Three Molecular Modification Strategies to Improve the Thermostability of Xylanase XynA from Streptomyces rameus L2001

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Gene, Strains, Media and Substrates
2.2. Construction of Mutant Xylanase
2.3. Expression and Purification of Xylanases
2.4. Xylanase Activity Assay
2.5. Biochemical Characterization of Xylanase
2.6. Hydrolytic Characteristics of XynA Mutants
2.7. Sequence and Structural Analysis
2.8. Statistical Analysis
3. Results and Discussion
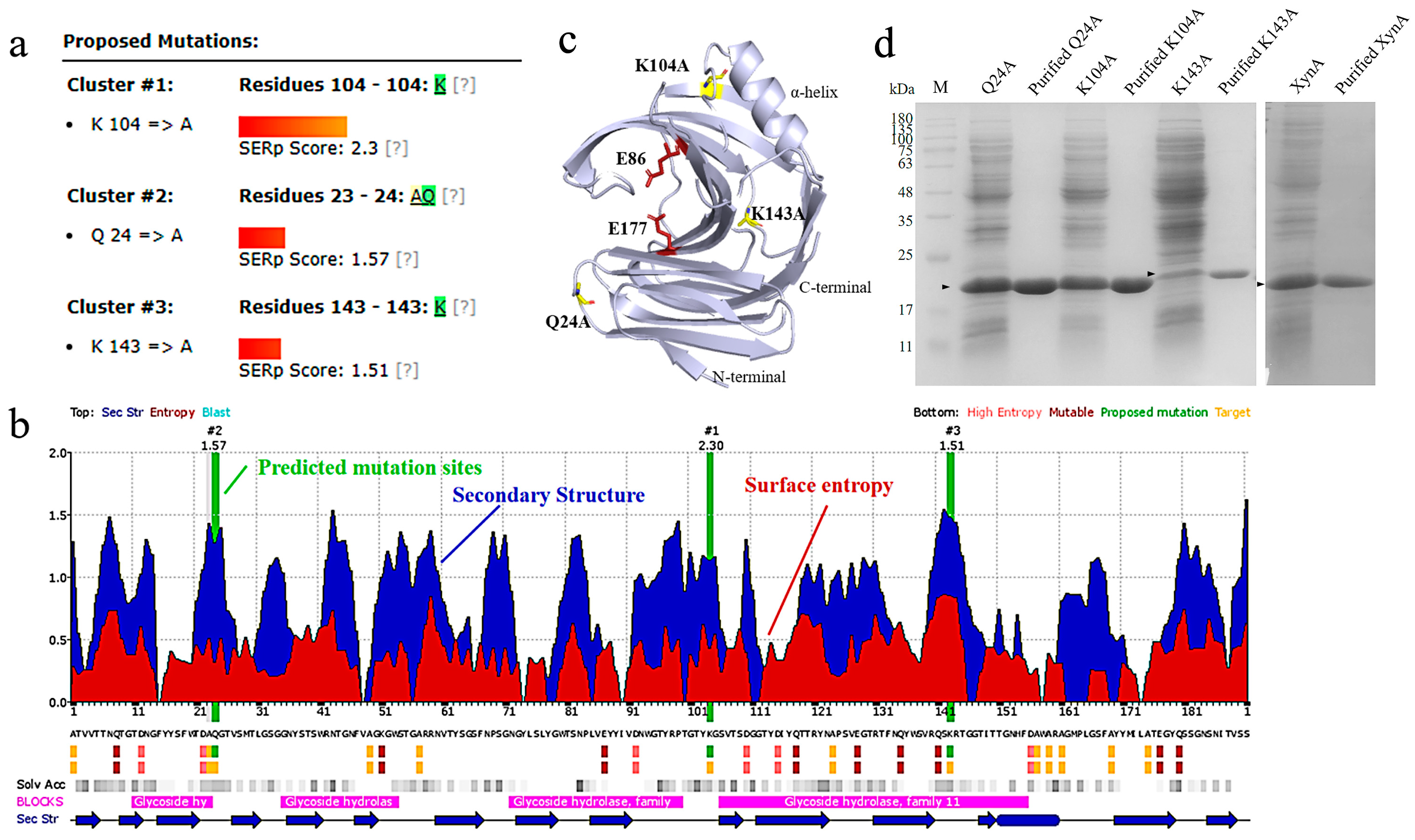
3.1. Modification of Thermostability Based on the Surface Entropy
3.1.1. Production of Surface Entropy Mutants
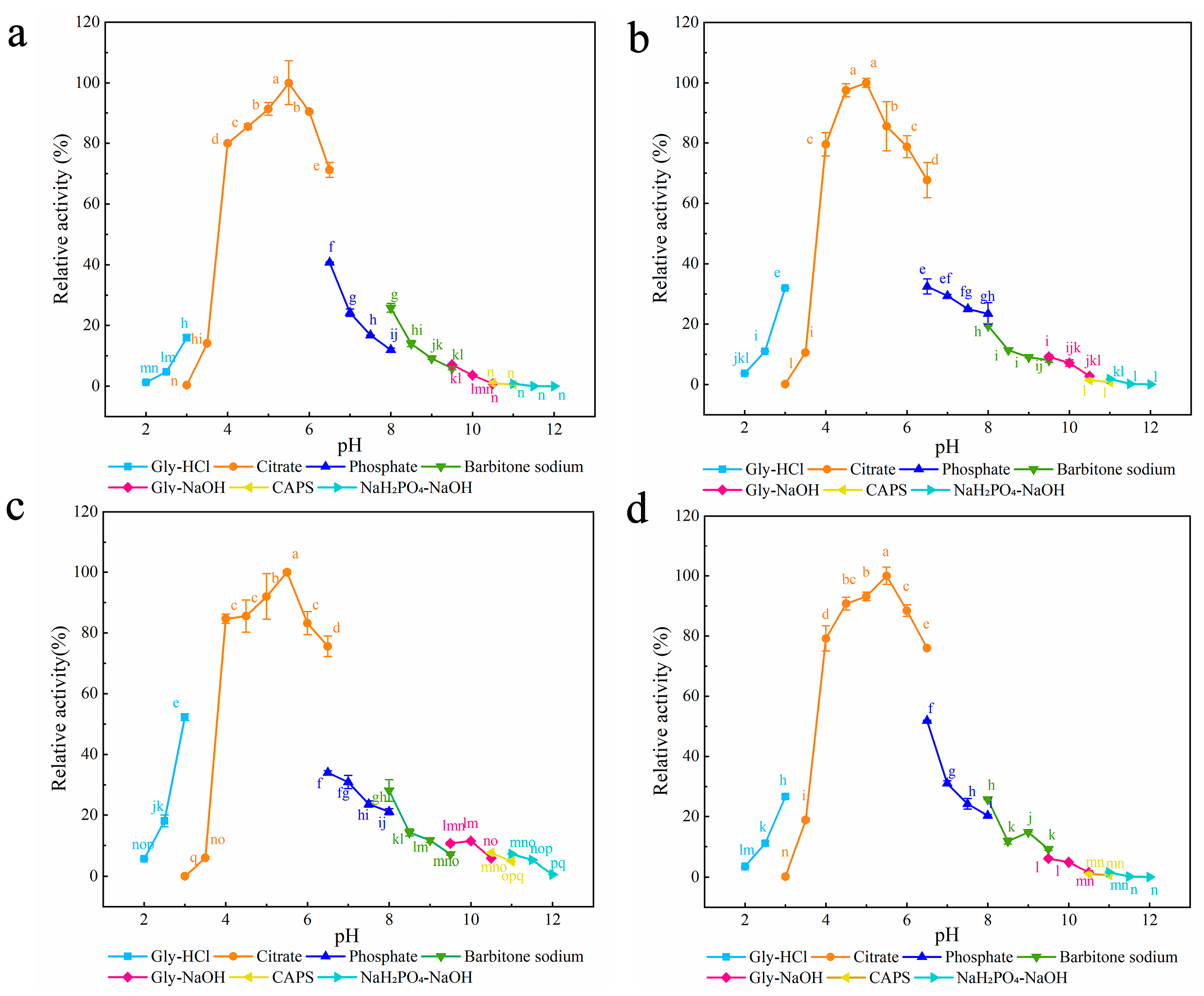
3.1.2. Characterization of Surface Entropy Mutants
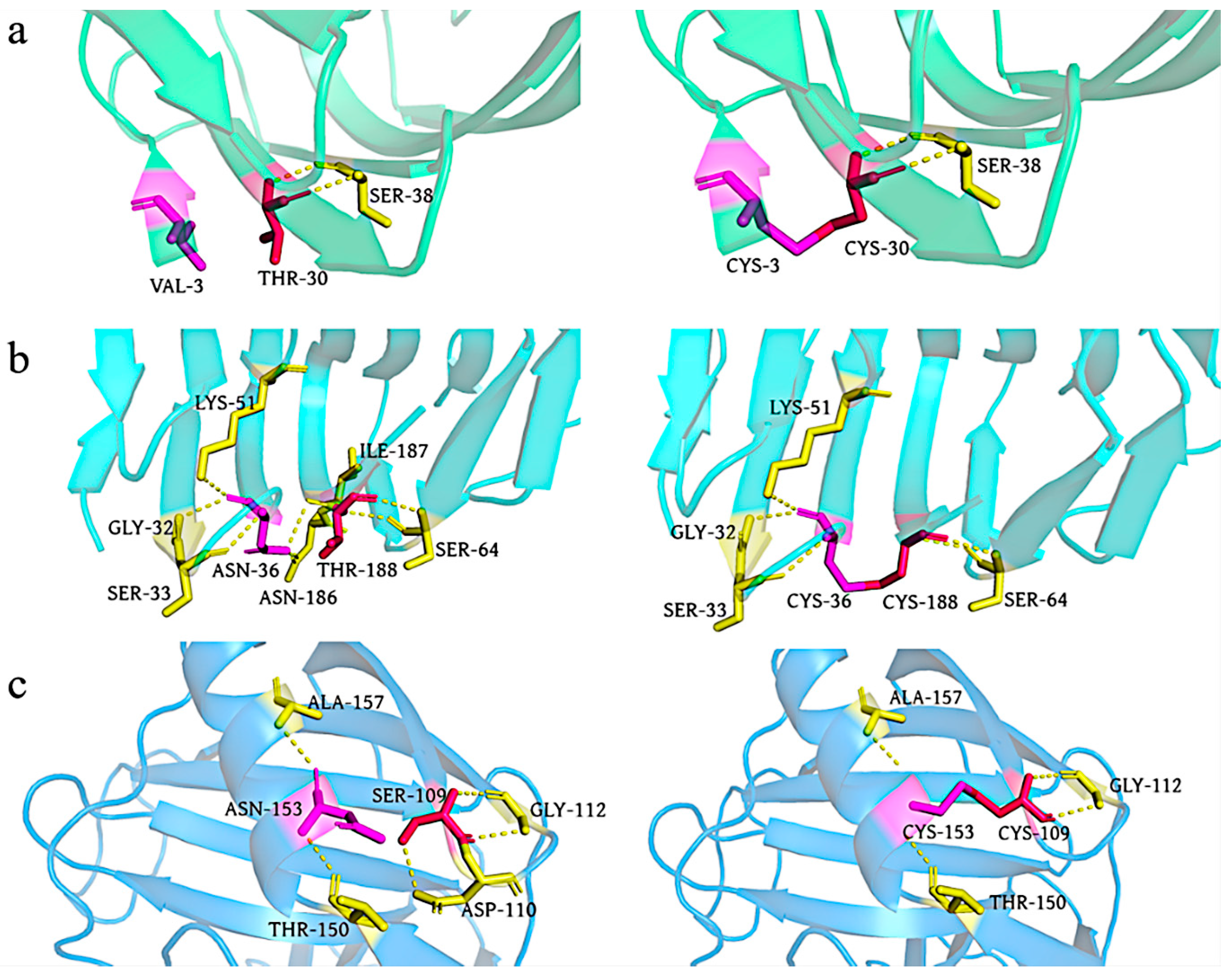
3.2. Thermostability of Disulfide Bonds Mutations of XynA
3.2.1. Construction of Disulfide Bonds Mutants
3.2.2. Characterization of Mutants with Disulfide Bonds
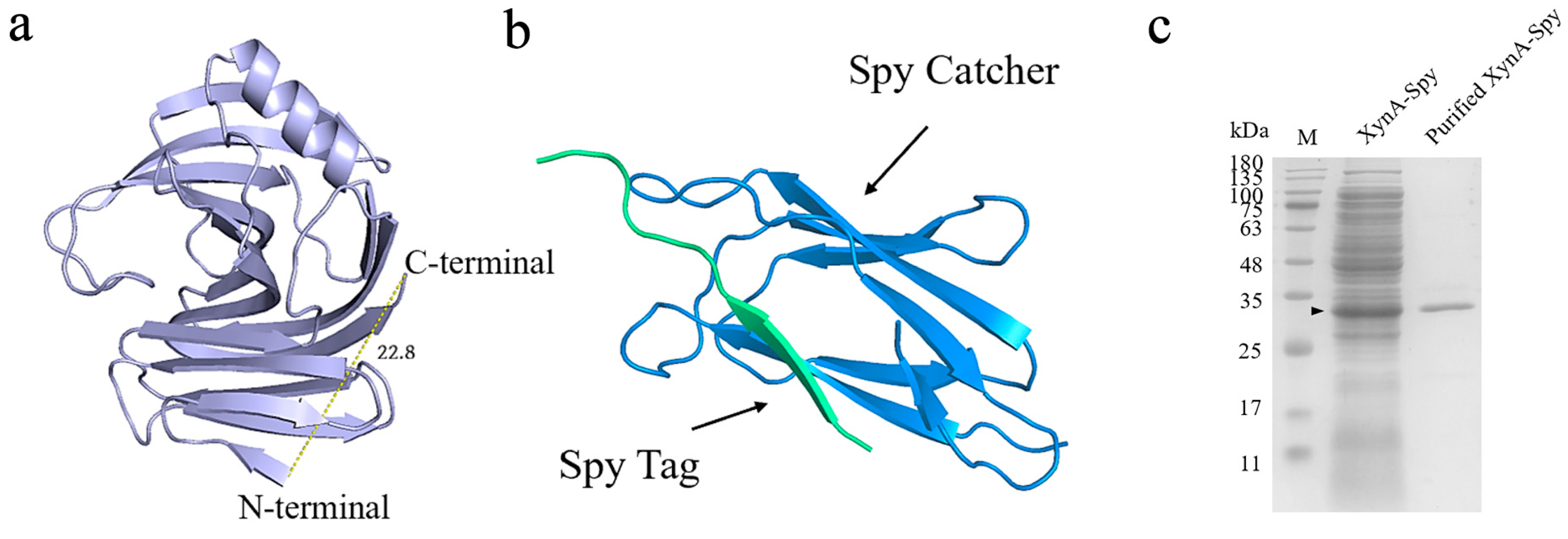
3.3. Thermostability of XynA with Heteropeptide-Mediated Cyclization
3.3.1. Construction of XynA Cyclization Mutants
3.3.2. Characterization of Cyclization Mutants
3.4. Comparison of the Three Strategies for Thermostability Modification
3.5. Catalytic and Hydrolytic Properties of XynA and the Mutants
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Glenwright, A.J.; Pothula, K.R.; Bhamidimarri, S.P.; Chorev, D.S.; Baslé, A.; Firbank, S.J.; Zheng, H.; Robinson, C.v.; Winterhalter, M.; Kleinekathöfer, U.; et al. Structural basis for nutrient acquisition by dominant members of the human gut microbiota. Nature 2017, 541, 407–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desai, M.S.; Seekatz, A.M.; Koropatkin, N.M.; Kamada, N.; Hickey, C.A.; Wolter, M.; Pudlo, N.A.; Kitamoto, S.; Terrapon, N.; Muller, A.; et al. A dietary fiber-deprived gut microbiota degrades the colonic mucus barrier and enhances pathogen susceptibility. Cell 2016, 167, 1339–1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Wu, M.; Niu, J.; Lin, M.; Zhu, H.; Wang, K.; Li, X.; Sun, B. Characteristics and correlation of the microbial communities and flavor compounds during the first three rounds of fermentation in Chinese sauce-flavor Baijiu. Foods 2023, 12, 207. [Google Scholar] [CrossRef]
- Xu, Y.; Huang, H.; Lu, H.; Wu, M.; Lin, M.; Zhang, C.; Zhao, Z.; Li, W.; Zhang, C.; Li, X.; et al. Characterization of an Aspergillus niger for efficient fatty acid ethyl ester synthesis in aqueous phase and the molecular mechanism. Front. Microbiol. 2022, 12, 820380. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhu, Y.; Li, X.; Sun, B. Dynamic balancing of intestinal short-chain fatty acids: The crucial role of bacterial metabolism. Trends Food Sci. Technol. 2020, 100, 118–130. [Google Scholar] [CrossRef]
- Li, Q.; Sun, B.; Li, X.; Xiong, K.; Xu, Y.; Yang, R.; Hou, J.; Teng, C. Improvement of the catalytic characteristics of a salt-tolerant GH10 xylanase from Streptomyce rochei L10904. Int. J. Biol. Macromol. 2018, 107, 1447–1455. [Google Scholar] [CrossRef]
- Abdulaziz Abbod Abdo, A.; Zhang, C.; Lin, Y.; Liang, X.; Kaddour, B.; Wu, Q.; Li, X.; Fan, G.; Yang, R.; Teng, C.; et al. Xylo-oligosaccharides ameliorate high cholesterol diet induced hypercholesterolemia and modulate sterol excretion and gut microbiota in hamsters. J. Funct Foods 2021, 77, 104334. [Google Scholar] [CrossRef]
- Ullah, S.F.; Souza, A.A.; de Freitas, S.M.; Noronha, E.F. Characterisation of biomass degrading xylanolytic enzymes of Penicillium chrysogenum produced using sugarcane bagasse. Process Biochem. 2022, 112, 62–70. [Google Scholar] [CrossRef]
- Poletto, P.; Pereira, G.N.; Monteiro, C.; Pereira, M.A.F.; Bordignon, S.E.; de Oliveira, D. Xylooligosaccharides: Transforming the lignocellulosic biomasses into valuable 5-carbon sugar prebiotics. Process Biochem. 2020, 91, 352–363. [Google Scholar] [CrossRef]
- Zouari Ayadi, D.; Hmida Sayari, A.; ben Hlima, H.; ben Mabrouk, S.; Mezghani, M.; Bejar, S. Improvement of Trichoderma reesei xylanase II thermal stability by serine to threonine surface mutations. Int. J. Biol. Macromol. 2015, 72, 163–170. [Google Scholar] [CrossRef]
- Zhuo, R.; Yu, H.; Qin, X.; Ni, H.; Jiang, Z.; Ma, F.; Zhang, X. Heterologous expression and characterization of a xylanase and xylosidase from white rot fungi and their application in synergistic hydrolysis of lignocellulose. Chemosphere 2018, 212, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; He, Y.; Yu, H.; Dong, Z. Seven N-terminal residues of a thermophilic xylanase are sufficient to confer hyperthermostability on its mesophilic counterpart. PLoS One 2014, 9, e87632. [Google Scholar] [CrossRef]
- Jeong, M.Y.; Kim, S.; Yun, C.W.; Choi, Y.J.; Cho, S.G. Engineering a de novo internal disulfide bridge to improve the thermal stability of xylanase from Bacillus stearothermophilus No. 236. J. Biotechnol. 2007, 127, 300–309. [Google Scholar] [CrossRef] [PubMed]
- Min, K.; Kim, H.; Park, H.J.; Lee, S.; Jung, Y.J.; Yoon, J.H.; Lee, J.S.; Park, K.; Yoo, Y.J.; Joo, J.C. Improving the catalytic performance of xylanase from Bacillus circulans through structure-based rational design. Bioresour. Technol. 2021, 340, 125737. [Google Scholar] [CrossRef]
- Liu, L.; Wang, B.; Chen, H.; Wang, S.; Wang, M.; Zhang, S.; Song, A.; Shen, J.; Wu, K.; Jia, X. Rational pH-engineering of the thermostable xylanase based on computational model. Process Biochem. 2009, 44, 912–915. [Google Scholar] [CrossRef]
- Paës, G.; Berrin, J.G.; Beaugrand, J. GH11 xylanases: Structure/function/properties relationships and applications. Biotechnol. Adv. 2012, 30, 564–592. [Google Scholar] [CrossRef]
- Perl, D.; Mueller, U.; Heinemann, U.; Schmid, F.X. Two exposed amino acid residues confer thermostability on a cold shock protein. Nat. Struct. Biol. 2000, 7, 380–383. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.; Markus, W. Engineering proteins for thermostability: The use of sequence alignments versus rational design and directed evolution sequence. Curr. Opin. Biotechnol. 2001, 12, 371–375. [Google Scholar] [CrossRef]
- Arnold, F.H. Combinatorial and computational challenges for biocatalyst design. Nature 2001, 409, 253–257. [Google Scholar] [CrossRef]
- Wang, L.; Cao, K.; Pedroso, M.M.; Wu, B.; Gao, Z.; He, B.; Schenk, G. Sequence- and structure-guided improvement of the catalytic performance of a GH11 family xylanase from Bacillus subtilis. J. Biol. Chem. 2021, 297, 101262. [Google Scholar] [CrossRef]
- Steipe, B. Evolutionary approaches to protein engineering. Comb. Chem. Biol. 1999, 243, 55–86. [Google Scholar] [CrossRef]
- Kazlauskas, R. Engineering more stable proteins. Chem. Soc. Rev. 2018, 47, 9026–9045. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, M.; Signor, G.; Matthews, B.W. Substantial increase of protein stability by multiple disulphide bonds. Nature 1989, 342, 291–293. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Fu, Z.; Huang, H.; Zhang, H.; Yao, B.; Xiong, H.; Turunen, O. Improved thermal performance of Thermomyces lanuginosus GH11 xylanase by engineering of an N-terminal disulfide bridge. Bioresour Technol. 2012, 112, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Han, N.; Ma, Y.; Mu, Y.; Tang, X.; Li, J.; Huang, Z. Enhancing thermal tolerance of a fungal GH11 xylanase guided by b-factor analysis and multiple sequence alignment. Enzym. Microb. Technol. 2019, 131, 109422. [Google Scholar] [CrossRef]
- Vogt, G.; Woell, S.; Argos, P. Protein thermal stability, hydrogen bonds, and ion pairs. J. Mol. Biol. 1997, 269, 631–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieira, D.S.; Degrève, L. An insight into the thermostability of a pair of xylanases: The role of hydrogen bonds. Mol. Phys. 2009, 107, 59–69. [Google Scholar] [CrossRef]
- Abdul Wahab, M.K.H.B.; bin Jonet, M.A.; Illias, R.M. Thermostability enhancement of xylanase Aspergillus fumigatus RT-1. J. Mol. Catal. B Enzym. 2016, 134, 154–163. [Google Scholar] [CrossRef]
- Kim, T.; Joo, J.C.; Yoo, Y.J. Hydrophobic interaction network analysis for thermostabilization of a mesophilic xylanase. J. Biotechnol. 2012, 161, 49–59. [Google Scholar] [CrossRef]
- Miyazaki, K.; Takenouchi, M.; Kondo, H.; Noro, N.; Suzuki, M.; Tsuda, S. Thermal stabilization of Bacillus subtilis family-11 xylanase by directed evolution. J. Biol. Chem. 2006, 281, 10236–10242. [Google Scholar] [CrossRef] [Green Version]
- Xue, H.; Zhou, J.; You, C.; Huang, Q.; Lu, H. Amino acid substitutions in the N-terminus, cord and α-helix domains improved the thermostability of a family 11 xylanase XynR8. J. Ind. Microbiol. Biotechnol. 2012, 39, 1279–1288. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Tsai, C.J.; Nussinov, R. Factors enhancing protein thermostability. Protein Eng. 2000, 13, 179–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Ma, B.Y.; Tsai, C.J.; Nussinov, R. Electrostatic strengths of salt bridges in thermophilic and mesophilic glutamate dehydrogenase monomers. Proteins-Struct. Funct. Bioinform. 2000, 38, 368–383. [Google Scholar] [CrossRef]
- Sutthibutpong, T.; Rattanarojpong, T.; Khunrae, P. Effects of helix and fingertip mutations on the thermostability of Xyn11A investigated by molecular dynamics simulations and enzyme activity assays. J. Biomol. Struct. Dyn. 2018, 36, 3978–3992. [Google Scholar] [CrossRef]
- Paës, G.; O’Donohue, M.J. Engineering increased thermostability in the thermostable GH-11 xylanase from Xylanilyticus. J. Biotechnol. 2006, 125, 338–350. [Google Scholar] [CrossRef] [PubMed]
- Derewenda, Z.S. It’s all in the crystals. Acta Crystallogr. Sect. D-Biol. Crystallogr. 2011, 67, 243–248. [Google Scholar] [CrossRef]
- Wang, R.; Wang, S.; Xu, Y.; Yu, X. Enhancing the thermostability of Rhizopus chinensis lipase by rational design and MD simulations. Int. J. Biol. Macromol. 2020, 160, 1189–1200. [Google Scholar] [CrossRef]
- Zhou, Y.B.; Cao, J.W.; Sun, X.B.; Wang, H.; Gao, D.Y.; Li, Y.N.; Wu, K.Y.; Wang, J.K.; Qian, G.Y.; Wang, Q. Enhanced stability of a rumen-derived xylanase Using SpyTag/SpyCatcher cyclization. World J. Microbiol. Biotechnol. 2020, 36, 33. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Li, J.; Teng, C.; Li, X. Cloning, over expression and characterization of a xylanase gene from a novel Streptomyces rameus L2001 in Pichia pastoris. J. Mol. Catal. B Enzym. 2016, 131, 85–93. [Google Scholar] [CrossRef]
- Wu, Q.; Zhang, C.; Zhu, W.; Lu, H.; Li, X.; Yang, Y.; Xu, Y.; Li, W. Improved thermostability, acid tolerance as well as catalytic efficiency of Streptomyces rameus L2001 GH11 xylanase by N-terminal replacement. Enzym. Microb. Technol. 2023, 162, 110143. [Google Scholar] [CrossRef]
- Landt, O.; Grunert, H.-P.; Hahn, U. A general method for rapid site-directed mutagenesis using the polymerase chain reaction. Gene 1990, 96, 125–128. [Google Scholar] [CrossRef]
- Tao, H.; Cornish, V.W. Milestones in directed enzyme evolution. Curr. Opin. Chem. Biol. 2002, 6, 858–864. [Google Scholar] [CrossRef]
- Li, Q.; Sun, B.; Jiang, H.; Hou, J.; Yang, R.; Xiong, K.; Xu, Y.; Li, X. Engineering a xylanase from Streptomyce rochei L10904 by mutation to improve its catalytic characteristics. Int. J. Biol. Macromol. 2017, 101, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Teng, C.; Xu, Y.; Xiong, K.; Xu, Y.; Li, L.; Li, X.; Li, X. Improving special hydrolysis characterization into Talaromyces thermophilus F1208 xylanase by engineering of N-terminal extension and site-directed mutagenesis in C-terminal. Int. J. Biol. Macromol. 2017, 96, 451–458. [Google Scholar] [CrossRef]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Frovenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Bailey, M.J.; Biely, P.; Poutanen, K. Interlaboratory testing of methods for assay of xylanase activity. J. Biotechnol. 1992, 23, 257–270. [Google Scholar] [CrossRef]
- Wu, Q.; Fan, G.; Yu, T.; Sun, B.; Tang, H.; Teng, C.; Yang, R.; Li, X. Biochemical characteristics of the mutant xylanase T-XynC(122)C(166) and production of xylo-oligosaccharides from corncobs. Ind. Crops Prod. 2019, 142, 111848. [Google Scholar] [CrossRef]
- You, C.; Huang, Q.; Xue, H.; Xu, Y.; Lu, H. Potential hydrophobic interaction between two cysteines in interior hydrophobic region improves thermostability of a family 11 xylanase from Neocallimastix patriciarum. Biotechnol. Bioeng. 2010, 105, 861–870. [Google Scholar] [CrossRef]
- Li, L.; Zhang, S.; Wu, W.; Guan, W.; Deng, Z.; Qiao, H. Enhancing thermostability of Yarrowia lipolytica lipase 2 through engineering multiple disulfide bonds and mitigating reduced lipase production associated with disulfide bonds. Enzym. Microb. Technol. 2019, 126, 41–49. [Google Scholar] [CrossRef]
- Singh, C.; Roy-Chowdhuri, S. Quantitative real-time PCR: Recent advances. In Clinical Applications of PCR; Luthra, R., Singh, R.R., Patel, K.P., Eds.; Springer: New York, NY, USA, 2016; pp. 161–176. ISBN 978-1-4939-3360-0. [Google Scholar] [CrossRef]
- Teng, C.; Jiang, Y.; Xu, Y.; Li, Q.; Li, X.; Fan, G.; Xiong, K.; Yang, R.; Zhang, C.; Ma, R.; et al. Improving the thermostability and catalytic efficiency of GH11 xylanase PjxA by adding disulfide bridges. Int. J. Biol. Macromol. 2019, 128, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Goldschmidt, L.; Cooper, D.R.; Derewenda, Z.S.; Eisenberg, D. Toward rational protein crystallization: A web server for the design of crystallizable protein variants. Protein Sci. 2007, 16, 1569–1576. [Google Scholar] [CrossRef] [Green Version]
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Rauscher, S.; Nawrocki, G.; Ran, T.; Feig, M.; de Groot, B.L.; Grubmüller, H.; MacKerell, A.D. Charmm36m: An improved force field for folded and intrinsically disordered proteins. Nat. Methods 2016, 14, 71–73. [Google Scholar] [CrossRef] [Green Version]
- Grant, B.J.; Skjærven, L.; Yao, X.Q. The Bio3D packages for structural bioinformatics. Protein Sci. 2021, 30, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Dalby, P.A. Coupled molecular dynamics mediate long- and short-range epistasis between mutations that affect stability and aggregation kinetics. Proc. Natl. Acad. Sci. USA 2018, 115, E11043–E11052. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, R.; Maranas, C.D. From directed evolution to computational enzyme engineering—A review. AIChE J. 2020, 66, e16847. [Google Scholar] [CrossRef]
- Purmonen, M.; Valjakka, J.; Takkinen, K.; Laitinen, T.; Rouvinen, J. Molecular dynamics studies on the thermostability of family 11 xylanases. Protein Eng. Des. Sel. 2007, 20, 551–559. [Google Scholar] [CrossRef] [Green Version]
- Cooper, D.R.; Boczek, T.; Grelewska, K.; Pinkowska, M.; Sikorska, M.; Zawadzki, M.; Derewenda, Z. Protein crystallization by surface entropy reduction: Optimization of the SER strategy. Acta Cryst. D Biol. Cryst. 2007, 63, 636–645. [Google Scholar] [CrossRef]
- Parthasarathy, G.; Cummings, R.; Becker, J.W.; Soisson, S.M. Surface-entropy reduction approaches to manipulate crystal forms of β-ketoacyl acyl carrier protein synthase II from Streptococcus pneumoniae. Acta Cryst. D Biol Cryst. 2008, 64, 141–148. [Google Scholar] [CrossRef]
- Zhou, Y.; Pérez, B.; Hao, W.; Lv, J.; Gao, R.; Guo, Z. The additive mutational effects from surface charge engineering: A compromise between enzyme activity, thermostability and ionic liquid tolerance. Biochem. Eng. J. 2019, 148, 195–204. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, K.; Chen, X.; Chu, X.; Sun, F.; Dong, Z. Five mutations in N-terminus confer thermostability on mesophilic xylanase. Biochem. Biophys. Res. Commun. 2010, 395, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Huang, H. Engineering proteins for thermostability through rigidifying flexible sites. Biotechnol. Adv. 2014, 32, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Pace, C.N.; Fu, H.L.; Fryar, K.L.; Landua, J.; Trevino, S.R.; Shirley, B.A.; Hendricks, M.M.; Iimura, S.; Gajiwala, K.; Scholtz, J.M.; et al. Contribution of hydrophobic interactions to protein stability. J. Mol. Biol. 2011, 408, 514–528. [Google Scholar] [CrossRef] [Green Version]
- Pace, C.N. Contribution of the hydrophobic effect to globular protein stability. J. Mol. Biol. 1992, 226, 29–35. [Google Scholar] [CrossRef]
- Okada, J.; Okamoto, T.; Mukaiyama, A.; Tadokoro, T.; You, D.-J.; Chon, H.; Koga, Y.; Takano, K.; Kanaya, S. Evolution and thermodynamics of the slow unfolding of hyperstable monomeric proteins. BMC Evol. Biol. 2010, 10, 207. [Google Scholar] [CrossRef] [Green Version]
- Xing, H.; Zou, G.; Liu, C.; Chai, S.; Yan, X.; Li, X.; Liu, R.; Yang, Y.; Zhou, Z. Improving the thermostability of a GH11 xylanase by directed evolution and rational design guided by b-factor analysis. Enzym. Microb. Technol. 2021, 143, 109720. [Google Scholar] [CrossRef]
- Tang, F.; Chen, D.; Yu, B.; Luo, Y.; Zheng, P.; Mao, X.; Yu, J.; He, J. Improving the thermostability of Trichoderma reesei xylanase 2 by introducing disulfide bonds. Electron. J. Biotechnol. 2017, 26, 52–59. [Google Scholar] [CrossRef]
- Xiong, K.; Hou, J.; Jiang, Y.; Li, X.; Teng, C.; Li, Q.; Fan, G.; Yang, R.; Zhang, C. Mutagenesis of N-terminal residues confer thermostability on a Penicillium janthinellum MA21601 xylanase. BMC Biotechnol. 2019, 19, 51. [Google Scholar] [CrossRef]
- Cai, L.; Zhang, M.; Shao, T.; He, Y.; Li, J.; Ren, B.; Zhou, C. Effect of introducing disulfide bridges in C-terminal structure on the thermostability of xylanase Xynzf-2 from Aspergillus niger. J. Gen. Appl. Microbiol. 2019, 65, 240–245. [Google Scholar] [CrossRef] [Green Version]
- Wakarchuk, W.W.; Sung, W.L.; Campbell, R.L.; Cunningham, A.; Watson, D.C.; Yaguchi, M. Thermostabilization of the Bacillus circulans xylanase by the introduction of disulfide bonds. Protein Eng. 1994, 7, 1379–1386. [Google Scholar] [CrossRef] [PubMed]
- Boonyaputthikul, H.; Muhammad, A.; Roekring, S.; Rattanarojpong, T.; Khunrae, P.; Sutthibutpong, T. Synergistic effects between the additions of a disulphide bridge and an N-terminal hydrophobic sidechain on the binding pocket tilting and enhanced Xyn11A activity. Arch. Biochem. Biophys. 2019, 672, 108068. [Google Scholar] [CrossRef]
- Waldhauer, M.C.; Schmitz, S.N.; Ahlmann-Eltze, C.; Gleixner, J.G.; Schmelas, C.C.; Huhn, A.G.; Bunne, C.; Büscher, M.; Horn, M.; Klughammer, N.; et al. Backbone circularization of Bacillus subtilis family 11 xylanase increases its thermostability and its resistance against aggregation. Mol. Biosyst. 2015, 11, 3231–3243. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Tang, F.; Chen, D.; Yu, B.; Luo, Y.; Zheng, P.; Mao, X.; Yu, J.; Yu, F. Design, expression and functional characterization of a thermostable xylanase from Trichoderma reesei. PLoS One 2019, 14, e0210548. [Google Scholar] [CrossRef]
- Wang, Y.; Chang, Y.; Jia, R.; Sun, H.; Tian, J.W.; Luo, H.; Yu, H.; Shen, Z. SpyTag/SpyCatcher cyclization and covalent immobilization in enhancing cephalosporin C acylase stability. Process Biochem. 2020, 95, 260–268. [Google Scholar] [CrossRef]
- Xu, Q.; Ming, D.; Shi, C.; Lu, D.; Tang, S.; Jiang, L.; Huang, H. Catcher/Tag cyclization introduces electrostatic interaction mediated protein-protein interactions to enhance the thermostability of luciferase. Process Biochem. 2019, 80, 64–71. [Google Scholar] [CrossRef]
- Li, Y.; Li, C.; Huang, H.; Rao, S.; Zhang, Q.; Zhou, J.; Li, J.; Du, G.; Liu, S. Significantly enhanced thermostability of Aspergillus niger xylanase by modifying its highly flexible regions. J. Agric. Food Chem. 2022, 70, 4620–4630. [Google Scholar] [CrossRef]
- Xiong, H.; Fenel, F.; Leisola, M.; Turunen, O. Engineering the thermostability of Trichoderma reesei endo-1,4-β-xylanase II by combination of disulphide bridges. Extremophiles 2004, 8, 393–400. [Google Scholar] [CrossRef]
- Ruller, R.; Deliberto, L.; Ferreira, T.L.; Ward, R.J. Thermostable variants of the recombinant xylanase a from Bacillus subtilis produced by directed evolution show reduced heat capacity changes. Proteins: Struct. Funct. Genet. 2008, 70, 1280–1293. [Google Scholar] [CrossRef]
- Yu, H.; Ma, S.; Li, Y.; Dalby, P.A. Hot spots-making directed evolution easier. Biotechnol. Adv. 2022, 5, 107926. [Google Scholar] [CrossRef]
- Yao, X.; Kang, T.; Pu, Z.; Zhang, T.; Lin, J.; Yang, L.; Yu, H.; Wu, M. Sequence and structure-guided engineering of urethanase from Agrobacterium tumefaciens d3 for improved catalytic activity. J. Agric. Food Chem. 2022, 70, 7267–7278. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.M.; Yao, B.; Meng, K.; Wang, Y.R.; Bai, Y.G.; Wu, N.F. Introduction of a disulfide bridge enhances the thermostability of a Streptomyces olivaceoviridis xylanase mutant. J. Ind. Microbiol. Biotechnol. 2007, 34, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Fenel, F.; Leisola, M.; Jänis, J.; Turunen, O. A de novo designed N-terminal disulphide bridge stabilizes the Trichoderma reesei endo-1,4-β-xylanase II. J. Biotechnol. 2004, 108, 137–143. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| XynA | Q24A | K104A | K143A | XynA-VT | XynA-NT | XynA-SN | XynA-Spy | |
|---|---|---|---|---|---|---|---|---|
| Optimum pH | 5.5 a | 5.0 b | 5.0 b | 5.0 b | 5.5 a | 5.0 b | 5.5 a | 5.0 b |
| pH stability rage (>80%) | 3.5–9.0 h | 2.0–8.5 e | 2.0–11.5 a | 2.0–11.0 b | 2.0–10.0 c | 2.0–9.5 d | 2.5–7.5 g | 2.5–10.5 f |
| Optimum temperature (°C) | 60 d | 65 b | 65 b | 60–65 c | 70 a | 70 a | 65 b | 60 d |
| Thermostability (65 °C, 30 min) | 20% d | >40% b | >40% b | >20% c | >90% a | >90% a | >90% a | 20% d |
| t1/260°C (min) | 86.64 d | 76.17 e | 88.87 c | 69.31 f | 1155.25 a | 693.15 b | nd h | 46.83 g |
| t1/270°C (min) | nd d | nd d | nd d | nd d | 92.42 a | 35.01 b | 210.04(s)c | nd d |
| Tm (°C) | 73.5 c | 73.5 c | 73.5 c | 73.5 c | 83.5 a | 78.5 b | 73.5 c | 72 d |
| XynA | Q24A | K104A | K143A | XynA-VT | XynA-NT | XynA-SN | XynA-Spy | |
|---|---|---|---|---|---|---|---|---|
| Beechwood xylan (U/mg) | 1358.86 ± 151.42 a | 1710.65 ± 58.88 a | 1532.84 ± 55.30 a | 1710.01 ± 59.64 a | 1963.81 ± 11.65 a | 2017.74 ± 106.78 a | 2010.55 ± 151.42 a | 400.38 ± 54.05 a |
| Birchwood xylan (U/mg) | 630.40 ± 37.70 b | 920.98 ± 2.26 b | 1058.89 ± 24.84 b | 1069.93 ± 33.26 b | 1585.67 ± 9.22 b | 1147.59 ± 96.21 b | 1198.67 ± 212.03 b | 206.72 ± 39.85 b |
| Oat-spelt xylan (U/mg) | 480.10 ± 29.30 c | 806.64 ± 26.30 c | 1013.57 ± 102.80 b | 1013.16 ± 5.73 b | 923.56 ± 45.62 c | 432.98 ± 64.11 c | 401.85 ± 105.75 c | 139.39 ± 11.79 c |
| XynA | Q24A | K104A | K143A | XynA-VT | XynA-NT | XynA-SN | XynA-Spy | |
|---|---|---|---|---|---|---|---|---|
| Km (mg/mL) | 19.18 ± 1.67 b | 8.31 ± 0.10 d | 19.46 ± 0.70 ab | 11.75 ± 1.02 c | 19.99 ± 1.24 ab | 13.26 ± 0.97 c | 17.83 ± 0.70 b | 21.57 ± 2.02 a |
| Vmax (μmol/min/mg) | 3484.00 ± 165.00 c | 3120.00 ± 516.00 c | 5185.00 ± 577.00 b | 3993.00 ± 135.00 c | 6524.43 ± 215.98 a | 3775.05 ± 487.55 c | 5195.38 ± 1237.22 b | 1417.45 ± 377.17 d |
| kcat (/s) | 1208.00 ± 57.00 cd | 1080.00 ± 179.00 cd | 1795.00 ± 200.00 b | 1382.00 ± 47.00 c | 2265.06 ± 74.75 a | 1309.94 ± 168.76 c | 1803.67 ± 428.25 b | 885.67 ± 130.55 d |
| kcat/Km (mL/s/mg) | 62.98 ± 7.78 e | 129.96 ± 12.83 a | 92.24 ± 3.12 d | 117.62 ± 3.08 ab | 113.31 ± 1.14 bc | 98.72 ± 8.52 d | 101.16 ± 9.29 cd | 41.07 ± 9.05 f |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, W.; Qin, L.; Xu, Y.; Lu, H.; Wu, Q.; Li, W.; Zhang, C.; Li, X. Three Molecular Modification Strategies to Improve the Thermostability of Xylanase XynA from Streptomyces rameus L2001. Foods 2023, 12, 879. https://doi.org/10.3390/foods12040879
Zhu W, Qin L, Xu Y, Lu H, Wu Q, Li W, Zhang C, Li X. Three Molecular Modification Strategies to Improve the Thermostability of Xylanase XynA from Streptomyces rameus L2001. Foods. 2023; 12(4):879. https://doi.org/10.3390/foods12040879
Chicago/Turabian StyleZhu, Weijia, Liqin Qin, Youqiang Xu, Hongyun Lu, Qiuhua Wu, Weiwei Li, Chengnan Zhang, and Xiuting Li. 2023. "Three Molecular Modification Strategies to Improve the Thermostability of Xylanase XynA from Streptomyces rameus L2001" Foods 12, no. 4: 879. https://doi.org/10.3390/foods12040879
APA StyleZhu, W., Qin, L., Xu, Y., Lu, H., Wu, Q., Li, W., Zhang, C., & Li, X. (2023). Three Molecular Modification Strategies to Improve the Thermostability of Xylanase XynA from Streptomyces rameus L2001. Foods, 12(4), 879. https://doi.org/10.3390/foods12040879