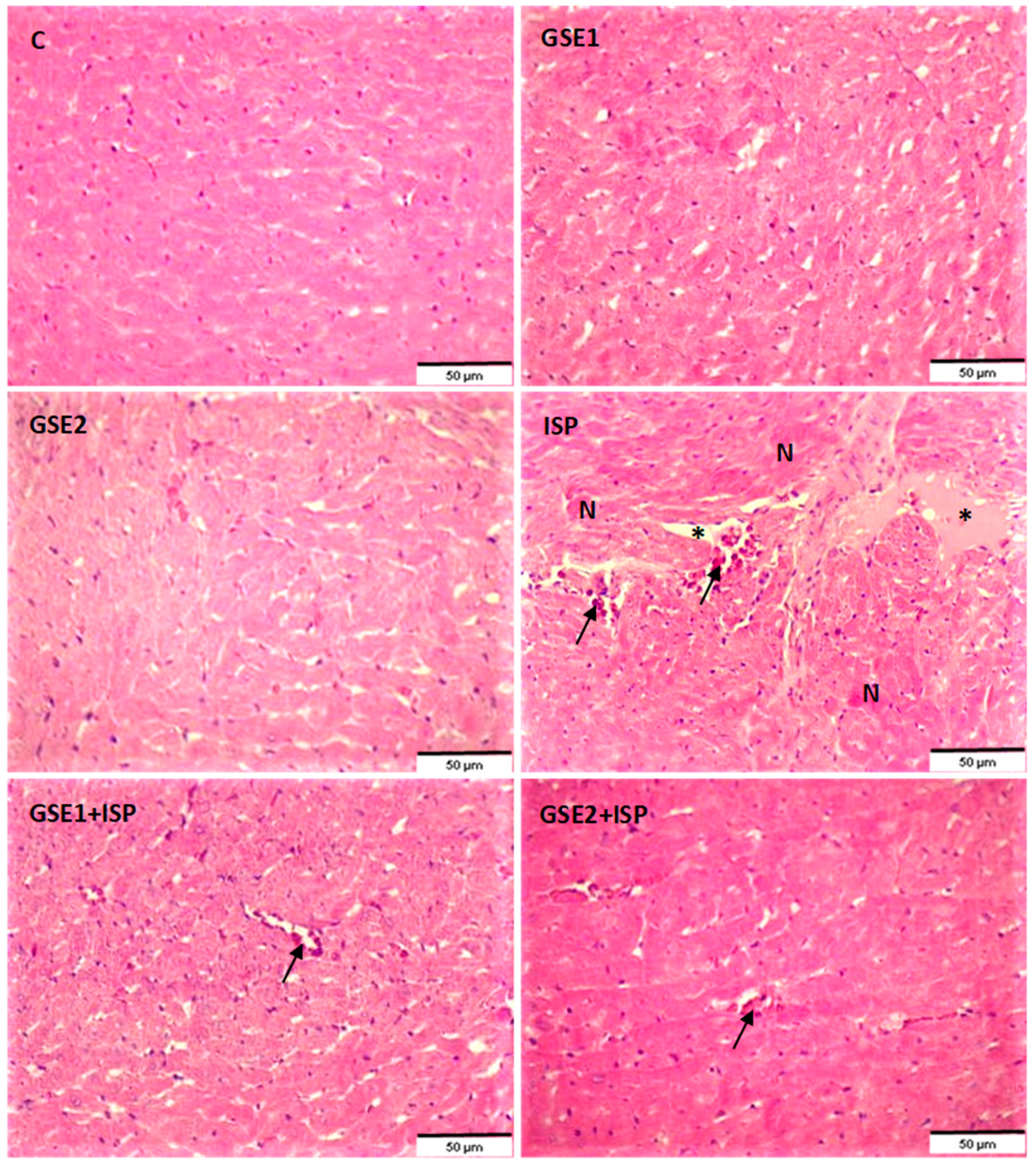
3.15. Effect of GSE on the Histology of the Heart Tissue
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- OIV. State of the World Vine and Wine Sector in 2022. Available online: https://www.oiv.int/sites/default/files/documents/2023-04_Press_Conf.pdf (accessed on 1 September 2023).
- Pérez-Bermúdez, P.; Olmo, M.; Gil, J.; García-Ferriz, L.; Olmo, C.; Boluda, R.; Gavidia, I. Effects of traditional and light pruning on viticultural and oenological performance of Bobal and Tempranillo vineyards. OENO One 2015, 49, 145–154. [Google Scholar] [CrossRef]
- Contreras, M.d.M.; Romero-García, J.M.; López-Linares, J.C.; Romero, I.; Castro, E. Residues from grapevine and wine production as feedstock for a biorefinery. Food Bioprod. Process. 2022, 134, 56–79. [Google Scholar] [CrossRef]
- Aliaño-González, M.J.; Richard, T.; Cantos-Villar, E. Grapevine Cane extracts: Raw plant material, extraction methods, quantification, and applications. Biomolecules 2020, 10, 1195. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Torre, M.P.; Ferreiro-Vera, C.; Priego-Capote, F.; Pérez-Juan, P.M.; Luque De Castro, M.D. Comparison of accelerated methods for the extraction of phenolic compounds from different vine-shoot cultivars. J. Agric. Food Chem. 2012, 60, 3051–3060. [Google Scholar] [CrossRef]
- Goufo, P.; Singh, R.K.; Cortez, I. A reference list of phenolic compounds (including stilbenes) in grapevine (Vitis vinifera L.) roots, woods, canes, stems, and leaves. Antioxidants 2020, 9, 398. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Gómez, R.; Garde-Cerdán, T.; Zalacain, A.; Garcia, R.; Cabrita, M.J.; Salinas, M.R. Vine-shoot waste aqueous extract applied as foliar fertilizer to grapevines: Effect on amino acids and fermentative volatile content. Food Chem. 2016, 197, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Dorosh, O.; Moreira, M.M.; Rodrigues, F.; Peixoto, A.F.; Freire, C.; Morais, S.; Delerue-Matos, C. Vine-canes valorisation: Ultrasound-assisted extraction from lab to pilot scale. Molecules 2020, 25, 1739. [Google Scholar] [CrossRef]
- Noviello, M.; Caputi, A.F.; Squeo, G.; Paradiso, V.M.; Gambacorta, G.; Caponio, F. Vine shoots as a source of trans-resveratrol and ε-Viniferin: A Study of 23 Italian varieties. Foods 2022, 11, 553. [Google Scholar] [CrossRef]
- Loupit, G.; Prigent, S.; Franc, C.; De Revel, G.; Richard, T.; Cookson, S.J.; Fonayet, J.V. Polyphenol profiles of just pruned grapevine canes from wild Vitis Accessions and Vitis vinifera cultivars. J. Agric. Food Chem. 2020, 68, 13397–13407. [Google Scholar] [CrossRef]
- Rajha, H.N.; El Kantar, S.; Afif, C.; Boussetta, N.; Louka, N.; Maroun, R.G.; Vorobiev, E. Selective multistage extraction process of biomolecules from vine shoots by a combination of biological, chemical, and physical treatments. Comptes Rendus Chim. 2018, 21, 581–589. [Google Scholar] [CrossRef]
- Moreira, M.M.; Barroso, M.F.; Porto, J.V.; Ramalhosa, M.J.; Švarc-Gajić, J.; Estevinho, L.; Morais, S.; Delerue-Matos, C. Potential of Portuguese vine shoot wastes as natural resources of bioactive compounds. Sci. Total Environ. 2018, 634, 831–842. [Google Scholar] [CrossRef]
- Raj, P.; Thandapilly, S.J.; Wigle, J.; Zieroth, S.; Netticadan, T. A Comprehensive analysis of the efficacy of resveratrol in atherosclerotic cardiovascular disease, myocardial infarction and heart failure. Molecules 2021, 26, 6600. [Google Scholar] [CrossRef]
- Gómez-Cruz, I.; Contreras, M.d.M.; Romero, I.; Ribeiro, B.; Roseiro, L.B.; Duarte, L.C.; Carvalheiro, F.; Castro, E. Strategies for the purification of hydroxytyrosol-rich extracts obtained from exhausted olive pomace. Sep. Purif. Technol. 2023, 325, 124664. [Google Scholar] [CrossRef]
- Contreras, M.d.M.; Gómez-Cruz, I.; Feriani, A.; Alwasel, S.; Harrath, A.H.; Romero, I.; Castro, E.; Tlili, N. Hepatopreventive properties of hydroxytyrosol and mannitol-rich extracts obtained from exhausted olive pomace using green extraction methods. Food Funct. 2022, 18, 11915–11928. [Google Scholar] [CrossRef]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1978; pp. 302–310. [Google Scholar]
- Aebi, H. Catalase in Vitro. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1985; pp. 121–126. [Google Scholar]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Ellman, G.L. Tissue Sulfhydryl Groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Chtourou, Y.; Aouey, B.; Kebieche, M.; Fetoui, H. Protective role of naringin against cisplatin induced oxidative stress, inflammatory response and apoptosis in rat striatum via suppressing ROS-mediated NF-ΚB and P53 signaling pathways. Chem. Biol. Interact. 2015, 239, 76–86. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Feriani, A.; Tir, M.; Gómez-Caravaca, A.M.; Contreras, M.d.M.; Talhaoui, N.; Taamalli, A.; Segura-Carretero, A.; Ghazouani, L.; Mufti, A.; Tlili, N.; et al. HPLC-DAD-ESI-QTOF-MS/MS profiling of Zygophyllum album roots extract and assessment of its cardioprotective effect against deltamethrin-induced myocardial injuries in rat, by suppression of oxidative stress-related inflammation and apoptosis via NF-ΚB signaling pathway. J. Ethnopharmacol. 2020, 247, 112266. [Google Scholar] [CrossRef]
- Feriani, A.; Khdhiri, E.; Tir, M.; Elmufti, A.; Tlili, N.; Hajji, R.; Ammar, H.; Allouche, N.; Abid, S.; Ghazouani, L.; et al. (E)-N ′-(1-(7-Hydroxy-2-Oxo-2H-Chromen-3-Yl) Ethylidene) benzohydrazide, a novel synthesized coumarin, ameliorates isoproterenol-induced myocardial infarction in rats through attenuating oxidative stress, inflammation, and apoptosis. Oxid. Med. Cell. Longev. 2020, 2020, 2432918. [Google Scholar] [CrossRef]
- Gabaston, J.; Leborgne, C.; Waffo-Teguo, P.; Valls, J.; Palos Pinto, A.; Richard, T.; Cluzet, S.; Mérillon, J.M. Wood and roots of major grapevine cultivars and rootstocks: A comparative analysis of stilbenes by UHPLC-DAD-MS/MS and NMR. Phytochem. Anal. 2019, 30, 320–331. [Google Scholar] [CrossRef] [PubMed]
- Attard, T.M.; Bukhanko, N.; Eriksson, D.; Arshadi, M.; Geladi, P.; Bergsten, U.; Budarin, V.L.; Clark, J.H.; Hunt, A.J. Supercritical extraction of waxes and lipids from biomass: A valuable first step towards an integrated biorefinery. J. Clean. Prod. 2018, 177, 684–698. [Google Scholar] [CrossRef]
- Weiskirchen, S.; Weiskirchen, R. Resveratrol: How much wine do you have to drink to stay healthy? Adv. Nutr. 2016, 7, 706–718. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Vlaisavljevic, S.; Adetunji, C.O.; Adetunji, J.B.; Kregiel, D.; Antolak, H.; Pawlikowska, E.; Uprety, Y.; Mileski, K.S.; Devkota, H.P.; et al. Plants of the genus Vitis: Phenolic compounds, anticancer properties and clinical relevance. Trends Food Sci. Technol. 2019, 91, 362–379. [Google Scholar] [CrossRef]
- Rayne, S.; Karacabey, E.; Mazza, G. Grape Cane Waste as a Source of trans-resveratrol and trans-viniferin: High-value phytochemicals with medicinal and anti-phytopathogenic applications. Ind. Crops Prod. 2008, 27, 335–340. [Google Scholar] [CrossRef]
- Jarosova, V.; Vesely, O.; Doskocil, I.; Tomisova, K.; Marsik, P.; Jaimes, J.D.; Smejkal, K.; Kloucek, P.; Havlik, J. Metabolism of cis-and trans-resveratrol and dihydroresveratrol in an intestinal epithelial model. Nutrients 2020, 12, 595. [Google Scholar] [CrossRef]
- Aboubakr, M.; Elsayd, F.; Soliman, A.; Fadl, S.E.; El-Shafey, A.; Abdelhiee, E.Y. L-Carnitine and vitamin E ameliorate cardiotoxicity induced by tilmicosin in rats. Environ. Sci. Pollut. Res. 2020, 27, 23026–23034. [Google Scholar] [CrossRef]
- Feriani, A.; Tir, M.; Aldahmash, W.; Mnafgui, K.; Hichem, A.; Gómez-Caravaca, A.M.; Contreras, M.d.M.; Taamalli, A.; Alwasel, S.; Segura-Carretero, A.; et al. In vivo evaluation and molecular docking studies of Schinus molle L. fruit extract protective effect against isoproterenol-induced infarction in rats. Environ. Sci. Pollut. Res. 2022, 29, 80910–80925. [Google Scholar] [CrossRef]
- Vijayakumar, R.; Nachiappan, V. Cassia Auriculata Flower Extract Attenuates Hyperlipidemia in Male Wistar Rats by Regulating the Hepatic Cholesterol Metabolism. Biomed. Pharmacother. 2017, 95, 394–401. [Google Scholar] [CrossRef]
- Venkadeswaran, K.; Thomas, P.A.; Geraldine, P. An experimental evaluation of the anti-atherogenic potential of the plant, piper betle, and its active constitutent, eugenol, in rats fed an atherogenic diet. Biomed. Pharmacother. 2016, 80, 276–288. [Google Scholar] [CrossRef]
- Roy, S.J.; Mainzen Prince, P.S. Protective Effects of Sinapic Acid on Cardiac Hypertrophy, Dyslipidaemia and Altered Electrocardiogram in Isoproterenol-Induced Myocardial Infarcted Rats. Eur. J. Pharmacol. 2013, 699, 213–218. [Google Scholar] [CrossRef]
- Fan, S.; Hu, Y.; You, Y.; Xue, W.; Chai, R.; Zhang, X.; Shou, X.; Shi, J. Role of resveratrol in inhibiting pathological cardiac remodeling. Front. Pharmacol. 2022, 13, 3452. [Google Scholar] [CrossRef] [PubMed]
- Mnafgui, K.; Khdhiri, E.; Hajji, R.; Feriani, A.; Ivan da Silva, F.; Laíres da Silva Santos, A.; Tlili, A.; Mlayeh, S.; Bouzidi, M.; Ammar, H.; et al. Potential effect of new (E)-4-hydroxy-N’-(1-(7-hydroxy-2-oxo-2H-chromen-3-Yl) ethylidene) benzohydrazide against acute myocardial infarction: Haemodynamic, biochemical and histological studies. Clin. Exp. Pharmacol. Physiol. 2021, 48, 107–120. [Google Scholar] [CrossRef]
- Fernández-Castillejo, S.; Macià, A.; Motilva, M.J.; Catalán, Ú.; Solà, R. Endothelial cells deconjugate resveratrol metabolites to free resveratrol: A possible role in tissue factor modulation. Mol. Nutr. Food Res. 2019, 63, 1800715. [Google Scholar] [CrossRef] [PubMed]
- Ardjmand, A.; Shahaboddin, M.E.; Mazoochi, T.; Heydari, A.; Ghavipanjeh, G. Cardioprotective effects of cerebrolysin on the lesion severity and inflammatory factors in a rat model of isoproterenol-induced myocardial injury. Pharmacol. Rep. 2019, 71, 682–687. [Google Scholar] [CrossRef] [PubMed]
- Feriani, A.; Contreras, M.d.M.; Talhaoui, N.; Gómez-Caravaca, A.M.; Taamalli, A.; Segura-Carretero, A.; Ghazouani, L.; El Feki, A.; Allagui, M.S. Protective effect of Globularia alypum leaves against deltamethrin-induced nephrotoxicity in rats and determination of its bioactive compounds using high-performance liquid chromatography coupled with electrospray ionization tandem quadrupole–time-of-flight. J. Funct. Foods 2017, 32, 139–148. [Google Scholar] [CrossRef]
- Paulino, E.T.; Barros Ferreira, A.K.; da Silva, J.C.G.; Ferreira Costa, C.D.; Smaniotto, S.; de Araújo-Júnior, J.X.; Silva Júnior, E.F.; Bortoluzzi, J.H.; Nogueira Ribeiro, Ê.A. Cardioprotective effects induced by hydroalcoholic extract of leaves of Alpinia zerumbet on myocardial infarction in rats. J. Ethnopharmacol. 2019, 242, 112037. [Google Scholar] [CrossRef]
- Eladwy, R.A.; Mantawy, E.M.; El-Bakly, W.M.; Fares, M.; Ramadan, L.A.; Azab, S.S. Mechanistic insights to the cardioprotective effect of blueberry nutraceutical extract in isoprenaline-induced cardiac hypertrophy. Phytomedicine 2018, 51, 84–93. [Google Scholar] [CrossRef]
- Kronenbitter, A.; Funk, F.; Hackert, K.; Gorreßen, S.; Glaser, D.; Boknik, P.; Poschmann, G.; Stühler, K.; Isić, M.; Krüger, M.; et al. Impaired Ca2+ cycling of nonischemic myocytes contributes to sarcomere dysfunction early after myocardial infarction. J. Mol. Cell. Cardiol. 2018, 119, 28–39. [Google Scholar] [CrossRef]
- Yang, S.; Chou, G.; Li, Q. Cardioprotective role of azafrin in against myocardial injury in rats via activation of the Nrf2-ARE Pathway. Phytomedicine 2018, 47, 12–22. [Google Scholar] [CrossRef]
- Han, X.; Li, M.; Zhao, Z.; Zhang, Y.; Zhang, J.; Zhang, X.; Zhang, Y.; Guan, S.; Chu, L. mechanisms underlying the cardio-protection of total ginsenosides against myocardial ischemia in rats in vivo and in vitro: Possible involvement of L-type Ca 2+ channels, contractility and Ca 2+ homeostasis. J. Pharmacol. Sci. 2019, 139, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Kang, N.N.; Fu, L.; Xu, J.; Han, Y.; Cao, J.X.; Sun, J.F.; Zheng, M. Testosterone improves cardiac function and alters angiotensin ii receptors in isoproterenol-induced heart failure. Arch. Cardiovasc. Dis. 2012, 105, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Aybastıer, Ö.; Dawbaa, S.; Demir, C. Investigation of antioxidant ability of grape seeds extract to prevent oxidatively induced DNA damage by gas chromatography-tandem mass spectrometry. J. Chromatogr. B 2018, 1072, 328–335. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Content |
|---|---|
| TPC (mg gallic acid eq./g GSE) | 207.3 ± 3.5 |
| Trans-resveratrol (mg/g GSE) | 27.4 ± 0.3 |
| Trans-ε-viniferin (mg/g GSE) | 29.2 ± 0.2 |
| FRAP (mg Trolox eq./g GSE) | 137.58 ± 3.57 |
| Proposed Compound | RT (min) | [M-H]- m/z | Molecular Formula | Score | Error (ppm) | Phenolic Class |
|---|---|---|---|---|---|---|
| Gallic acid | 0.8 | 169.014 | C7H6O5 | 98.9 | −1.0 | Hydroxybenzoic acid |
| Dihydroxybenzaldehyde | 2.1 | 137.025 | C7H6O3 | 99.2 | −0.7 | Phenol aldehyde |
| Hydroxybenzaldehyde | 3.4 | 121.030 | C7H6O2 | 99.7 | −0.7 | Phenol aldehyde |
| (Epi)catechin B-type dimer | 4.4 | 577.135 | C30H26O12 | 91.6 | −1.3 | Flavanol dimer |
| Catechin | 4.4 | 289.073 | C15H14O6 | 94.7 | −3.0 | Flavanol monomer |
| Epicatechin | 6.5 | 289.072 | C15H14O6 | 97.0 | −2.0 | Flavanol monomer |
| Oxidized resveratrol dimer 1 | 7.6 | 471.145 | C28H24O7 | 98.4 | 0.5 | Stilbenoid dimer |
| Oxidized resveratrol dimer 2 | 7.9 | 471.146 | C28H24O7 | 96.3 | −2.0 | Stilbenoid dimer |
| Resveratrol hexoside | 9.3 | 389.125 | C20H22O8 | 96.0 | −1.9 | Stilbenoid monomer |
| Oxidized resveratrol dimer 3 | 9.4 | 471.145 | C28H24O7 | 95.8 | −1.2 | Stilbenoid dimer |
| (Epi)catechin gallate | 9.6 | 441.084 | C22H18O10 | 97.7 | −1.8 | Flavanol monomer |
| Piceatannol | 9.9 | 243.067 | C14H12O4 | 96.7 | −1.4 | Stilbenoid monomer |
| Ampelopsin A | 10.5 | 469.129 | C28H22O7 | 97.5 | −0.4 | Stilbenoid dimer |
| trans-Resveratrol | 12.3 | 227.072 | C14H12O3 | 96.6 | −2.0 | Stilbenoid monomer |
| Pallidol | 12.7 | 453.135 | C28H22O6 | 97.1 | −1.8 | Stilbenoid dimer |
| Resveratrol isomer | 14.0 | 227.072 | C14H12O3 | 98.2 | −0.7 | Stilbenoid monomer |
| Viniferol E | 15.5 | 923.270 | C56H44O13 | 97.8 | 1.3 | Stilbenoid tetramer |
| Hopeaphenol | 16.1 | 905.260 | C56H42O12 | 98.7 | 0.6 | Stilbenoid tetramer |
| Viniferin isomer 1 | 16.6 | 453.135 | C28H22O6 | 96.3 | −1.3 | Stilbenoid dimer |
| Hopeaphenol isomer | 16.6 | 905.260 | C56H42O12 | 94.0 | 0.0 | Stilbenoid tetramer |
| Vitisinol C | 17.2 | 427.155 | C27H24O5 | 98.3 | −0.4 | Stilbenoid dimer |
| trans-ε-Viniferin | 17.6 | 453.135 | C28H22O6 | 96.6 | −0.2 | Stilbenoid dimer |
| Miyabenol C | 18.8 | 679.198 | C42H32O9 | 97.0 | −0.6 | Stilbenoid trimer |
| Viniferin isomer 2 | 18.9 | 453.135 | C28H22O6 | 98.4 | −1.4 | Stilbenoid dimer |
| α-Viniferin | 19.9 | 677.183 | C42H30O9 | 90.0 | −2.0 | Stilbenoid trimer |
| Vitisin B isomer 1 | 23.2 | 905.259 | C56H42O12 | 96.5 | 1.9 | Stilbenoid tetramer |
| Vitisin B isomer 2 | 23.4 | 905.259 | C56H42O12 | 98.3 | 1.3 | Stilbenoid tetramer |
| Component | Content |
|---|---|
| Ultimate analysis (%, g/100 g GSE) | |
| Carbon | 52.87 ± 0.11 |
| Hydrogen | 6.69 ± 0.01 |
| Nitrogen | 1.06 ± 0.03 |
| Sulfur | 0.07 ± 0.09 |
| Other components (%, g/100 g GSE) | |
| Moisture | 9.24 ± 0.11 |
| Ash | 6.33 ± 0.08 |
| Other components (mg/g GSE) | |
| Protein 2 | 66.3 ± 1.9 |
| Oligomeric glucose | 59.8 ± 4.4 |
| Oligomeric galactose | 3.1 ± 0.3 |
| Monomeric glucose | 55.7 ± 5.3 |
| Monomeric galactose | 5.0 ± 1.0 |
| Mannitol | 27.8 ± 2.3 |
| Organic acids 1 | 38.9 ± 0.5 |
| Parameter | C | GSE1 | GSE2 | ISP | GSE1 + ISP | GSE2 + ISP |
|---|---|---|---|---|---|---|
| Body weight (g) | 285.5 ± 2 | 291.8 ± 2.9 | 289.9 ± 0.9 | 282.7 ± 4.2 | 291.4 ± 2.1 | 286.4 ± 4.2 |
| Heart weight (g) | 0.73 ± 0.10 | 0.86 ± 0.05 | 0.80 ± 0.12 | 1.74 ± 0.10 **** | 1.07 ± 0.11 *** | 0.89 ± 0.10 *** |
| CWI | 0.25 ± 0.02 | 0.29 ± 0.03 | 0.27 ± 0.03 | 0.61 ± 0.06 **** | 0.36 ± 0.07 *** | 0.31 ± 0.01 *** |
| Parameter | C | GSE1 | GSE2 | ISP | GSE1 + ISP | GSE2 + ISP |
|---|---|---|---|---|---|---|
| Plasma lipids | ||||||
| TC (mg/dL) | 67.6 ± 2.0 | 70.4 ± 3.6 | 70.8 ± 7.8 | 142.0 ± 6.5**** | 82.4 ± 9.9 **** | 85.7 ± 4.0 **** |
| TG (mg/dL) | 32.8 ± 3.6 | 30.5 ± 4.9 | 35.1 ± 4.6 | 75.4 ± 4.9 **** | 46.3 ± 10.8 **** | 40.4 ± 4.0 **** |
| LDL-C (mg/dL) | 23.9 ± 2.5 | 28.8 ± 5.8 | 25.1 ± 7.0 | 98.9 ± 10.9 **** | 52.2 ± 14.3 **** | 40.0 ± 8.5 **** |
| HDL-C (mg/dL) | 36.5 ± 6.1 | 32.4 ± 4.42 | 30.3 ± 2.8 | 17.8 ± 3.2 **** | 29.0 ± 5.7 *** | 30.5 ± 4.7 *** |
| Plasma electrolytes | ||||||
| Na+ (mmol/L) | 133.8 ± 2.3 | 130.2 ± 4.4 | 119.7 ± 19.7 | 90.97 ± 4.6 **** | 117.1 ± 8.2 ** | 122.1 ± 12.7 *** |
| Ca2+ (mmol/L) | 6.9 ± 1.0 | 6.9 ± 2.4 | 6.348 ± 4.1 | 14.4 ± 0.8 **** | 8.7 ± 2.2 ** | 8.3 ± 1.5 *** |
| TBARS and antioxidant status | ||||||
| TBARS (nmol MDA/g tissue) | 0.79 ± 0.38 | 1.19 ± 0.76 | 1.27 ± 0.41 | 6.62 ± 0.71 **** | 2.10 ± 1.02 **** | 1.97 ± 0.93 **** |
| CAT (µmol destroyed H2O2/min per mg protein) | 21.77 ± 3.38 | 21.28 ± 4.72 | 19.23 ± 4.95 | 7.11 ± 3.71 **** | 15.65 ± 3.33 ** | 18.58 ± 2.41 *** |
| SOD (U/mg protein) | 26.53 ± 3.14 | 24.07 ± 4.47 | 27.03 ± 4.89 | 11.85 ± 3.45 **** | 20.90 ± 4.98 ** | 21.62 ± 4.09 ** |
| GSH (U/mg protein) | 6.32 ± 1.09 | 6.028 ± 0.43 | 6.34 ± 0.65 | 2.15 ± 0.55 **** | 5.33 ± 0.61 **** | 5.38 ± 1.01 **** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Contreras, M.d.M.; Feriani, A.; Gómez-Cruz, I.; Hfaiedh, N.; Harrath, A.H.; Romero, I.; Castro, E.; Tlili, N. Grapevine Shoot Extract Rich in Trans-Resveratrol and Trans-ε-Viniferin: Evaluation of Their Potential Use for Cardiac Health. Foods 2023, 12, 4351. https://doi.org/10.3390/foods12234351
Contreras MdM, Feriani A, Gómez-Cruz I, Hfaiedh N, Harrath AH, Romero I, Castro E, Tlili N. Grapevine Shoot Extract Rich in Trans-Resveratrol and Trans-ε-Viniferin: Evaluation of Their Potential Use for Cardiac Health. Foods. 2023; 12(23):4351. https://doi.org/10.3390/foods12234351
Chicago/Turabian StyleContreras, María del Mar, Anouar Feriani, Irene Gómez-Cruz, Najla Hfaiedh, Abdel Halim Harrath, Inmaculada Romero, Eulogio Castro, and Nizar Tlili. 2023. "Grapevine Shoot Extract Rich in Trans-Resveratrol and Trans-ε-Viniferin: Evaluation of Their Potential Use for Cardiac Health" Foods 12, no. 23: 4351. https://doi.org/10.3390/foods12234351
APA StyleContreras, M. d. M., Feriani, A., Gómez-Cruz, I., Hfaiedh, N., Harrath, A. H., Romero, I., Castro, E., & Tlili, N. (2023). Grapevine Shoot Extract Rich in Trans-Resveratrol and Trans-ε-Viniferin: Evaluation of Their Potential Use for Cardiac Health. Foods, 12(23), 4351. https://doi.org/10.3390/foods12234351