
Hurdle Technology Approach to Control Listeria monocytogenes Using Rhamnolipid Biosurfactant
 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biosurfactant
2.2. Bacterial Strain and Culture Conditions
2.3. Inactivation of L. monocytogenes by Heat and RL in Reduced Moisture Systems
2.4. Growth Inhibitory Activity
2.5. Growth Curves
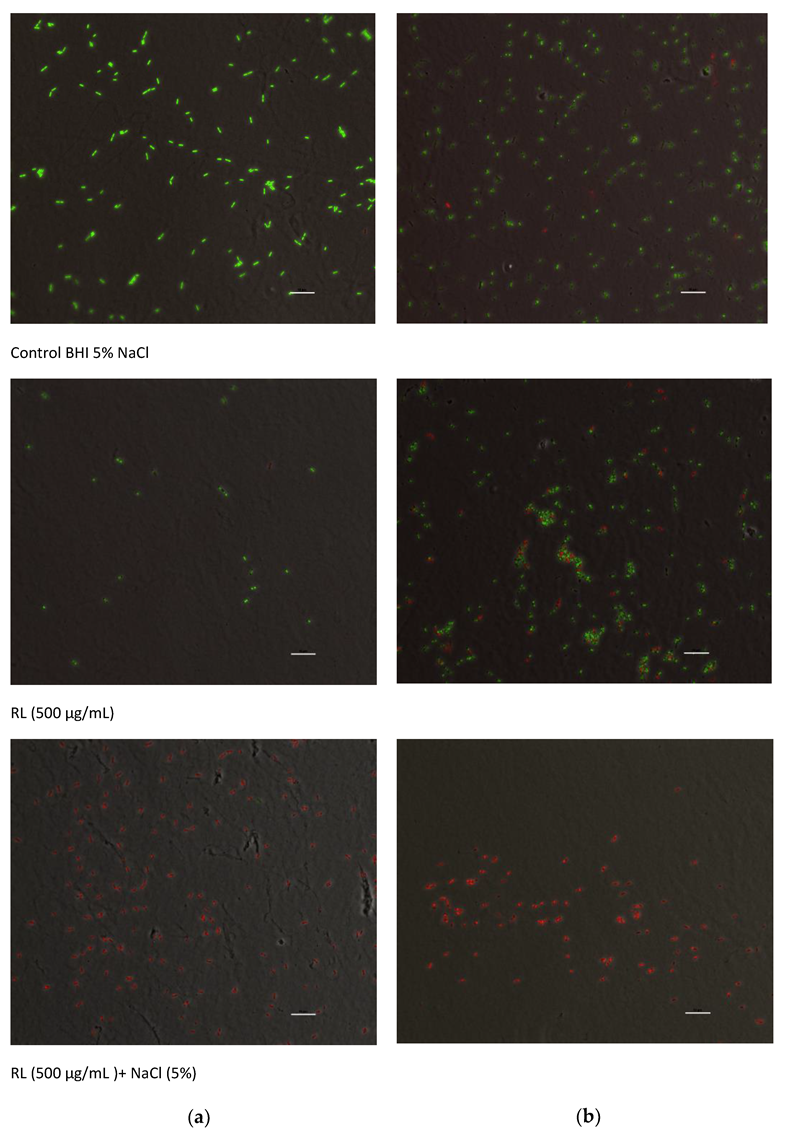
2.6. Microscopy
2.7. CMC Determination
2.8. Statistics
3. Results and Discussion
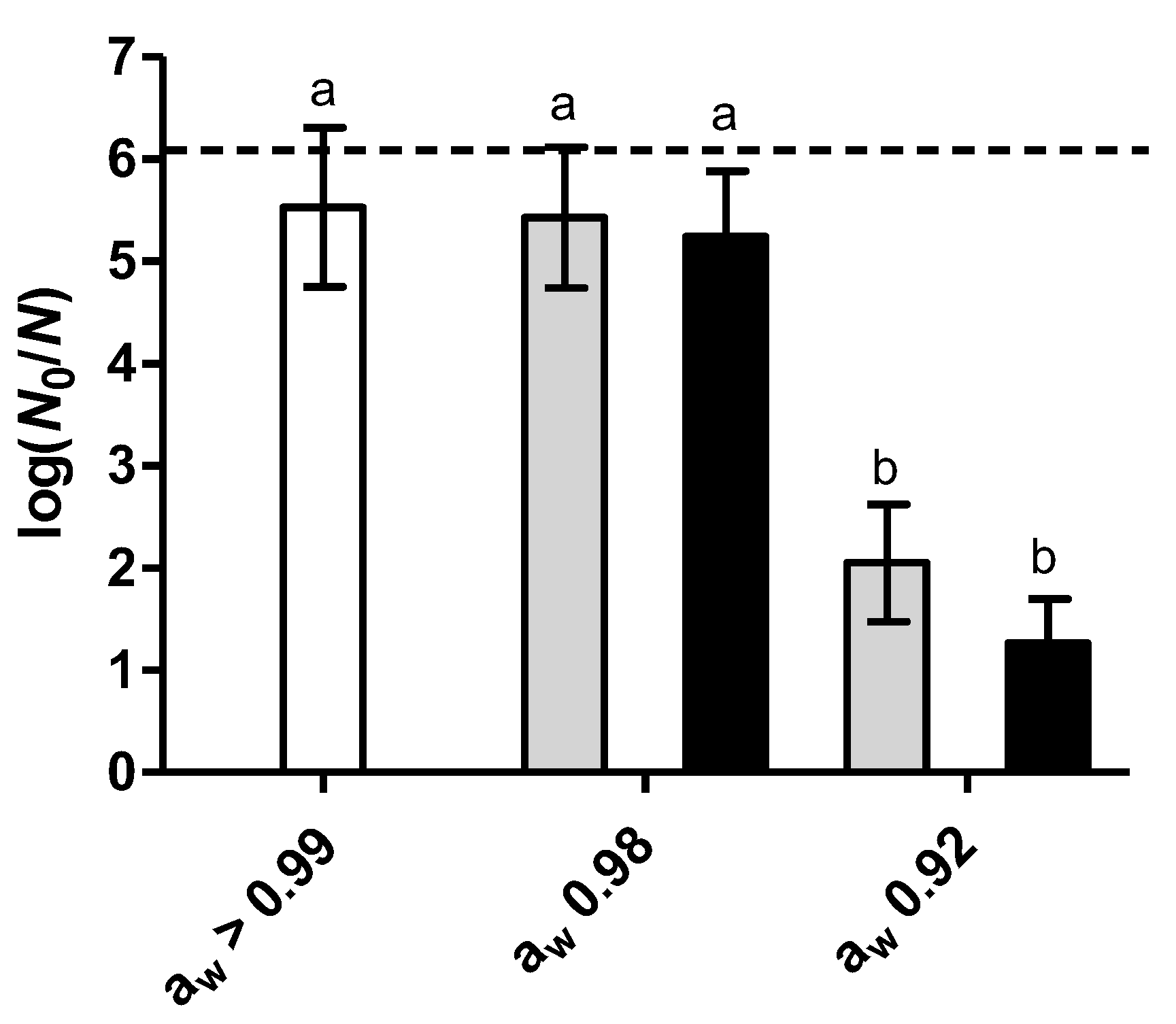
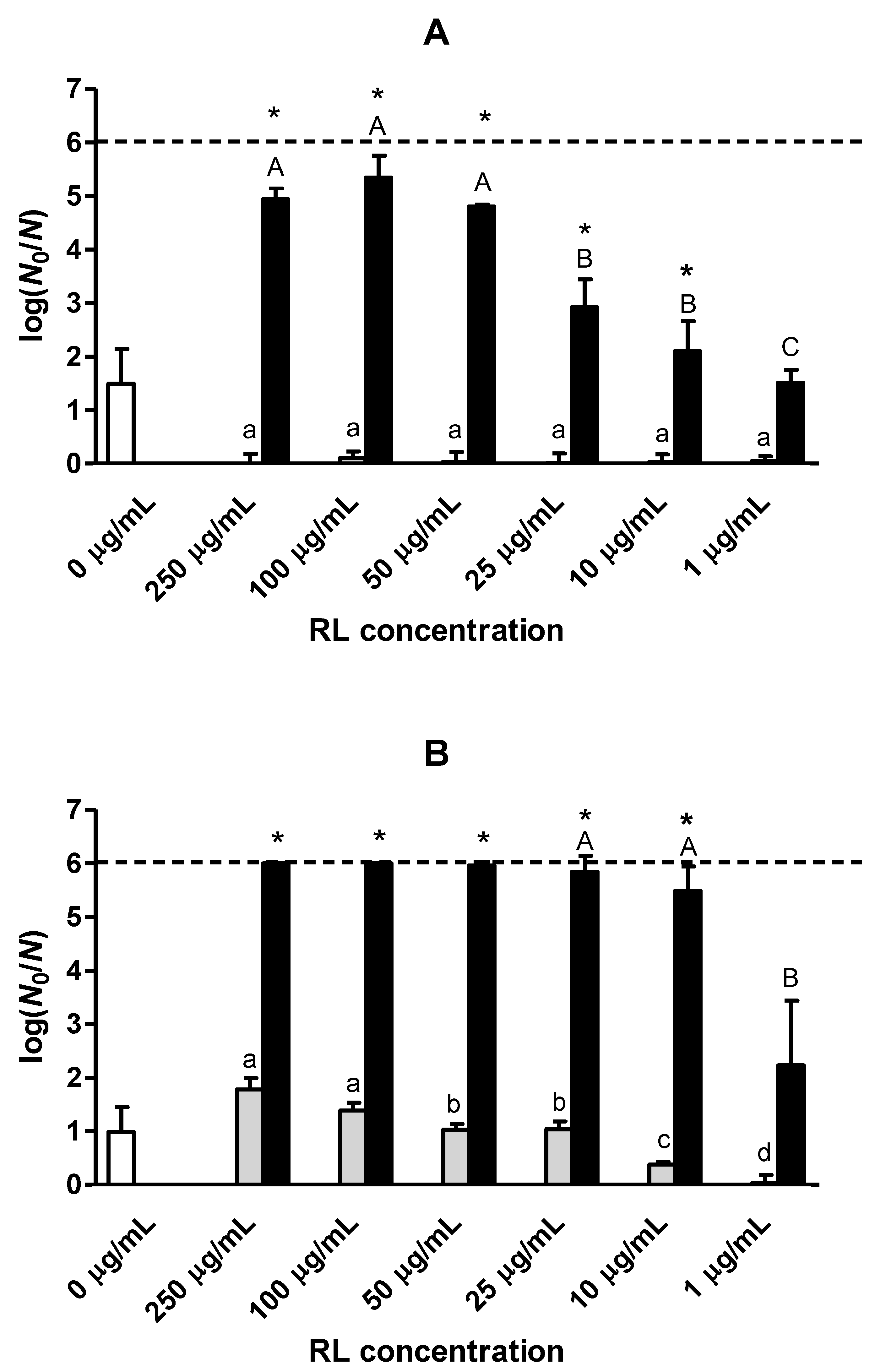
3.1. Effect of RL on Heat Inactivation at Low Water Activity
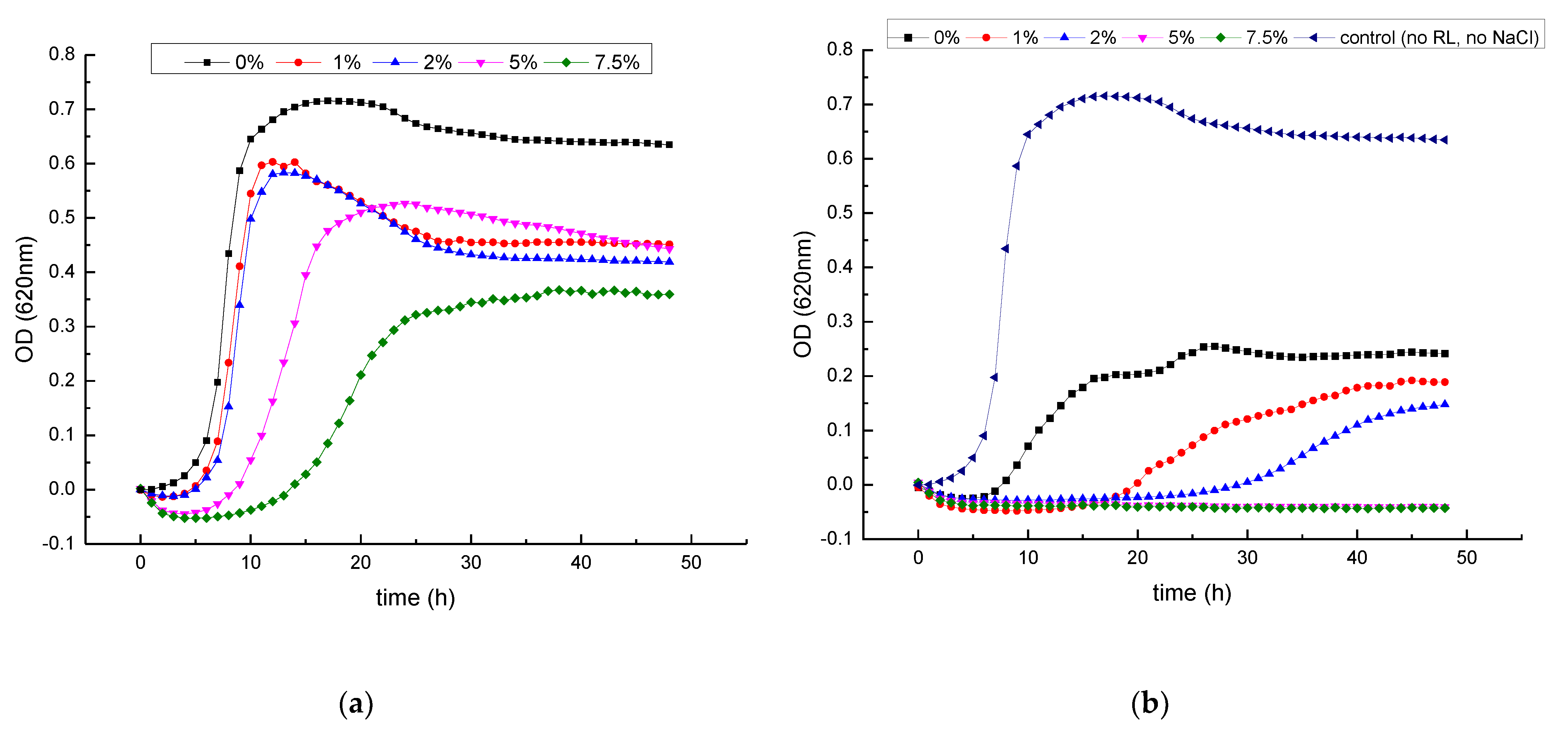
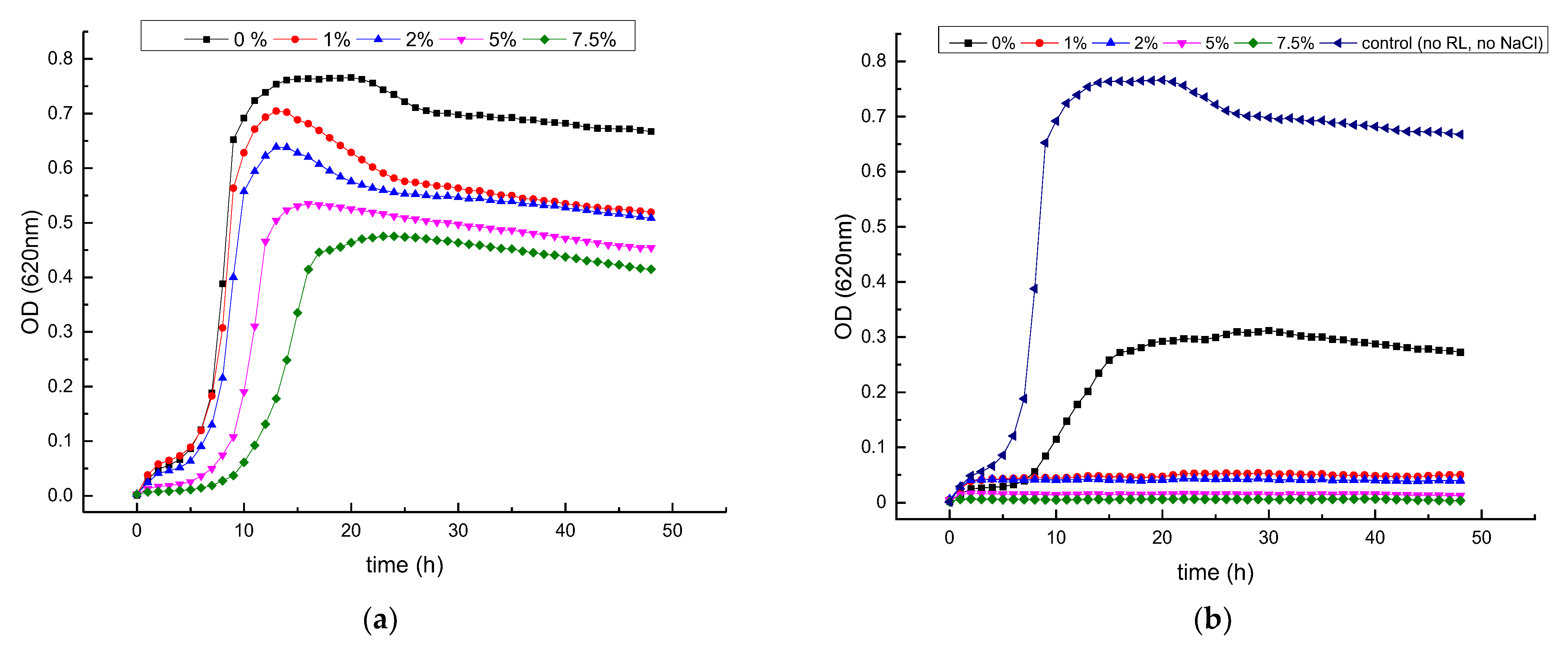
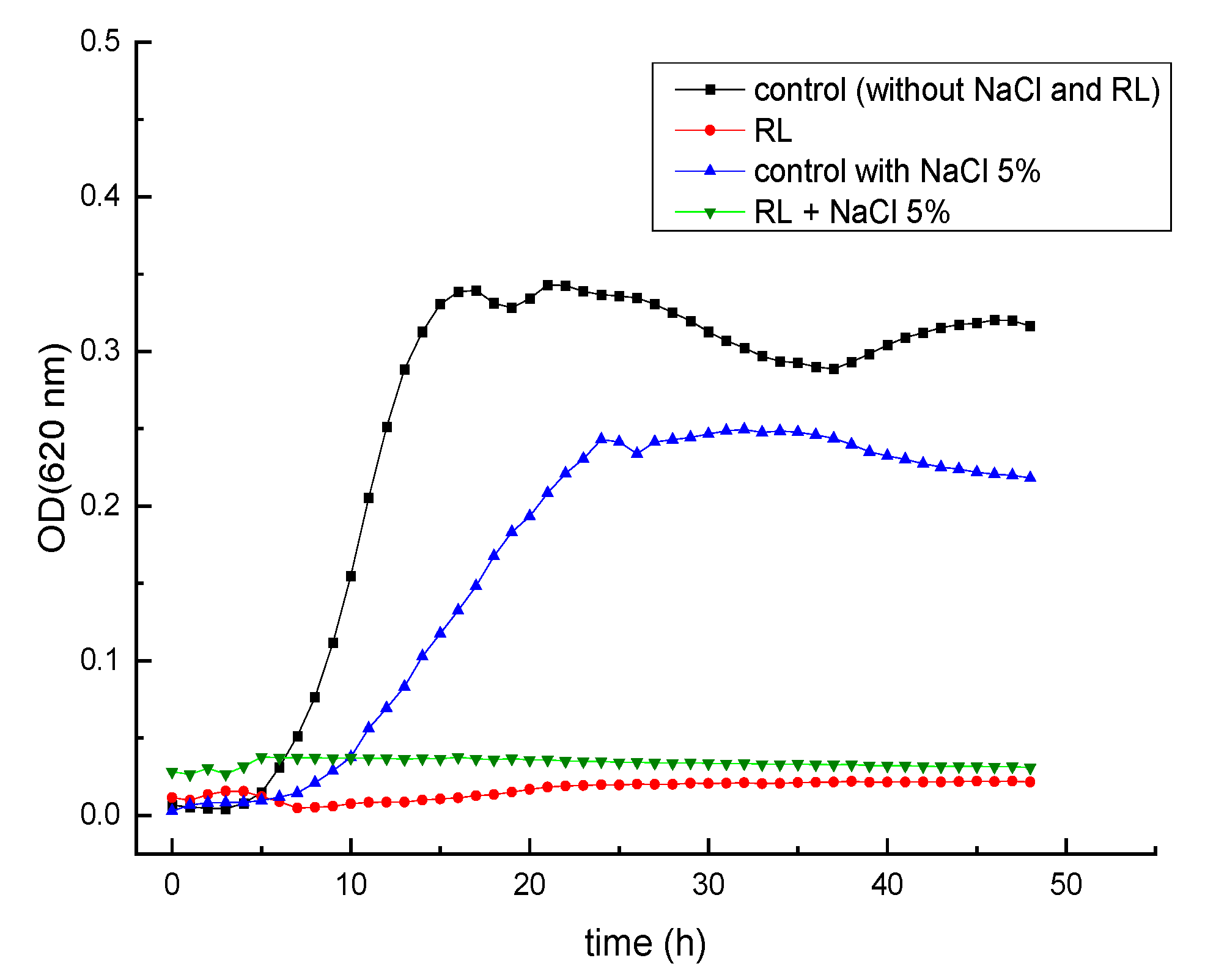
3.2. Growth Inhibitory Activity of RL in Combination with Salts
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barba, F.J.; Koubaa, M.; do Prado-Silva, L.; Orlien, V.; Sant’Ana, A.d.S. Mild processing applied to the inactivation of the main foodborne bacterial pathogens: A review. Trends Food Sci. Technol. 2017, 66, 20–35. [Google Scholar] [CrossRef]
- Ekonomou, S.I.; Boziaris, I.S. Non-thermal methods for ensuring the microbiological quality and safety of seafood. Appl. Sci. 2021, 11, 833. [Google Scholar] [CrossRef]
- De Corato, U. Improving the shelf-life and quality of fresh and minimally-processed fruits and vegetables for a modern food industry: A comprehensive critical review from the traditional technologies into the most promising advancements. Crit. Rev. Food Sci. Nutr. 2020, 60, 940–975. [Google Scholar] [CrossRef]
- Pasha, I.; Saeed, F.; Sultan, M.T.; Khan, M.R.; Rohi, M. Recent developments in minimal processing: A tool to retain nutritional quality of food. Crit. Rev. Food Sci. Nutr. 2014, 54, 340–351. [Google Scholar] [CrossRef]
- Beuchat, L.R.; Komitopoulou, E.; Beckers, H.; Betts, R.P.; Bourdichon, F.; Fanning, S.; Joosten, H.M.; Ter Kuile, B.H. Low–water activity foods: Increased concern as vehicles of foodborne pathogens. J. Food Prot. 2013, 76, 150–172. [Google Scholar] [CrossRef]
- Leistner, L. Basic aspects of food preservation by hurdle technology. Int. J. Food Microbiol. 2000, 55, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Naqash, S.; Naqash, F.; Fayaz, S.; Khan, S.; Dar, B.N.; Makroo, H.A. Application of natural antimicrobial agents in different food packaging systems and their role in shelf-life extension of food: A Review. J. Packag. Technol. Res. 2022, 6, 73–89. [Google Scholar]
- Boskovic, M.; Djordjevic, J.; Ivanovic, J.; Janjic, J.; Zdravkovic, N.; Glisic, M.; Glamoclija, N.; Baltic, B.; Djordjevic, V.; Baltic, M. Inhibition of Salmonella by thyme essential oil and its effect on microbiological and sensory properties of minced pork meat packaged under vacuum and modified atmosphere. Int. J. Food Microbiol. 2017, 258, 58–67. [Google Scholar] [CrossRef]
- García-Díez, J.; Alheiro, J.; Pinto, A.L.; Soares, L.; Falco, V.; Fraqueza, M.J.; Patarata, L. Behaviour of food-borne pathogens on dry cured sausage manufactured with herbs and spices essential oils and their sensorial acceptability. Food Control 2016, 59, 262–270. [Google Scholar] [CrossRef]
- Mulligan, C.N.; Sharma, S.K.; Mudhoo, A.; Makhijani, K. Green chemistry and biosurfactant research. In Biosurfactants Research Trends and Applications; Mulligan, C.N., Sharma, S.K., Mudhoo, A., Eds.; CRC Press: Boca Raton, FL, USA, 2014; pp. 1–30. [Google Scholar]
- Nitschke, M.; Silva, S.S. Recent food applications of microbial surfactants. Crit. Rev. Food Sci. Nutr. 2018, 58, 631–638. [Google Scholar] [CrossRef]
- US Environmental Protection Agency Office of Pesticide Programs. Biopesticides Registration Action Document. Rhamnolipid Biosurfactant (PC Code 110029). 2004. Available online: https://www3.epa.gov/pesticides/chem_search/reg_actions/registration/decision_PC-110029_11-May-04.pdf (accessed on 11 January 2023).
- Hamon, M.; Bierne, H.; Cossart, P. Listeria monocytogenes: A multifaceted model. Nat. Rev. Microbiol. 2006, 4, 423–434. [Google Scholar] [CrossRef] [PubMed]
- Townsend, A.; Strawn, L.K.; Chapman, B.J.; Dunn, L.L. A systematic review of Listeria species and Listeria monocytogenes prevalence, persistence, and diversity throughout the fresh produce supply chain. Foods 2021, 10, 1427. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, R.L.; Gorris, L.G.M.; Hayman, M.M.; Jackson, T.C.; Whiting, R.C. A review of Listeria monocytogenes: An update on outbreaks, virulence, dose-response, ecology, and risk assessments. Food Control 2017, 75, 1–13. [Google Scholar] [CrossRef]
- Tienungoon, S.; Ratkowsky, D.A.; McMeekin, T.A.; Ross, T. Growth limits of Listeria monocytogenes as a function of temperature, pH, NaCl, and lactic acid. Appl. Environ. Microbiol. 2000, 66, 4979–4987. [Google Scholar] [CrossRef] [Green Version]
- Miller, A.J. Combined water activity and solute effects on growth and survival of Listeria monocytogenes Scott A. J. Food Prot. 1992, 55, 414–418. [Google Scholar] [CrossRef]
- Pang, X.; Wong, C.; Chung, H.-J.; Yuk, H.-G. Biofilm formation of Listeria monocytogenes and its resistance to quaternary ammonium compounds in a simulated salmon processing environment. Food Control 2019, 98, 200–208. [Google Scholar] [CrossRef]
- Shamloo, E.; Abdimoghadam, Z.; Nazari, K.; Hosseini, S.M.; Hosseini, H.; Alebouyeh, M. Long-term survival of Listeria monocytogenes in stress conditions: High pH and salt concentrations. J. Res. Med. Dent. Sci. 2018, 6, 96–100. [Google Scholar]
- Araujo, L.V.; Abreu, F.; Lins, U.; Santa Anna, L.M.d.M.; Nitschke, M.; Freire, D.M.G. Rhamnolipid and surfactin inhibit Listeria monocytogenes adhesion. Food Res. Int. 2011, 44, 481–488. [Google Scholar] [CrossRef]
- Magalhães, L.; Nitschke, M. Antimicrobial activity of rhamnolipids against Listeria monocytogenes and their synergistic interaction with nisin. Food Control 2013, 29, 138–142. [Google Scholar] [CrossRef] [Green Version]
- Sotirova, A.V.; Spasova, D.I.; Galabova, D.N.; Karpenko, E.; Shulga, A. Rhamnolipid–biosurfactant permeabilizing effects on Gram-positive and Gram-negative bacterial strains. Curr. Microbiol. 2008, 56, 639–644. [Google Scholar] [CrossRef]
- Smelt, J.P.P.M.; Brul, S. Thermal inactivation of microorganisms. Crit. Rev. Food Sci. Nutr. 2014, 54, 1371–1385. [Google Scholar] [CrossRef] [PubMed]
- Syamaladevi, R.M.; Tang, J.; Villa-Rojas, R.; Sablani, S.; Carter, B.; Campbell, G. Influence of water activity on thermal resistance of microorganisms in low-moisture foods: A review. Compr. Rev. Food. Sci. Food Saf. 2016, 15, 353–370. [Google Scholar] [CrossRef] [PubMed]
- Gayán, E.; Geens, E.; Berdejo, D.; García-Gonzalo, D.; Pagán, R.; Aertsen, A.; Michiels, C.W. Combination of mild heat and plant essential oil constituents to inactivate resistant variants of Escherichia coli in buffer and in coconut water. Food Microbiol. 2020, 87, 103388. [Google Scholar]
- Espina, L.; Somolinos, M.; Ouazzou, A.A.; Condón, S.; García-Gonzalo, D.; Pagán, R. Inactivation of Escherichia coli O157:H7 in fruit juices by combined treatments of citrus fruit essential oils and heat. Int. J. Food Microbiol. 2012, 159, 9–16. [Google Scholar] [PubMed]
- Arioli, S.; Montanari, C.; Magnani, M.; Tabanelli, G.; Patrignani, F.; Lanciotti, R.; Mora, D.; Gardini, F. Modelling of Listeria monocytogenes Scott A after a mild heat treatment in the presence of thymol and carvacrol: Effects on culturability and viability. J. Food Eng. 2019, 240, 73–82. [Google Scholar] [CrossRef]
- Fugett, E.; Fortes, E.; Nnoka, C.; Wiedmann, M. International Life Sciences Institute North America Listeria monocytogenes strain collection: Development of standard Listeria monocytogenes strain sets for research and validation studies. J. Food Prot. 2006, 69, 2929–2938. [Google Scholar] [CrossRef] [PubMed]
- Sieuwerts, S.; de Bok, F.A.M.; Mols, E.; de Vos, W.M.; van Hylckama Vlieg, J.E.T. A simple and fast method for determining colony forming units. Lett. Appl. Microbiol. 2008, 47, 275–278. [Google Scholar] [CrossRef]
- Feyaerts, J.; Rogiers, G.; Corthouts, J.; Michiels, C.W. Thiol-reactive natural antimicrobials and high-pressure treatment synergistically enhance bacterial inactivation. Innov. Food Sci. Emerg. Technol. 2015, 27, 26–34. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 9th ed.; Approved Standard; CLSI Document M07-A9; Clinical and Laboratory Standards Institute (CLSI): Wayne, PA, USA, 2012. [Google Scholar]
- Rodríguez-Melcón, C.; Alonso-Calleja, C.; García-Fernández, C.; Carballo, J.; Capita, R. Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC) for twelve antimicrobials (biocides and antibiotics) in eight strains of Listeria monocytogenes. Biology 2022, 11, 46. [Google Scholar] [CrossRef]
- Sun, L.; Rogiers, G.; Courtin, P.; Chapot-Chartier, M.P.; Bierne, H.; Michiels, C.W. AsnB mediates amidation of meso-diaminopimelic acid residues in the peptidoglycan of Listeria monocytogenes and affects bacterial surface properties and host cell invasion. Front. Microbiol. 2021, 12, 760253. [Google Scholar] [CrossRef]
- LIVE/DEAD BacLight Bacterial Viability Kit Protocol. Available online: https://www.thermofisher.com/br/en/home/references/protocols/cell-and-tissue-analysis/protocols/live-dead-baclight-bacterial-viability-protocol.html (accessed on 9 January 2023).
- Fernández, A.; López, M.; Bernardo, A.; Condón, S.; Raso, J. Modelling thermal inactivation of Listeria monocytogenes in sucrose solutions of various water activities. Food Microbiol. 2007, 24, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Cava-Roda, R.M.; Taboada, A.; Palop, A.; López-Gómez, A.; Marin-Iniesta, F. Heat resistance of Listeria monocytogenes in semi-skim milk supplemented with vanillin. Int. J. Food Microbiol. 2012, 157, 314–318. [Google Scholar] [CrossRef] [PubMed]
- Cherrat, L.; Espina, L.; Bakkali, M.; García-Gonzalo, D.; Pagán, R.; Laglaoui, A. Chemical composition and antioxidant properties of Laurus nobilis L. and Myrtus communis L. essential oils from Morocco and evaluation of their antimicrobial activity acting alone or in combined processes for food preservation. J. Sci. Food Agric. 2014, 94, 1197–1204. [Google Scholar] [CrossRef] [PubMed]
- Maté, J.; Periago, P.M.; Palop, A. When nanoemulsified, d-limonene reduces Listeria monocytogenes heat resistance about one hundred times. Food Control 2016, 59, 824–828. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Guan, X.; Lin, B.; Li, R.; Wang, S. Oregano oil, epsilon-polylysine and citric acid assisted inactivation of Salmonella in two kinds of tahini during thermal treatment and storage. Foods 2021, 10, 1272. [Google Scholar] [CrossRef]
- Chen, W.; Golden, D.A.; Critzer, F.J.; Davidson, P.M. Antimicrobial activity of cinnamaldehyde, carvacrol, and lauric arginate against Salmonella Tennessee in a glycerol-sucrose model and peanut paste at different fat concentrations. J. Food Prot. 2015, 78, 1488–1495. [Google Scholar]
- Ortiz, A.; Teruel, J.A.; Espuny, M.J.; Marqués, A.; Manresa, Á.; Aranda, F.J. Effects of dirhamnolipid on the structural properties of phosphatidylcholine membranes. Int. J. Pharma. 2006, 325, 99–107. [Google Scholar] [CrossRef]
- Gaur, V.K.; Tripathi, V.; Gupta, P.; Dhiman, N.; Regar, R.K.; Gautam, K.; Srivastava, J.K.; Patnaik, S.; Patel, D.K.; Manickam, N. Rhamnolipids from Planococcus spp. and their mechanism of action against pathogenic bacteria. Bioresour. Technol. 2020, 307, 123206. [Google Scholar] [CrossRef]
- Yutaka, I.; Yasuo, G.; Hitoshi, N.; Muneo, Y.; Hisae, N.; Toshio, K. The pH-Sensitive conversion of molecular aggregates of rhamnolipid biosurfactant. Chem. Lett. 1987, 16, 763–766. [Google Scholar]
- Zhou, C.; Wang, F.; Chen, H.; Li, M.; Qiao, F.; Liu, Z.; Hou, Y.; Wu, C.; Fan, Y.; Liu, L.; et al. Selective antimicrobial activities and action mechanism of micelles self-assembled by cationic oligomeric surfactants. ACS Appl. Mat. Interfaces 2016, 8, 4242–4249. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.I.; Gudiña, E.J.; Teixeira, J.A.; Rodrigues, L.R. Sodium chloride effect on the aggregation behaviour of rhamnolipids and their antifungal activity. Sci. Rep. 2017, 7, 12907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, S.S.; Carvalho, J.W.P.; Aires, C.P.; Nitschke, M. Disruption of Staphylococcus aureus biofilms using rhamnolipid biosurfactants. J. Dairy Sci. 2017, 100, 7864–7873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, H.; Yang, L.; Yang, X.; Zeng, G.; Liu, Z.; Liu, Y.; Yuan, X. Aggregation of low-concentration dirhamnolipid biosurfactant in electrolyte solution. RSC Adv. 2015, 5, 88578–88582. [Google Scholar] [CrossRef] [Green Version]
- Helvacı, Ş.Ş.; Peker, S.; Özdemir, G. Effect of electrolytes on the surface behavior of rhamnolipids R1 and R2. Colloids Surf. B Biointerfaces 2004, 35, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Demissie, H.; Duraisamy, R. Effects of electrolytes on the surface and micellar characteristics of sodium dodecyl sulphate surfactant solution. J. Sci. Innov. Res. 2016, 5, 208–214. [Google Scholar] [CrossRef]
- Cepanec, K.; Vugrinec, S.; Cvetković, T.; Ranilović, J. Potassium chloride-based salt substitutes: A critical review with a focus on the patent literature. Compr. Rev. Food. Sci. Food Saf. 2017, 16, 881–894. [Google Scholar] [CrossRef] [Green Version]
- Lebrón-Paler, A.; Pemberton, J.E.; Becker, B.A.; Otto, W.H.; Larive, C.K.; Maier, R.M. Determination of the acid dissociation constant of the biosurfactant monorhamnolipid in aqueous solution by potentiometric and spectroscopic methods. Anal. Chem. 2006, 78, 7649–7658. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, M.A.; Shadizadeh, S.R.; Salari, Z. Dependency of critical micellization concentration of an anionic surfactant on temperature and potassium chloride salt. Pet. Sci. Technol. 2014, 32, 1913–1920. [Google Scholar] [CrossRef]
- Sood, A.K.; Aggarwal, M. Evaluation of micellar properties of sodium dodecylbenzene sulphonate in the presence of some salts. J. Chem. Sci. 2018, 130, 39. [Google Scholar] [CrossRef] [Green Version]
- Abbasi, H.; Noghabi, K.A.; Hamedi, M.M.; Zahiri, H.S.; Moosavi-Movahedi, A.A.; Amanlou, M.; Teruel, J.A.; Ortiz, A. Physicochemical characterization of a monorhamnolipid secreted by Pseudomonas aeruginosa MA01 in aqueous media. An experimental and molecular dynamics study. Colloids Surf. B Biointerfaces 2013, 101, 256–265. [Google Scholar] [CrossRef]
- Ferreira, J.F.; Vieira, E.A.; Nitschke, M. The antibacterial activity of rhamnolipid biosurfactant is pH dependent. Food Res. Int. 2019, 116, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Raza, Z.A.; Khalid, Z.M.; Khan, M.S.; Banat, I.M.; Rehman, A.; Naeem, A.; Saddique, M.T. Surface properties and sub-surface aggregate assimilation of rhamnolipid surfactants in different aqueous systems. Biotechnol. Lett. 2010, 32, 811–816. [Google Scholar] [CrossRef] [PubMed]
- Ciesla, J.; Koczanska, M.; Bieganowski, A. An interaction of rhamnolipids with Cu2+ ions. Molecules 2018, 23, 488. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Salt Concentration | 0.0% | 1.0% | 2.0% | 5.0% | 7.5% |
|---|---|---|---|---|---|
| NaCl | >2500 (-) | 2500 (-) | 312 (-) | 78 (78) | 39 (39) |
| KCl | >2500 (-) | 312 (625) | 156 (156) | 78 (78) | 39 (39) |
| %NaCl | 0.0% | 1.0% (171 mM) | 2.0% (342 mM) | 5.0% (855 mM) |
| pH 5.0 | 19.6 | 11.5 | 10.4 | 11.9 |
| pH 6.0 | 93.2 | 32.0 | 17.4 | 14.2 |
| pH 7.0 | 94.2 | 35.8 | 26.3 | 16.0 |
| pH 8.0 | 121.4 | 39.1 | 25.6 | 15.5 |
| % KCl | 0.0% | 1.0% (134 mM) | 2.0% (268 mM) | 5.0% (670 mM) |
| pH 7.0 | 94.2 | 46.9 | 38.3 | 21.7 |
| % KCl | 1.3% (171mM) | 2.5% (342 mM) | 6.4% (855 mM) | |
| pH 7.0 | 25.5 | 19.5 | 13.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lenaerts, L.; Passos, T.F.; Gayán, E.; Michiels, C.W.; Nitschke, M. Hurdle Technology Approach to Control Listeria monocytogenes Using Rhamnolipid Biosurfactant. Foods 2023, 12, 570. https://doi.org/10.3390/foods12030570
Lenaerts L, Passos TF, Gayán E, Michiels CW, Nitschke M. Hurdle Technology Approach to Control Listeria monocytogenes Using Rhamnolipid Biosurfactant. Foods. 2023; 12(3):570. https://doi.org/10.3390/foods12030570
Chicago/Turabian StyleLenaerts, Lowieze, Tathiane Ferroni Passos, Elisa Gayán, Chris W. Michiels, and Marcia Nitschke. 2023. "Hurdle Technology Approach to Control Listeria monocytogenes Using Rhamnolipid Biosurfactant" Foods 12, no. 3: 570. https://doi.org/10.3390/foods12030570
APA StyleLenaerts, L., Passos, T. F., Gayán, E., Michiels, C. W., & Nitschke, M. (2023). Hurdle Technology Approach to Control Listeria monocytogenes Using Rhamnolipid Biosurfactant. Foods, 12(3), 570. https://doi.org/10.3390/foods12030570