
Impact of Starch-Rich Food Matrices on Black Rice Anthocyanin Accessibility and Carbohydrate Digestibility

Abstract
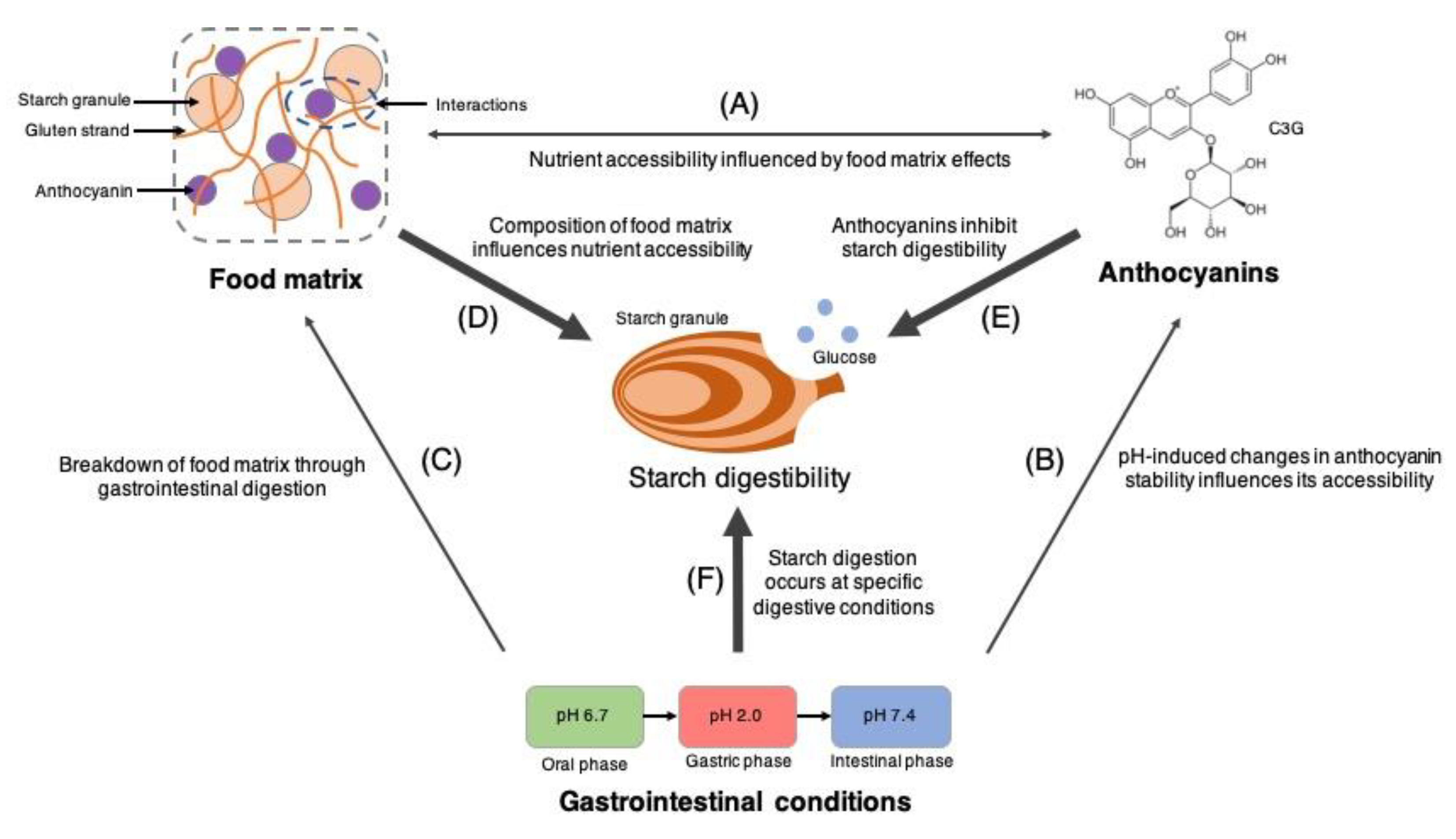
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Bread Sample Preparation
2.3. Evaluation of Starch Digestibility
2.4. Evaluation of Anthocyanin Accessibility
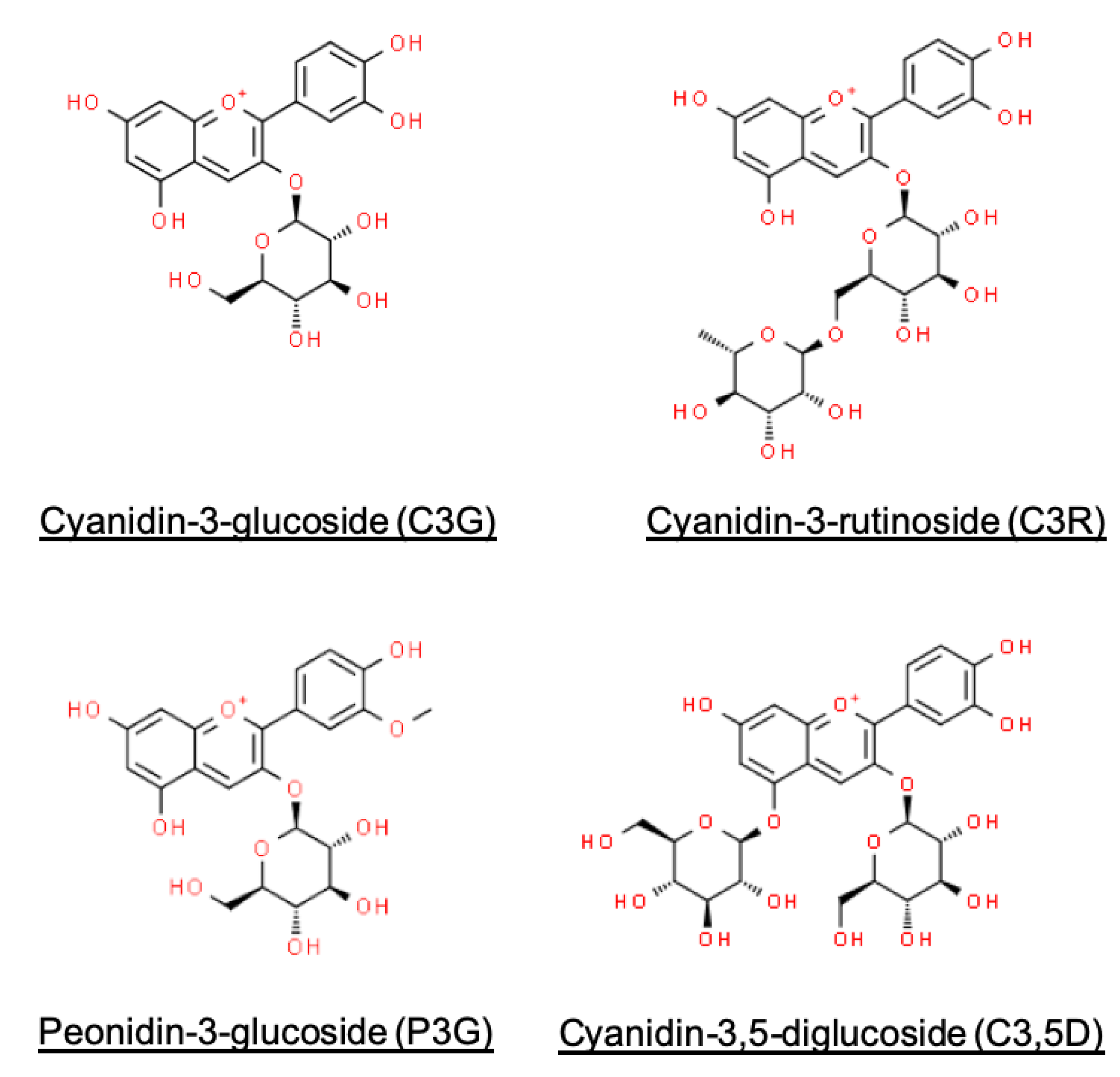
2.5. Determination of Anthocyanin Content
2.6. Statistical Analysis
3. Results
3.1. Total Monomeric Anthocyanin Content
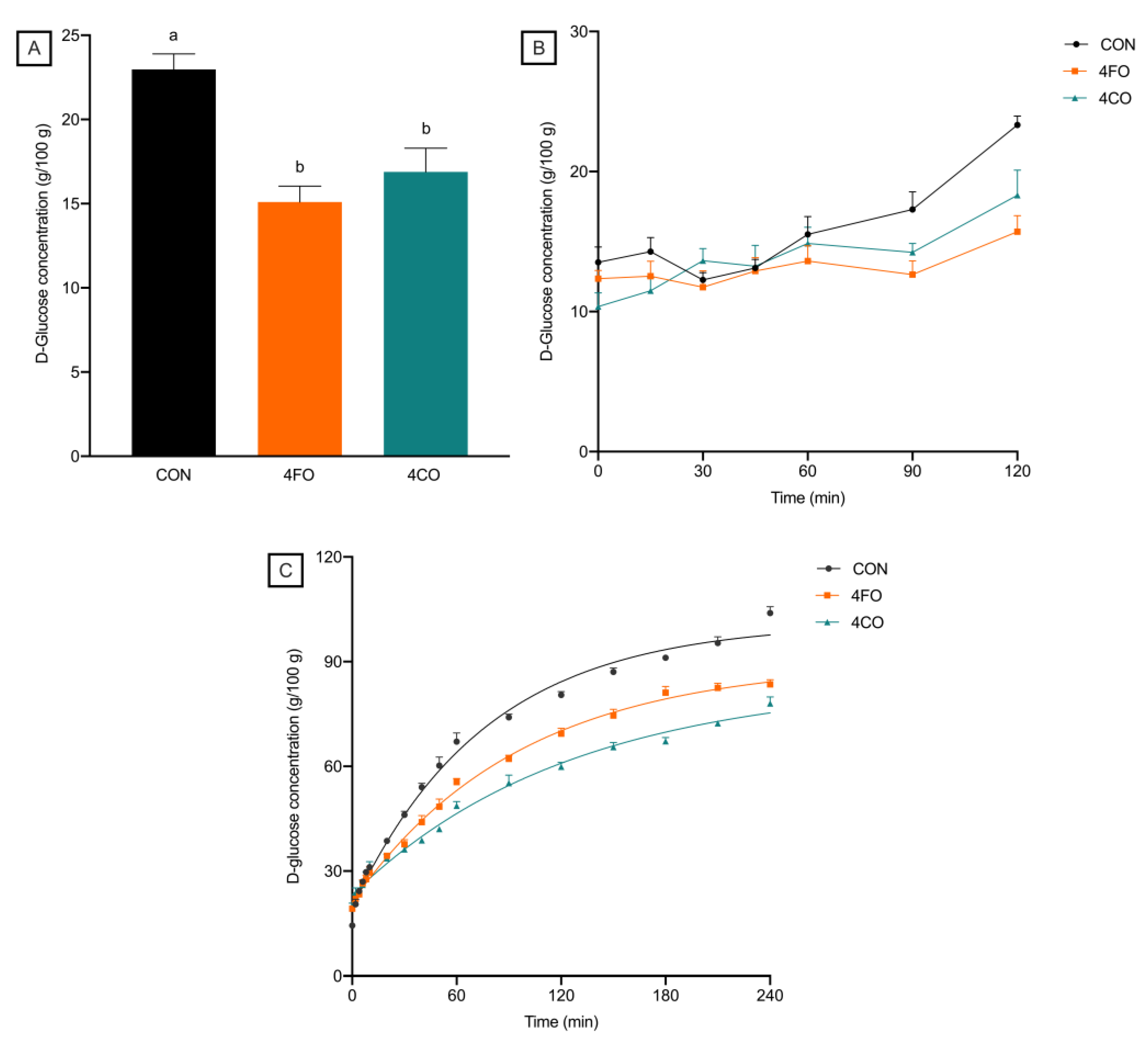
3.2. Effects of BRAE Fortification and Co-Digestion on Bread Digestibility
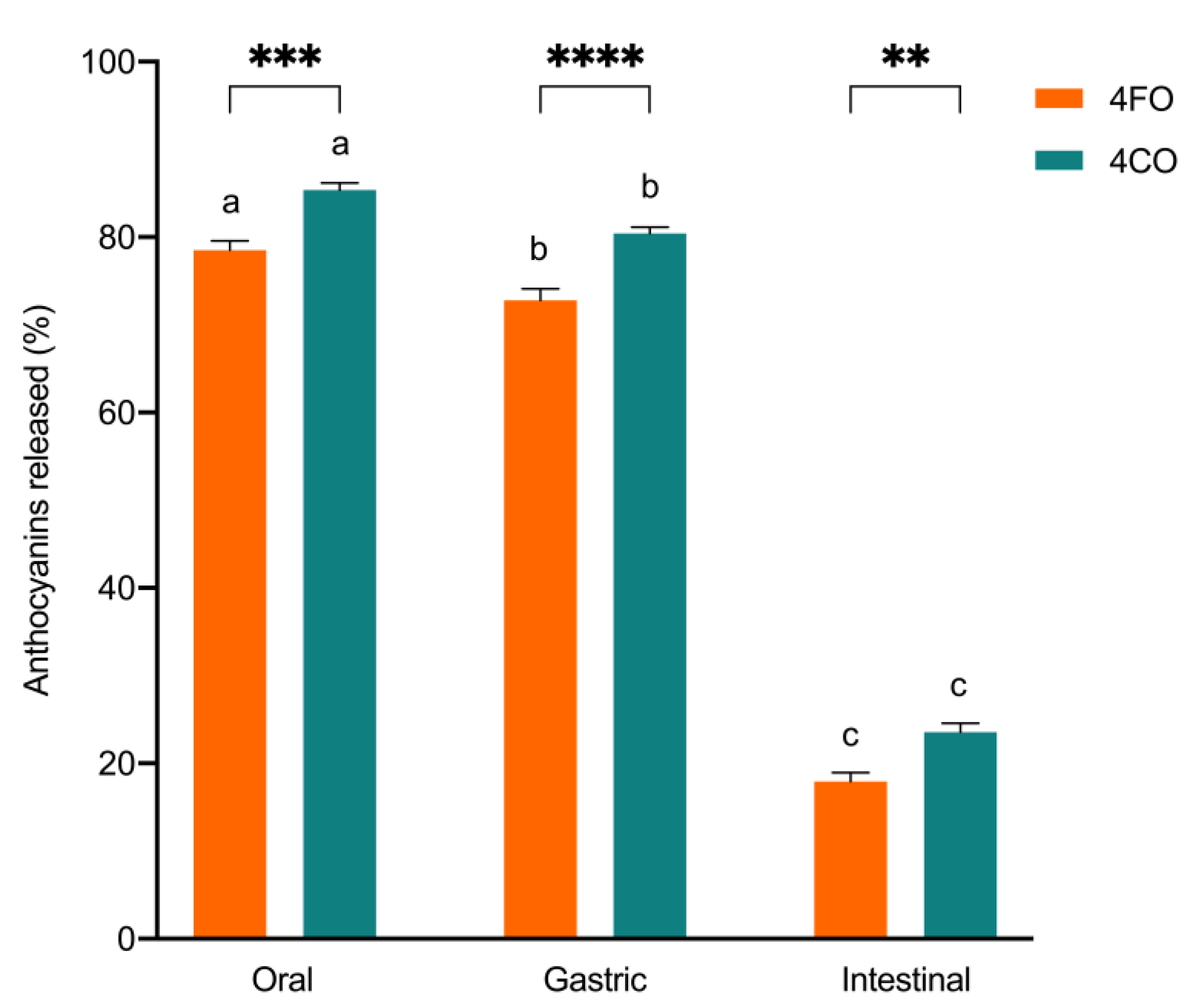
3.3. Simulated Gastrointestinal Accessibility of Anthocyanin
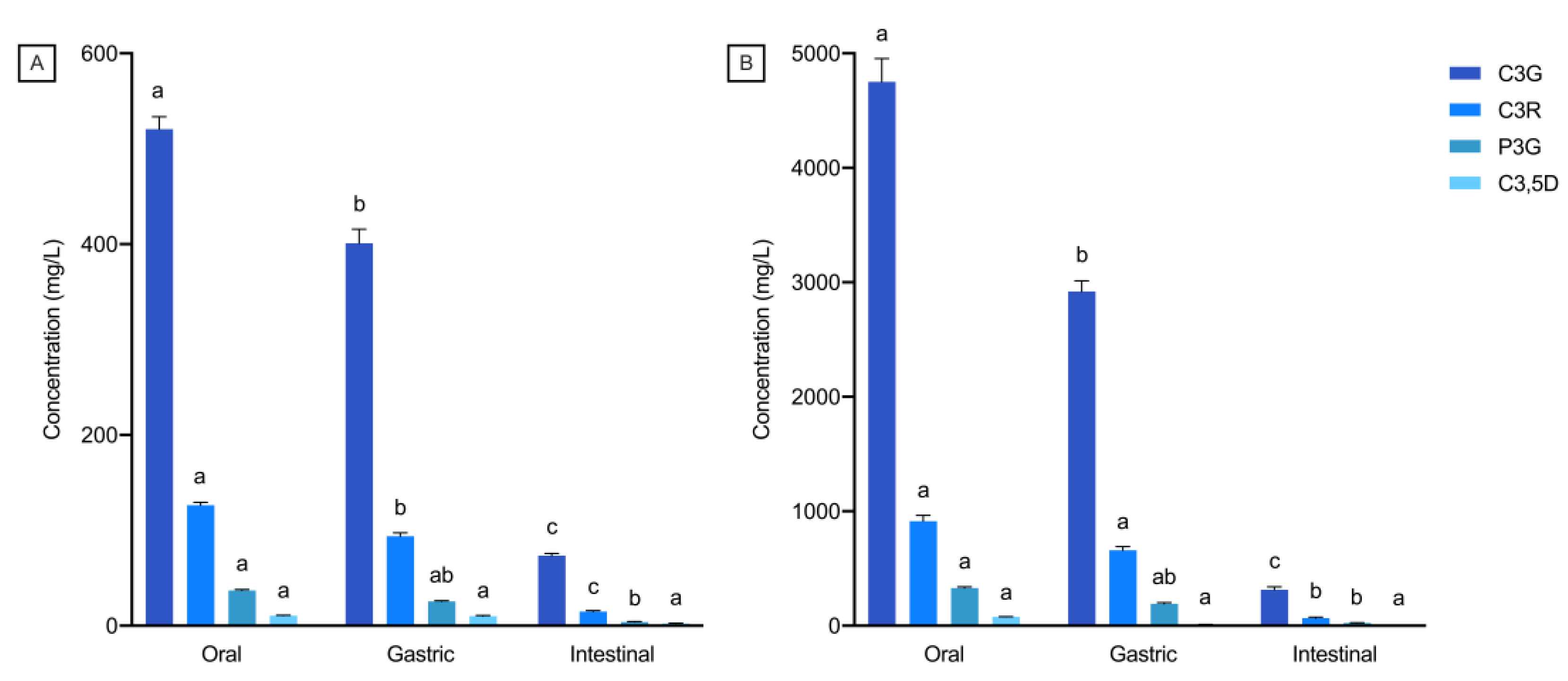
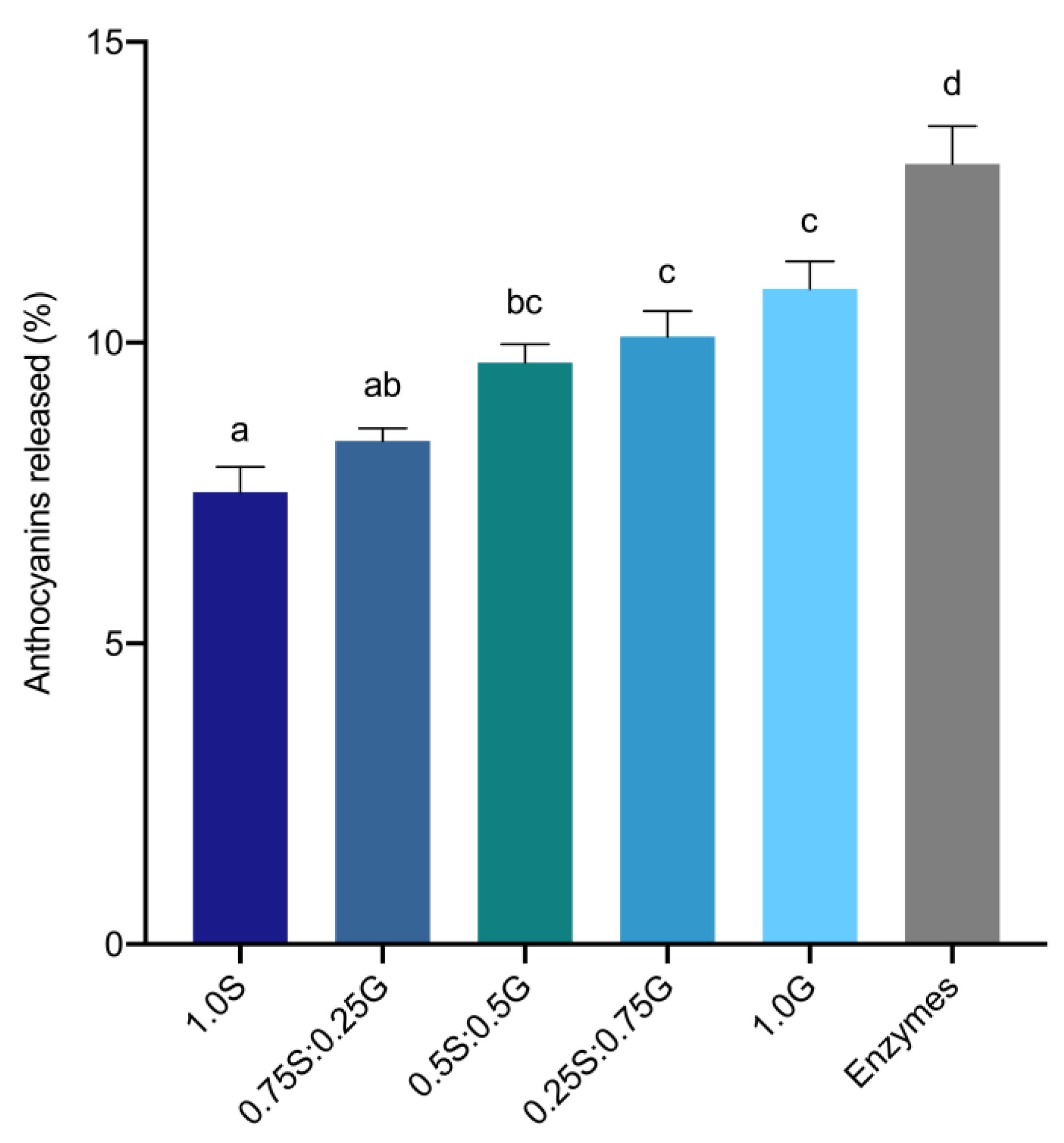
3.4. Interactions between Anthocyanins and Constituents of Bread Digestion Matrices
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A. Preparation of Reagents for the Simulated Digestion Model

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| For 1000 mL of Each of SF | ||||||||
|---|---|---|---|---|---|---|---|---|
| Stock Solution | Stock Concentration | SSF (pH 7) | SGF (pH 3) | SIF (pH 7) | ||||
| Volume of Stock Added | Final Concentration | Volume of Stock Added | Final Concentration | Volume of Stock Added | Final Concentration | |||
| (g/L) | (M) | (mL) | (mM) | (mL) | (mM) | (mL) | (mM) | |
| KCl | 37.3 | 0.5 | 37.75 | 15.1 | 17.25 | 6.9 | 17 | 6.8 |
| KH2PO4 | 68 | 0.5 | 9.25 | 3.7 | 2.25 | 0.9 | 2 | 0.8 |
| NaHCO3 | 84 | 1 | 17 | 13.6 | 31.25 | 25 | 106.25 | 85 |
| NaCl | 117 | 2 | 0 | 0 | 29.5 | 47.2 | 24 | 38.4 |
| MgCl2(H2O)6 | 30.5 | 0.15 | 1.25 | 0.15 | 1 | 0.12 | 2.75 | 0.33 |
| (NH4)2CO3 | 48 | 0.5 | 0.15 | 0.06 | 1.25 | 0.5 | 0 | 0 |
| CaCl2(H2O)2 | 44.1 | 0.3 | 0.0625 | 1.5 | 0.0125 | 0.15 | 0.1 | 0.6 |
| HCl | 6 | 0.225 | 1.1 | 3.25 | 15.6 | 1.75 | 8.4 | |
| Oral Phase (pH 7) | Gastric Phase (pH 3) | Intestinal Phase (pH 7) | ||
|---|---|---|---|---|
| Sample (g) | 2 | 4 | 8 | |
| Simulation fluid (μL) | 1300 | 2950 | 4510 | |
| 0.3M CaCl2 (μL) | 10 | 2 | 16 | |
| Enzyme | α-amylase from porcine pancreas | Pepsin from porcine pancreas | Pancreatin from porcine pancreas | α-amyloglucosidase from Aspergillus niger |
| Final enzyme activity (U/mL) | 75 | 2000 | 100 | 1.3 |
| Volume of 6M HCl/NaOH | Added to adjust to pH 7 | Added to adjust to pH 3 | Added to adjust to pH 7 | |
| Volume of water | Top up to total volume | Top up to total volume | Top up to total volume | |
| Total volume (mL) | 4 | 8 | 16 | |
Appendix B. Quantification of Anthocyanin Content
| BRAE | 4FO | |||
|---|---|---|---|---|
| Incorporated into Dough | Post-Baking | Retention | ||
| (mg/g Extract) | (mg/g Dough) | (mg/g Bread) | (%) | |
| TMAC (C3G equivalents) | 240.6 (4) | 624.3 (7) | 127.3 (1) | 20.4 |
| C3G | 211.2 (6) | 546.8 (3) | 112.9 (3) | 20.6 |
| C3R | 25.5 (0.2) | 66.4 (1.7) | 21.2 (0.5) | 31.9 |
| P3G | 7.0 (0.1) | 18.3 (0.5) | 3.7 (0.2) | 20.3 |
| C3,5D | 0.9 (0.1) | 2.2 (0.1) | 0.5 (0.0) | 23.7 |
Appendix C. Effects of BRAE Fortification and Co-Digestion of Bread Digestibility

Appendix D. Effects of Interactions between Anthocyanins and Digestion Matrix Constituents on Anthocyanin Accessibility
| Matrix | Starch (g) | Gluten (g) |
|---|---|---|
| 1.0S | 3.0 | 0 |
| 0.75S:0.25G | 2.25 | 0.75 |
| 0.5S:0.5G | 1.5 | 1.5 |
| 0.25S:0.75G | 0.75 | 2.25 |
| 1.0G | 0 | 3.0 |
References
- Yang, L.; Ling, W.; Du, Z.; Chen, Y.; Li, D.; Deng, S.; Liu, Z.; Yang, L. Effects of Anthocyanins on Cardiometabolic Health: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Adv. Nutr. 2017, 8, 684–693. [Google Scholar] [CrossRef] [Green Version]
- Victoria-Campos, C.I.; de Jesús Ornelas-Paz, J.; Rocha-Guzmán, N.E.; Gallegos-Infante, J.A.; Failla, M.L.; Pérez-Martínez, J.D.; Rios-Velasco, C.; Ibarra-Junquera, V. Gastrointestinal metabolism and bioaccessibility of selected anthocyanins isolated from commonly consumed fruits. Food Chem. 2022, 383, 132451. [Google Scholar] [CrossRef] [PubMed]
- Ito, V.C.; Lacerda, L.G. Black rice (Oryza sativa L.): A review of its historical aspects, chemical composition, nutritional and functional properties, and applications and processing technologies. Food Chem. 2019, 301, 125304. [Google Scholar] [CrossRef]
- Akkarachiyasit, S.; Charoenlertkul, P.; Yibchok-anun, S.; Adisakwattana, S. Inhibitory Activities of Cyanidin and Its Glycosides and Synergistic Effect with Acarbose against Intestinal α-Glucosidase and Pancreatic α-Amylase. Int. J. Mol. Sci. 2010, 11, 3387–3396. [Google Scholar] [CrossRef] [Green Version]
- Ou, S.J.L.; Yu, J.; Zhou, W.; Liu, M.H. Effects of anthocyanins on bread microstructure, and their combined impact on starch digestibility. Food Chem. 2022, 374, 131744. [Google Scholar] [CrossRef]
- Sui, X.; Zhang, Y.; Zhou, W. Bread fortified with anthocyanin-rich extract from black rice as nutraceutical sources: Its quality attributes and in vitro digestibility. Food Chem. 2016, 196, 910–916. [Google Scholar] [CrossRef]
- Tarko, T.; Duda-Chodak, A. Influence of Food Matrix on the Bioaccessibility of Fruit Polyphenolic Compounds. J. Agric. Food Chem. 2020, 68, 1315–1325. [Google Scholar] [CrossRef]
- Kamiloglu, S.; Tomas, M.; Ozdal, T.; Capanoglu, E. Effect of food matrix on the content and bioavailability of flavonoids. Trends Food Sci. Technol. 2021, 117, 15–33. [Google Scholar] [CrossRef]
- Sengul, H.; Surek, E.; Nilufer-Erdil, D. Investigating the effects of food matrix and food components on bioaccessibility of pomegranate (Punica granatum) phenolics and anthocyanins using an in-vitro gastrointestinal digestion model. Food Res. Int. 2014, 62, 1069–1079. [Google Scholar] [CrossRef] [Green Version]
- Lang, Y.; Li, B.; Gong, E.; Shu, C.; Si, X.; Gao, N.; Zhang, W.; Cui, H.; Meng, X. Effects of α-casein and β-casein on the stability, antioxidant activity and bioaccessibility of blueberry anthocyanins with an in vitro simulated digestion. Food Chem. 2021, 334, 127526. [Google Scholar] [CrossRef]
- Pineda-Vadillo, C.; Nau, F.; Guerin-Dubiard, C.; Jardin, J.; Lechevalier, V.; Sanz-Buenhombre, M.; Guadarrama, A.; Tóth, T.; Csavajda, É.; Hingyi, H.; et al. The food matrix affects the anthocyanin profile of fortified egg and dairy matrices during processing and in vitro digestion. Food Chem. 2017, 214, 486–496. [Google Scholar] [CrossRef]
- Pešić, M.B.; Milinčić, D.D.; Kostić, A.Ž.; Stanisavljević, N.S.; Vukotić, G.N.; Kojić, M.O.; Gašić, U.M.; Barać, M.B.; Stanojević, S.P.; Popović, D.A.; et al. In vitro digestion of meat- and cereal-based food matrix enriched with grape extracts: How are polyphenol composition, bioaccessibility and antioxidant activity affected? Food Chem. 2019, 284, 28–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sui, X. Impact of Food Processing on Anthocyanins; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar] [CrossRef]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Durst, R.W.; Wrolstad, R.E. Determination of Total Monomeric Anthocyanin Pigment Content of Fruit Juices, Beverages, Natural Colorants, and Wines by the pH Differential Method: Collaborative Study. J. of AOAC INTERNATIONAL 2019, 88, 1269–1278. [Google Scholar] [CrossRef] [Green Version]
- Pedro, A.C.; Granato, D.; Rosso, N.D. Extraction of anthocyanins and polyphenols from black rice (Oryza sativa L.) by modeling and assessing their reversibility and stability. Food Chem. 2016, 191, 12–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minich, D.M. A Review of the Science of Colorful, Plant-Based Food and Practical Strategies for “Eating the Rainbow”. J. Nutr. Metab. 2019, 2019, 2125070. [Google Scholar] [CrossRef]
- Kalita, D.; Holm, D.G.; LaBarbera, D.V.; Petrash, J.M.; Jayanty, S.S. Inhibition of α-glucosidase, α-amylase, and aldose reductase by potato polyphenolic compounds. PLoS ONE 2018, 13, e0191025. [Google Scholar] [CrossRef]
- Shozib, H.B.; Islam, M.M.; Mahmud, S.A.S.; Bari, M.N.; Akter, N.; Jahan, S.; Hosen, S.; Hossain, M.N.; Nabi, A.H.M.N.; Siddiquee, M.A.; et al. Application of Cyanidin-3-Glucosides as a functional food ingredient in rice-based bakery products. Saudi J. Biol. Sci. 2021, 28, 7472–7480. [Google Scholar] [CrossRef]
- Aguilera, J.M. The food matrix: Implications in processing, nutrition and health. Crit. Rev. Food Sci. Nutr. 2019, 59, 3612–3629. [Google Scholar] [CrossRef]
- Capuano, E.; Janssen, A.E.M. Food Matrix and Macronutrient Digestion. Annu. Rev. Food Sci. Technol. 2021, 12, 193–212. [Google Scholar] [CrossRef]
- Jenkins, D.J.; Thorne, M.J.; Wolever, T.M.; Jenkins, A.L.; Rao, A.V.; Thompson, L.U. The effect of starch-protein interaction in wheat on the glycemic response and rate of in vitro digestion. Am. J. Clin. Nutr. 1987, 45, 946–951. [Google Scholar] [CrossRef] [Green Version]
- Kan, L.; Oliviero, T.; Verkerk, R.; Fogliano, V.; Capuano, E. Interaction of bread and berry polyphenols affects starch digestibility and polyphenols bio-accessibility. J. Funct. Foods 2020, 68, 103924. [Google Scholar] [CrossRef]
- Sun, L.; Gidley, M.J.; Warren, F.J. Tea polyphenols enhance binding of porcine pancreatic α-amylase with starch granules but reduce catalytic activity. Food Chem. 2018, 258, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Amoako, D.B.; Awika, J.M. Polymeric tannins significantly alter properties and in vitro digestibility of partially gelatinized intact starch granule. Food Chem. 2016, 208, 10–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, Z.; Qin, P.; Zhang, Y.; Cui, S.; Ren, G. Identification of anthocyanins isolated from black rice (Oryza sativa L.) and their degradation kinetics. Food Res. Int. 2013, 50, 691–697. [Google Scholar] [CrossRef]
- Fleschhut, J.; Kratzer, F.; Rechkemmer, G.; Kulling, S.E. Stability and biotransformation of variousdietary anthocyanins in vitro. Eur. J. Nutr. 2006, 45, 7–18. [Google Scholar] [CrossRef]
- McDougall, G.J.; Dobson, P.; Smith, P.; Blake, A.; Stewart, D. Assessing Potential Bioavailability of Raspberry Anthocyanins Using an in Vitro Digestion System. J. Agric. Food Chem. 2005, 53, 5896–5904. [Google Scholar] [CrossRef] [PubMed]
- Lamothe, S.; Azimy, N.; Bazinet, L.; Couillard, C.; Britten, M. Interaction of green tea polyphenols with dairy matrices in a simulated gastrointestinal environment. Food Funct. 2014, 5, 2621–2631. [Google Scholar] [CrossRef]
- Vitali Čepo, D.; Radić, K.; Turčić, P.; Anić, D.; Komar, B.; Šalov, M. Food (Matrix) Effects on Bioaccessibility and Intestinal Permeability of Major Olive Antioxidants. Foods 2020, 9, 1831. [Google Scholar] [CrossRef]
- Song, J.; Yu, Y.; Chen, M.; Ren, Z.; Chen, L.; Fu, C.; feei Ma, Z.; Li, Z. Advancement of Protein- and Polysaccharide-Based Biopolymers for Anthocyanin Encapsulation. Front. Nutr. 2022, 9. [Google Scholar] [CrossRef]
- Dangles, O.; Fenger, J.-A. The Chemical Reactivity of Anthocyanins and Its Consequences in Food Science and Nutrition. Molecules 2018, 23, 1970. [Google Scholar] [CrossRef] [PubMed] [Green Version]





| Parameter | CON | 4FO | 4CO | p | |
|---|---|---|---|---|---|
| Oral | Cmean (mmol/L) | 23.0 (0.9) a | 15.1 (0.9) b | 16.9 (1.4) b | 0.0001 |
| Intestinal | Cmean (mmol/L) | 41.3 (7.1) | 29.1 (5.6) | 24.3 (4.6) | 0.116 |
| AUC (mmol/L × min) | 18,134 (163) a | 15,494 (183) b | 13,669 (142) c | <0.0001 | |
| IAUC (mmol/L × min) | 14,681 (264) a | 10,877 (246) b | 8905 (301) c | <0.0001 | |
| k | 0.0131 (0.0009) a | 0.0105 (0.0008) ab | 0.00777 (0.001) b | 0.003 |
| Oral to Gastric | Gastric to Intestinal | p | |||
|---|---|---|---|---|---|
| 4FO | 4CO | 4FO | 4CO | ||
| Total anthocyanins (%) | 10.1 (0.9) a | 9.8 (0.8) a | 68.7 (0.8) b | 73.4 (0.7) c | <0.0001 |
| C3G (%) | 24.0 (2.6) a | 37.3 (2.5) b | 83.0 (0.7) c | 90.2 (0.6) d | <0.0001 |
| C3R (%) | 24.2 (2.0) a | 25.9 (4.6) a | 81.5 (0.6) b | 90.0 (1.3) c | <0.0001 |
| P3G (%) | 34.5 (1.8) a | 44.5 (2.8) b | 81.0 (1.2) c | 88.3 (1.1) d | <0.0001 |
| C3,5D (%) | 29.2 (2.5) a | 58.9 (2.1) b | 58.8 (3.1) b | 69.2 (1.4) c | <0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ou, S.J.L.; Fu, A.S.; Liu, M.H. Impact of Starch-Rich Food Matrices on Black Rice Anthocyanin Accessibility and Carbohydrate Digestibility. Foods 2023, 12, 880. https://doi.org/10.3390/foods12040880
Ou SJL, Fu AS, Liu MH. Impact of Starch-Rich Food Matrices on Black Rice Anthocyanin Accessibility and Carbohydrate Digestibility. Foods. 2023; 12(4):880. https://doi.org/10.3390/foods12040880
Chicago/Turabian StyleOu, Sean Jun Leong, Amanda Simin Fu, and Mei Hui Liu. 2023. "Impact of Starch-Rich Food Matrices on Black Rice Anthocyanin Accessibility and Carbohydrate Digestibility" Foods 12, no. 4: 880. https://doi.org/10.3390/foods12040880
APA StyleOu, S. J. L., Fu, A. S., & Liu, M. H. (2023). Impact of Starch-Rich Food Matrices on Black Rice Anthocyanin Accessibility and Carbohydrate Digestibility. Foods, 12(4), 880. https://doi.org/10.3390/foods12040880