
Shoot Nutrition and Flavor Variation in Two Phyllostachys Species: Does the Quality of Edible Bamboo Shoot Diaphragm and Flesh Differ?


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
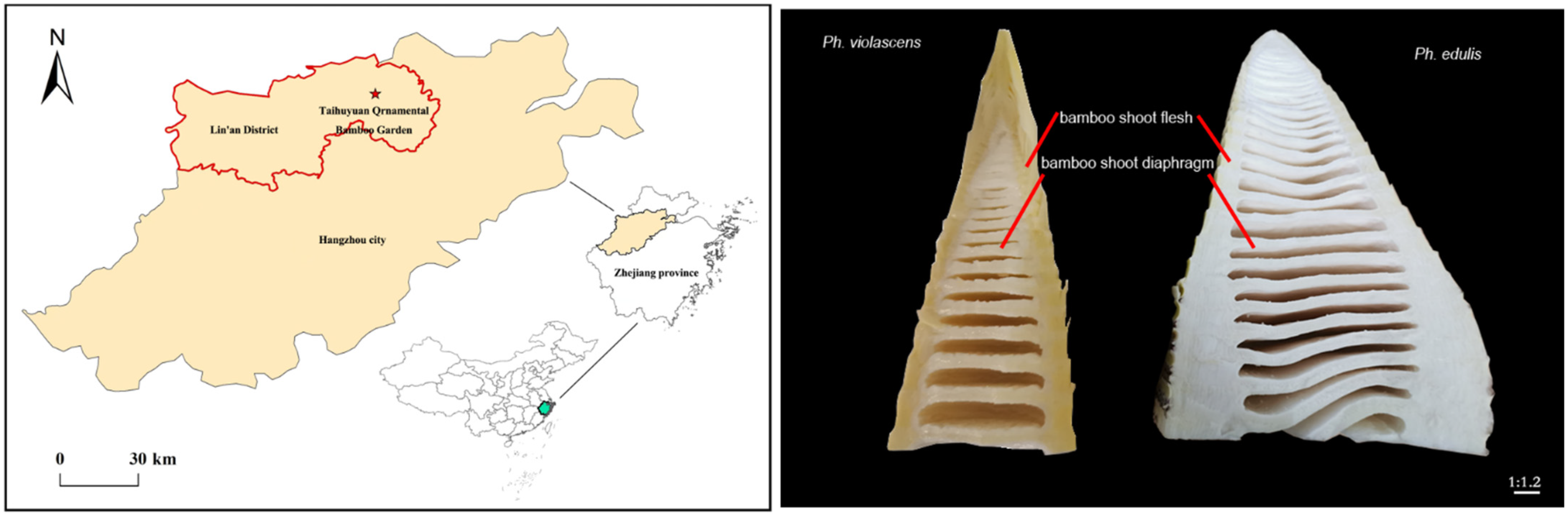
2.1. Study Site
2.2. Sampling Material Description
2.3. Determination of Nutritional Indicators
2.4. Determination of Taste Quality Indicators
2.5. Determination of Free Amino Acids Contents
2.6. Data Analysis
3. Results
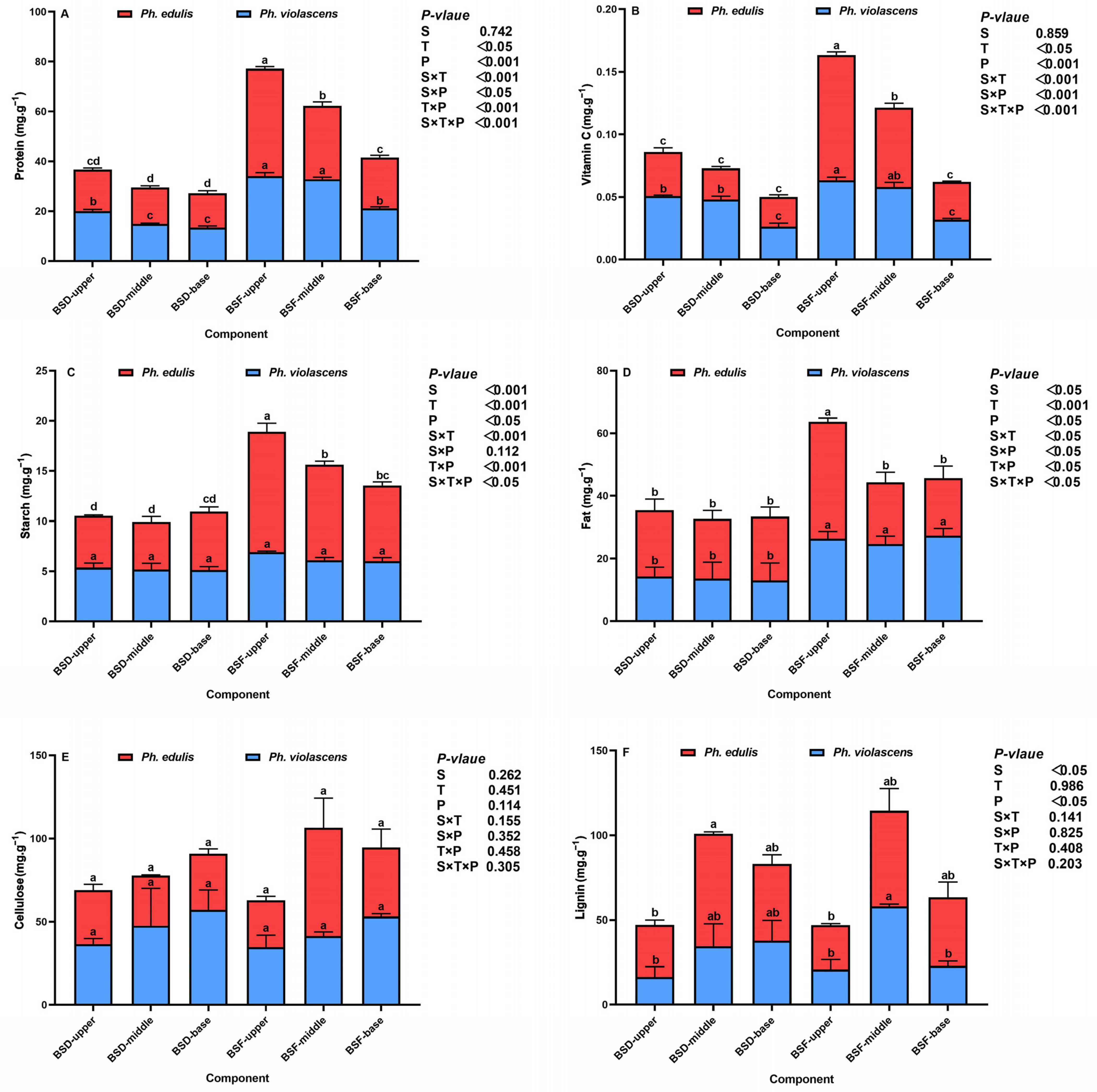
3.1. Shoot Diaphragm (BSD) and Flesh (BSF) Nutritional Status of the Two Bamboo Species
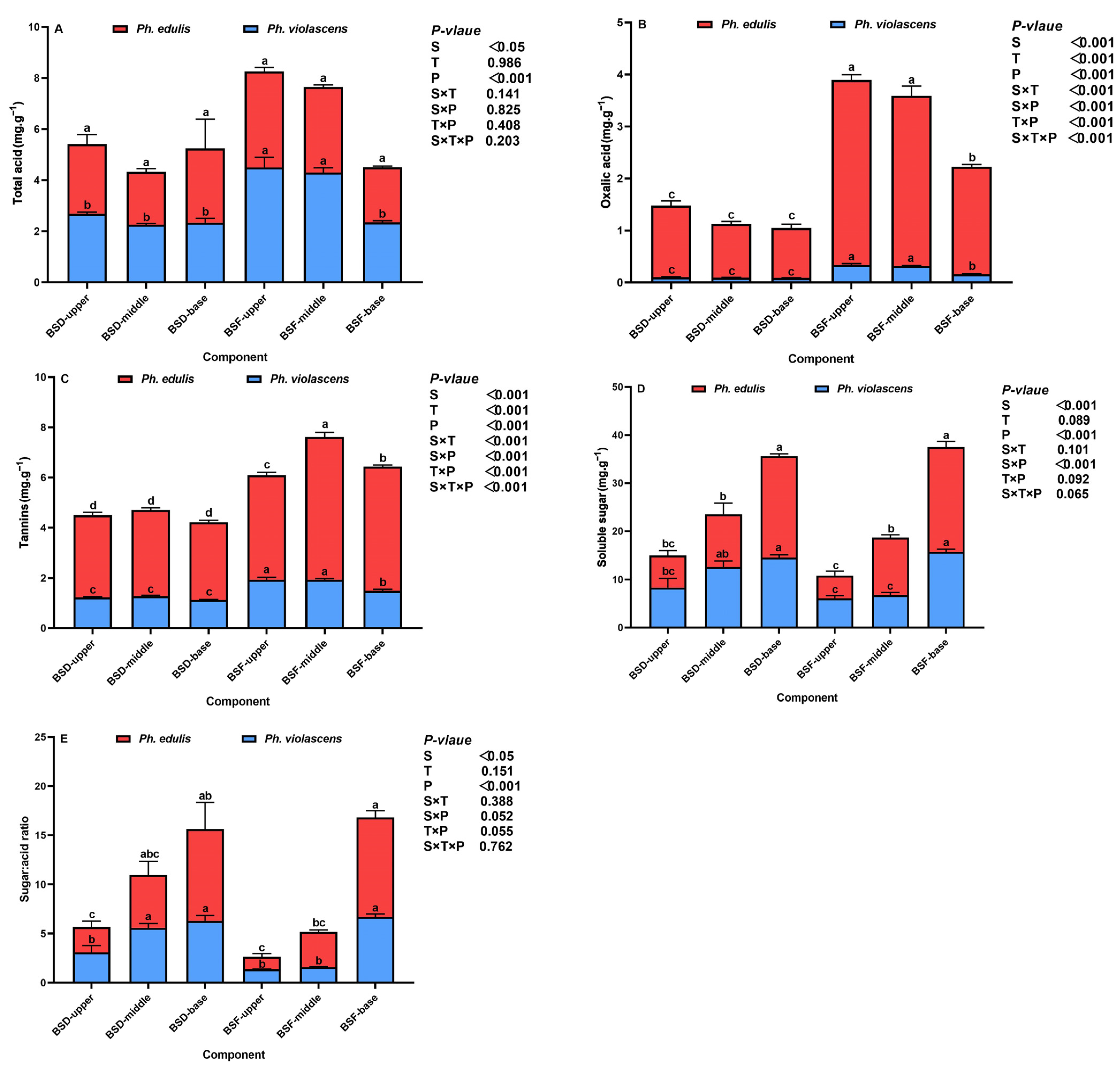
3.2. Shoot Diaphragm (BSD) and Flesh (BSF) Taste Quality of the Two Bamboo Species
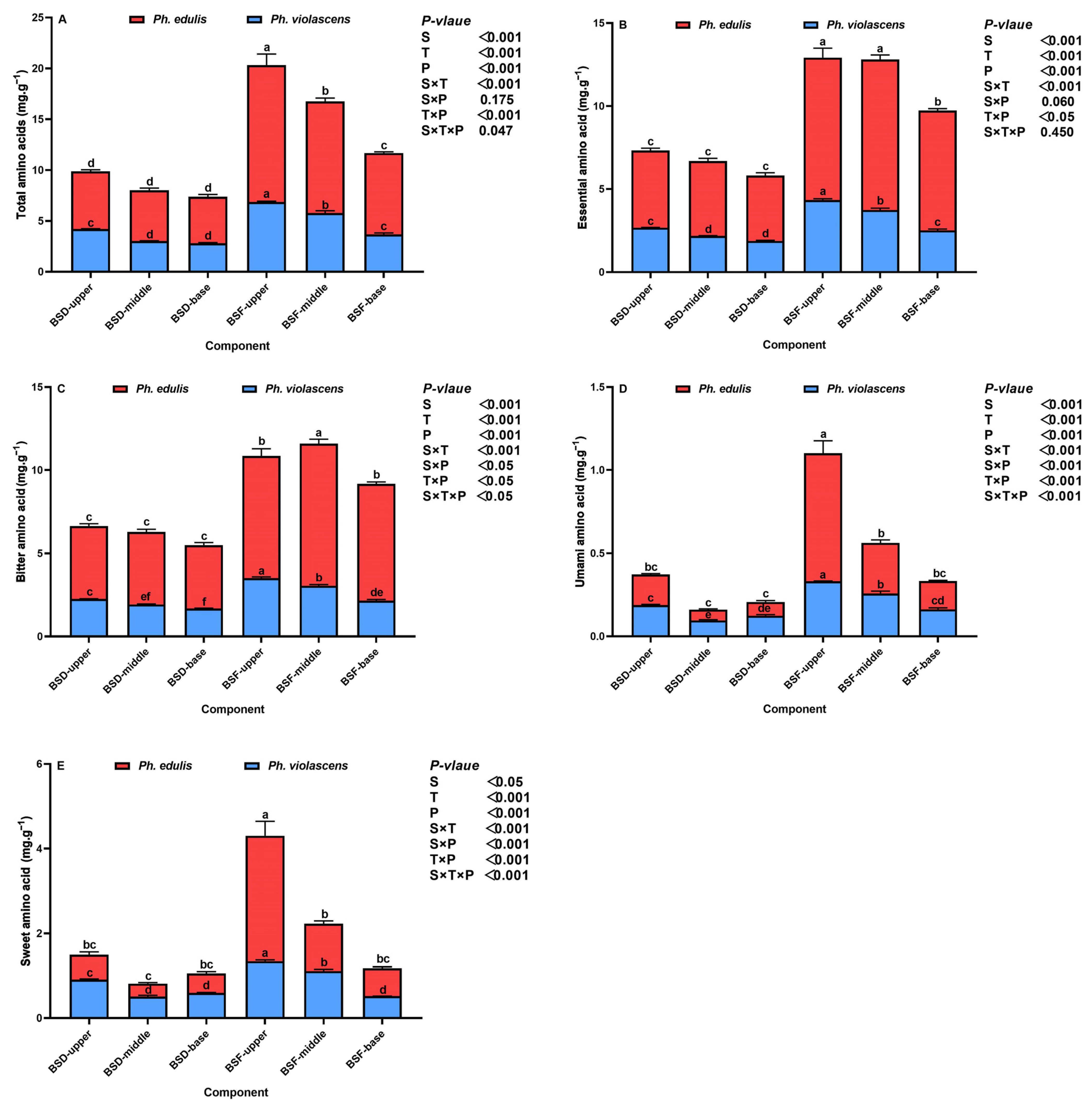
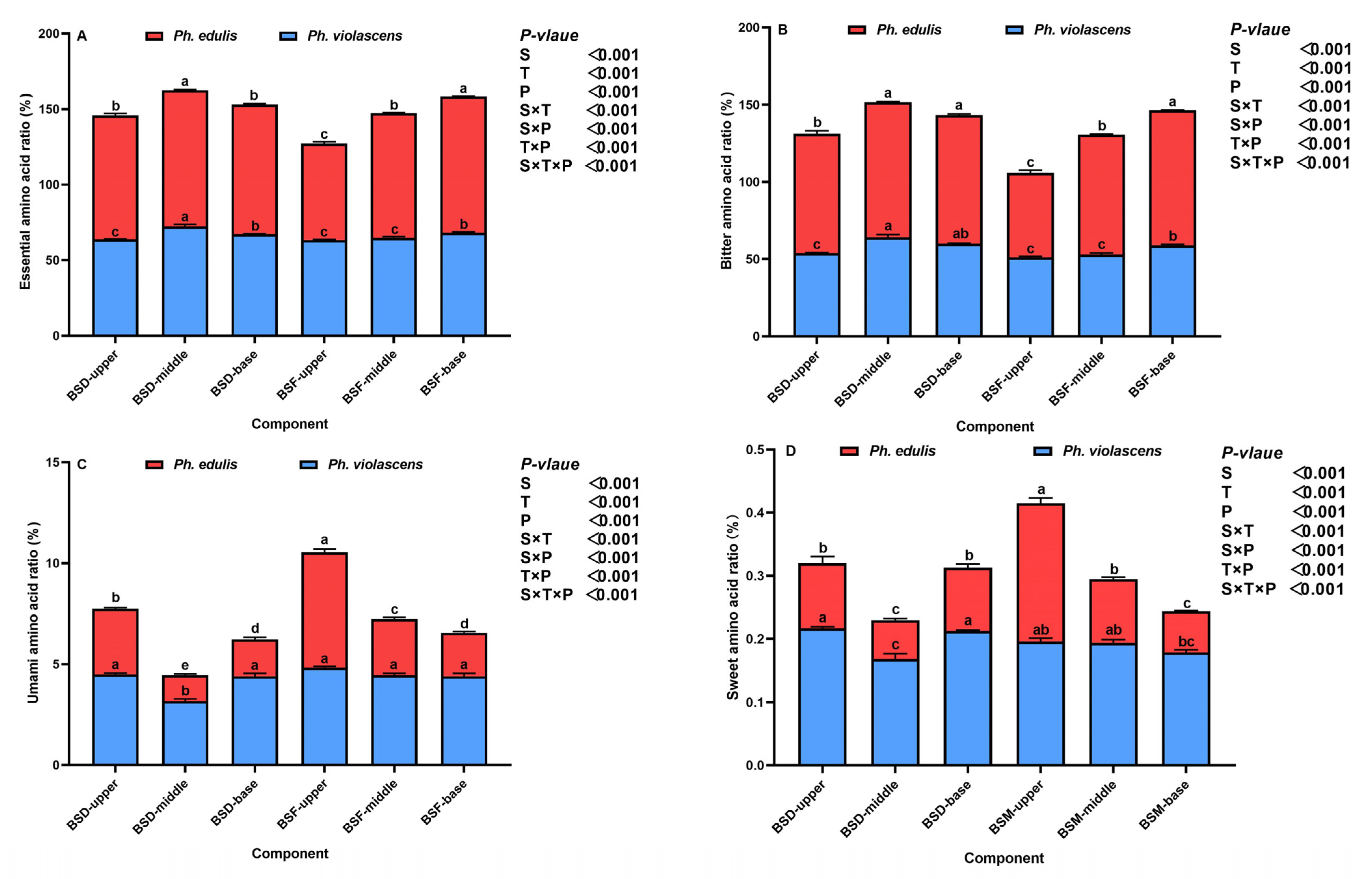
3.3. Amino Acid Composition and the BSD and BSF Ratios of the Two Bamboo Species
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Basumatary, A.; Middha, S.K.; Usha, T.; Basumatary, A.K.; Brahma, B.K.; Goyal, A.K. Bamboo Shoots as a Nutritive Boon for Northeast India: An Overview. 3 Biotech 2017, 7, 169. [Google Scholar] [CrossRef] [PubMed]
- Nongdam, P.; Tikendra, L. The Nutritional Facts of Bamboo Shoots and Their Usage as Important Traditional Foods of Northeast India. Int. Sch. Res. Not. 2014, 2014, 679073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singhal, P.; Bal, L.M.; Satya, S.; Sudhakar, P.; Naik, S.N. Bamboo Shoots: A Novel Source of Nutrition and Medicine. Crit. Rev. Food Sci. Nutr. 2013, 53, 517–534. [Google Scholar] [CrossRef]
- Umemura, M.; Takenaka, C. Retranslocation and Localization of Nutrient Elements in Various Organs of Moso Bamboo (Phyllostachys pubescens). Sci. Total Environ. 2014, 493, 845–853. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-M.; Zhang, S.-C.; Gao, H.-L.; Wang, Q.; Zhou, L.; Zhao, H.-Y.; Li, X.-Y.; Gong, M.; Pan, X.-F.; Cui, C.; et al. Mechanically Robust Bamboo Node and Its Hierarchically Fibrous Structural Design. Natl. Sci. Rev. 2022, 10, nwac195. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.K.; Mondal, S.; Das, S.K.; Ramachandrarao, P. Bamboo—A Functionally Graded Composite-Correlation between Microstructure and Mechanical Strength. J. Mater. Sci. 2005, 40, 5249–5253. [Google Scholar] [CrossRef]
- Taylor, D.; Kinane, B.; Sweeney, C.; Sweetnam, D.; O’Reilly, P.; Duan, K. The Biomechanics of Bamboo: Investigating the Role of the Nodes. Wood Sci. Technol. 2015, 49, 345–357. [Google Scholar] [CrossRef]
- Broadley, M.; Brown, P.; Cakmak, I.; Rengel, Z.; Zhao, F. Function of Nutrients. In Marschner’s Mineral Nutrition of Higher Plants; Elsevier: Amsterdam, The Netherlands, 2012; pp. 191–248. [Google Scholar]
- Wei, X.; Zhou, H.; Chen, F.; Wang, G. Bending Flexibility of Moso Bamboo (Phyllostachys edulis) with Functionally Graded Structure. Materials 2019, 12, 2007. [Google Scholar] [CrossRef] [Green Version]
- Cui, J.; Qin, Z.; Masic, A.; Buehler, M.J. Multiscale Structural Insights of Load Bearing Bamboo: A Computational Modeling Approach. J. Mech. Behav. Biomed. Mater. 2020, 107, 103743. [Google Scholar] [CrossRef]
- Peng, G.; Jiang, Z.; Liu, X.; Fei, B.; Yang, S.; Qin, D.; Ren, H.; Yu, Y.; Xie, H. Detection of Complex Vascular System in Bamboo Node by X-Ray ΜCT Imaging Technique. Holzforschung 2014, 68, 223–227. [Google Scholar] [CrossRef]
- Huang, W.; Yang, Y.; Fu, H.; Ding, Y. Application of Magnetic Resonance Imaging (MRI) Technology in the Characterization of Microstructure and Moisture Content of Young Moso Bamboo. Chin. J. Appl. Ecol. 2022, 34, 1–12. [Google Scholar]
- Liu, S.; Tong, Z.; Tang, Z.; Liu, Y.; Zhang, Z. Bionic Design Modification of Non-Convex Multi-Corner Thin-Walled Columns for Improving Energy Absorption through Adding Bulkheads. Thin-Walled Struct. 2015, 88, 70–81. [Google Scholar] [CrossRef]
- Wang, F.; Yu, Z.; Zhang, M.; Wang, M.; Lu, X.; Liu, X.; Li, Y.; Zhang, X.; Tan, B.; Li, C.; et al. ZmTE1 Promotes Plant Height by Regulating Intercalary Meristem Formation and Internode Cell Elongation in Maize. Plant Biotechnol. J. 2022, 20, 526–537. [Google Scholar] [CrossRef] [PubMed]
- Gamuyao, R.; Nagai, K.; Ayano, M.; Mori, Y.; Minami, A.; Kojima, M.; Suzuki, T.; Sakakibara, H.; Higashiyama, T.; Ashikari, M.; et al. Hormone Distribution and Transcriptome Profiles in Bamboo Shoots Provide Insights on Bamboo Stem Emergence and Growth. Plant Cell Physiol. 2017, 58, 702–716. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Cheng, Z.; Ma, Y.; Bai, Q.; Li, X.; Cao, Z.; Wu, Z.; Gao, J. The Association of Hormone Signalling Genes, Transcription and Changes in Shoot Anatomy during Moso Bamboo Growth. Plant Biotechnol. J. 2018, 16, 72–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.; Gu, R.; Chen, S.; Guo, Z.; Yang, L.; Dong, Y. Interspecific Differences in Morphological Traits of Sheath Leaves and Their Relationships with Taste Quality Indices of Bamboo Shoots. Chin. J. Ecol. 2022, 41, 270–277. [Google Scholar]
- Zhang, J.; Ma, R.; Ding, X.; Huang, M.; Shen, K.; Zhao, S.; Xiao, Z.; Xiu, C. Association among Starch Storage, Metabolism, Related Genes and Growth of Moso Bamboo (Phyllostachys heterocycla) Shoots. BMC Plant Biol. 2021, 21, 477. [Google Scholar] [CrossRef]
- Wang, H.; Zeng, Q.; Cheng, P.; Yu, L.; Kuang, X.; Li, Y. Nutrition Components and Mineral Element Content in 7 Different Cultivated Types (Provenances) of Phyllostachys Violascens Shoots. South China For. Sci. 2019, 47, 36–39. [Google Scholar]
- Wang, S.; Pei, J.; Li, J.; Tang, G.; Zhao, J.; Peng, X.; Nie, S.; Ding, Y.; Wang, C. Sucrose and Starch Metabolism during Fargesia Yunnanensis Shoot Growth. Physiol. Plant 2020, 168, 188–204. [Google Scholar] [CrossRef]
- Guo, Z.; Jiang, Z.; Chen, S.; Xu, B.; Ye, S.; Li, M. Comparative Study on Quality and Palatability of Rhizome Shoot of Phyllostachys Prominens and Phyllostachys edulis. For. Res. 2015, 28, 447–450. [Google Scholar]
- Xu, S.; Chen, S.; Guo, Z.; He, Y.; Yang, L.; Dong, Y.; Xie, Y.; Zhang, J. Effect of Sheath Blade Removal on Phyllostachys Violascens Shoot Quality. Agriculture 2022, 12, 1396. [Google Scholar] [CrossRef]
- Nuzul, M.I.; Jong, V.Y.M.; Koo, L.F.; Chan, T.H.; Ang, C.H.; Idris, J.; Husen, R.; Wong, S.W. Effects of Extraction Methods on Phenolic Content in the Young Bamboo Culm Extracts of Bambusa Beecheyana Munro. Molecules 2022, 27, 2359. [Google Scholar] [CrossRef] [PubMed]
- Leontowicz, H.; Leontowicz, M.; Latocha, P.; Jesion, I.; Park, Y.-S.; Katrich, E.; Barasch, D.; Nemirovski, A.; Gorinstein, S. Bioactivity and Nutritional Properties of Hardy Kiwi Fruit Actinidia Arguta in Comparison with Actinidia Deliciosa ‘Hayward’ and Actinidia Eriantha ‘Bidan’. Food Chem. 2016, 196, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Gao, J. Experimental Guide of Plant Physiology; Higher Education Press: Beijing, China, 2006. [Google Scholar]
- Yu, L.; Peng, X.; Yang, C.; Liu, Y.; Fan, Y. Determination of Oxalic Acid in Plant Tissue and Root Exudate by Reversed Phase High per Formance Liquid Chromatography. Chin. J. Anal. Chem. 2002, 30, 1119–1122. [Google Scholar]
- Dewi, M.A.; Ratnawati, J.; Purwasih, R.W. Determination of Total Tannin of White and Red Rind Pomegranate (Punica granatum L.) by Colorimetry Method Using Reagent 1, 10 Phenantroline. Procedia Chem. 2014, 13, 214–217. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.; Yang, L.; Lin, H.; Chen, S.; Yang, Q. Effects of Altitude on the Variation of Appearance, Nutrition, and Taste for Bamboo Shoots of Pleioblastus Amarus in Shaxian, Fujian Province. Chin. J. Ecol. 2019, 38, 83–88. [Google Scholar]
- Fang, D.; Mei, T.; Röll, A.; Hölscher, D. Water Transfer Between Bamboo Culms in the Period of Sprouting. Front. Plant Sci. 2019, 10, 786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mei, T.; Liu, X.; Fang, D.; Zhou, G.; Ye, C.; Li, P.; Shi, Y.; Du, H.; Berninger, F.; Hölscher, D. Spring Leafing Phenology Favors Younger Culms of Moso Bamboo: Aspects from Water Use Relations. Front. Plant Sci. 2020, 11, 550. [Google Scholar] [CrossRef]
- Mason, M.G.; Ross, J.J.; Babst, B.A.; Wienclaw, B.N.; Beveridge, C.A. Sugar Demand, Not Auxin, Is the Initial Regulator of Apical Dominance. Proc. Natl. Acad. Sci. USA 2014, 111, 6092–6097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, D.K.; Young, J.D. Carbon and Nitrogen Provisions Alter the Metabolic Flux in Developing Soybean Embryos. Plant Physiol. 2013, 161, 1458–1475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbier, F.; Péron, T.; Lecerf, M.; Perez-Garcia, M.-D.; Barrière, Q.; Rolčík, J.; Boutet-Mercey, S.; Citerne, S.; Lemoine, R.; Porcheron, B.; et al. Sucrose Is an Early Modulator of the Key Hormonal Mechanisms Controlling Bud Outgrowth in Rosa Hybrida. J. Exp. Bot. 2015, 66, 2569–2582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Ying, Y.; Wang, J.; Zhao, X.; Zeng, W.; Beahan, C.; He, J.; Chen, X.; Bacic, A.; Song, L.; et al. Transcriptome Analysis Provides Insights into Xylogenesis Formation in Moso Bamboo (Phyllostachys edulis) Shoot. Sci. Rep. 2018, 8, 3951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waldron, K.W.; Selvendran, R.R. Effect of Maturation and Storage on Asparagus (Asparagus officinalis) Cell Wall Composition. Physiol. Plant 1990, 80, 576–583. [Google Scholar] [CrossRef]
- Femenia, A.; Waldron, K.W.; Robertson, J.A.; Selvendran, R.R. Compositional and Structural Modification of the Cell Wall of Cauliflower (Brassica oleracea L. Var Botrytis) during Tissue Development and Plant Maturation. Carbohydr. Polym. 1999, 39, 101–108. [Google Scholar] [CrossRef]
- Xu, Y.G.; Lu, S.M.; Wang, Q. Changes of Cell Wall Components and PAL Activity in Fresh-Cut Bamboo Shoots during Cold Storage. J. Food Biochem. 2004, 28, 169–177. [Google Scholar] [CrossRef]
- Zhu, Y.; Huang, J.; Wang, K.; Wang, B.; Sun, S.; Lin, X.; Song, L.; Wu, A.; Li, H. Characterization of Lignin Structures in Phyllostachys edulis (Moso bamboo) at Different Ages. Polymers 2020, 12, 187. [Google Scholar] [CrossRef] [Green Version]
- Besada, C.; Salvador, A.; Arnal, L.; Martínez-Jávega, J.M. Optimization of the Duration of Deastringency Treatment Depending on Persimmon Maturity. Acta Hortic. 2010, 858, 69–74. [Google Scholar] [CrossRef]
- Ghasemzadeh, A.; Jaafar, H. Effect of CO2 Enrichment on Synthesis of Some Primary and Secondary Metabolites in Ginger (Zingiber officinale Roscoe). Int. J. Mol. Sci. 2011, 12, 1101–1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Ding, X.; Zhang, S.; Zhang, Z.; Cai, H.; Zhang, Y. The Distributions of Bitter and Astringent Taste Compounds in the Bamboo Shoot of Dendrocalamus Latiflorus under Different Light Intensities. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2015, 39, 161–166. [Google Scholar]
- Li, R.; Dao, D.; Xiang, M.; Sun, A.; Yang, Q. Analysis on Nutrient Quantity in Different Parts of Dendrocalamus Hamilonii Shoots. J. For. Eng. 2010, 24, 76–78. [Google Scholar]
- Gao, G.; Zhong, H.; Pan, Y.; Wu, L.; Wu, Z.; Wen, X. Effects of Ecological Factors on Biomass Allocation of Indocalamus Decorus Pot. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2017, 41, 35–41. [Google Scholar]
- Chongtham, N.; Bisht, M.S.; Premlata, T.; Bajwa, H.K.; Sharma, V.; Santosh, O. Quality Improvement of Bamboo Shoots by Removal of Antinutrients Using Different Processing Techniques: A Review. J. Food Sci. Technol. 2022, 59, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ju, F.; Liu, S.; Zhang, S.; Ma, H.; Chen, J.; Ge, C.; Shen, Q.; Zhang, X.; Zhao, X.; Zhang, Y.; et al. Transcriptome Analysis and Identification of Genes Associated with Fruiting Branch Internode Elongation in Upland Cotton. BMC Plant Biol. 2019, 19, 415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkes, R.A.; Waldbauer, J.; Aristilde, L. Analogous Metabolic Decoupling in Pseudomonas Putida and Comamonas Testosteroni Implies Energetic Bypass to Facilitate Gluconeogenic Growth. mBio 2021, 12, e03259-21. [Google Scholar] [CrossRef]
- Christian, A.L.; Knott, K.K.; Vance, C.K.; Falcone, J.F.; Bauer, L.L.; Fahey, G.C.; Willard, S.; Kouba, A.J. Nutrient and Mineral Composition during Shoot Growth in Seven Species of Phyllostachys and Pseudosasa Bamboo Consumed by Giant Panda. J. Anim. Physiol. Anim. Nutr. 2015, 99, 1172–1183. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, L.; Hu, J.; Guo, Z.; Chen, S.; He, Q. Shoot Nutrition and Flavor Variation in Two Phyllostachys Species: Does the Quality of Edible Bamboo Shoot Diaphragm and Flesh Differ? Foods 2023, 12, 1180. https://doi.org/10.3390/foods12061180
Fan L, Hu J, Guo Z, Chen S, He Q. Shoot Nutrition and Flavor Variation in Two Phyllostachys Species: Does the Quality of Edible Bamboo Shoot Diaphragm and Flesh Differ? Foods. 2023; 12(6):1180. https://doi.org/10.3390/foods12061180
Chicago/Turabian StyleFan, Lili, Junjing Hu, Ziwu Guo, Shuanglin Chen, and Qijiang He. 2023. "Shoot Nutrition and Flavor Variation in Two Phyllostachys Species: Does the Quality of Edible Bamboo Shoot Diaphragm and Flesh Differ?" Foods 12, no. 6: 1180. https://doi.org/10.3390/foods12061180
APA StyleFan, L., Hu, J., Guo, Z., Chen, S., & He, Q. (2023). Shoot Nutrition and Flavor Variation in Two Phyllostachys Species: Does the Quality of Edible Bamboo Shoot Diaphragm and Flesh Differ? Foods, 12(6), 1180. https://doi.org/10.3390/foods12061180