

Comparative Responses of Silicon to Reduce Cadmium and Enrich Selenium in Rice Varieties


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Material
2.2. Nutrient Treatments Used in the Experiment
2.3. Tested Soil
2.4. Pot Experiment Design
2.5. Determination Method
2.5.1. Agricultural Traits
2.5.2. Determination of Non-Protein Thiol (NPT) Content
2.5.3. Preparation of Samples from Various Parts of the Rice
2.5.4. Determination of Cd Content in Various Parts of the Rice
2.5.5. Determination of Se Content in Various Parts of the Rice
2.5.6. Determination of Se and Cd in Soil Samples
2.5.7. Inorganic Se Measurement Method
2.5.8. Organic Se Measurement Method
2.6. Data Analysis
3. Results
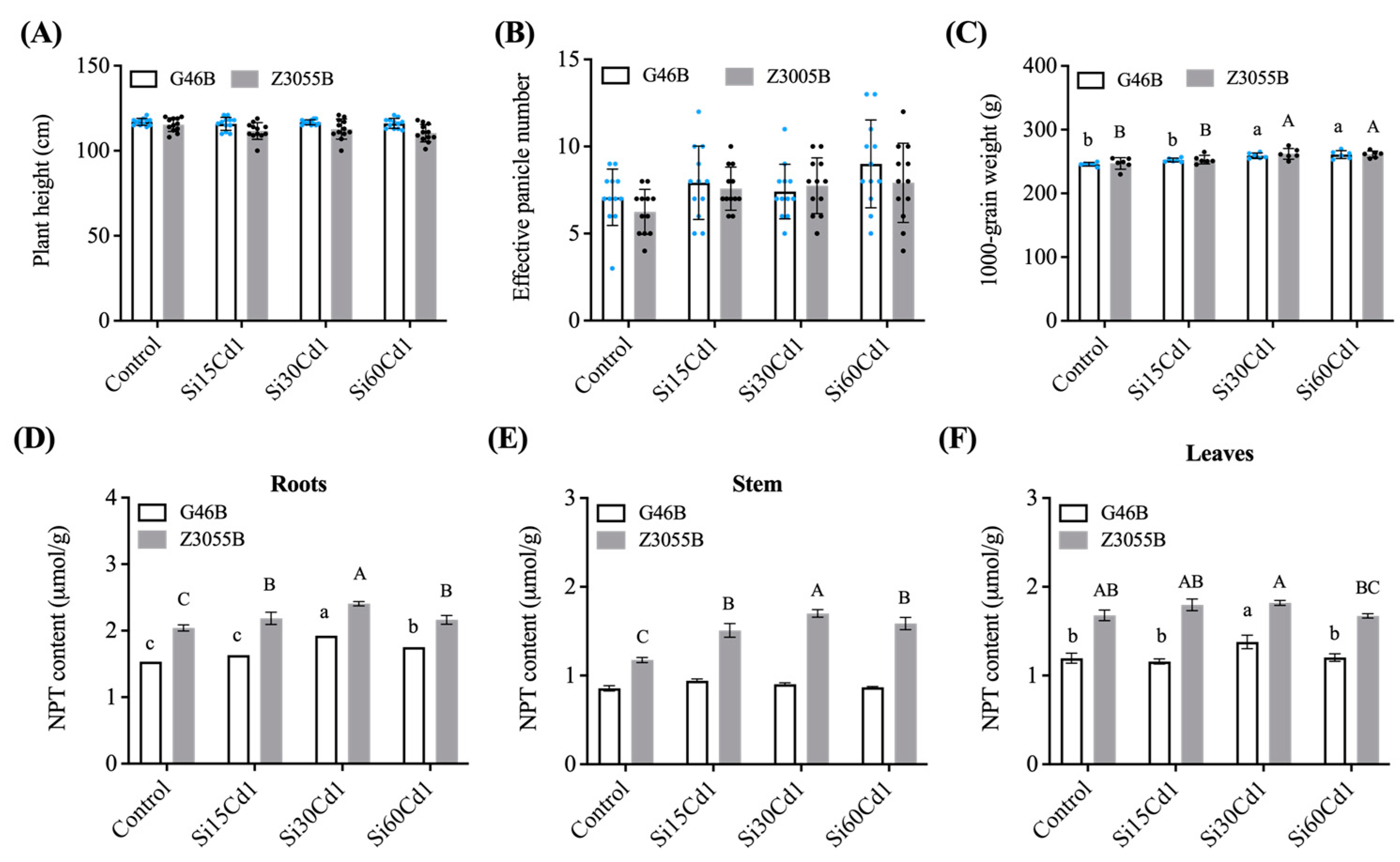
3.1. Agronomic Traits
3.2. NPT Content
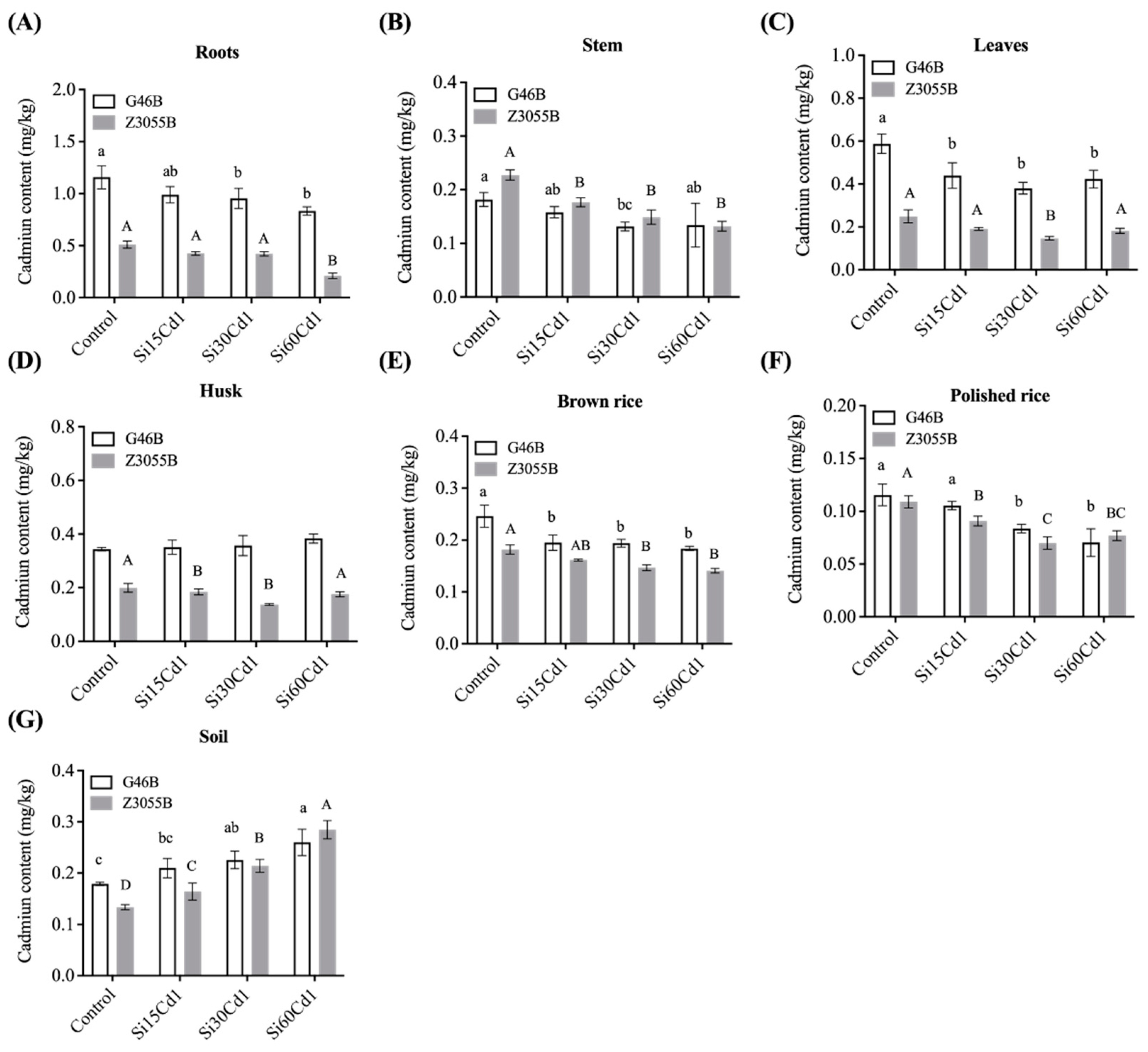
3.3. Cd Concentration in Different Parts of Plants of Two Different Genotypes of Rice
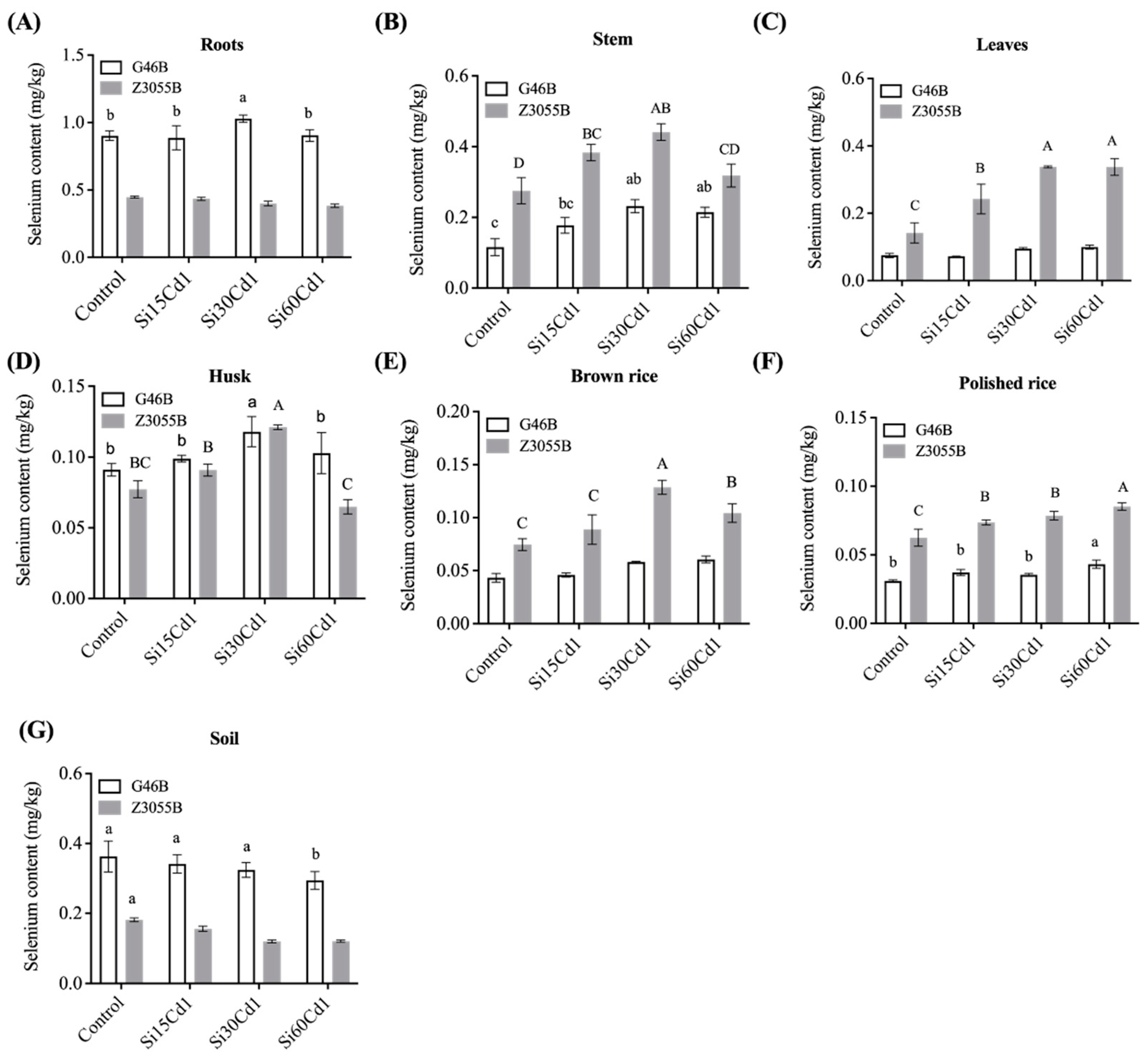
3.4. Se Concentration in Different Parts of Two Different Genotypes of Rice
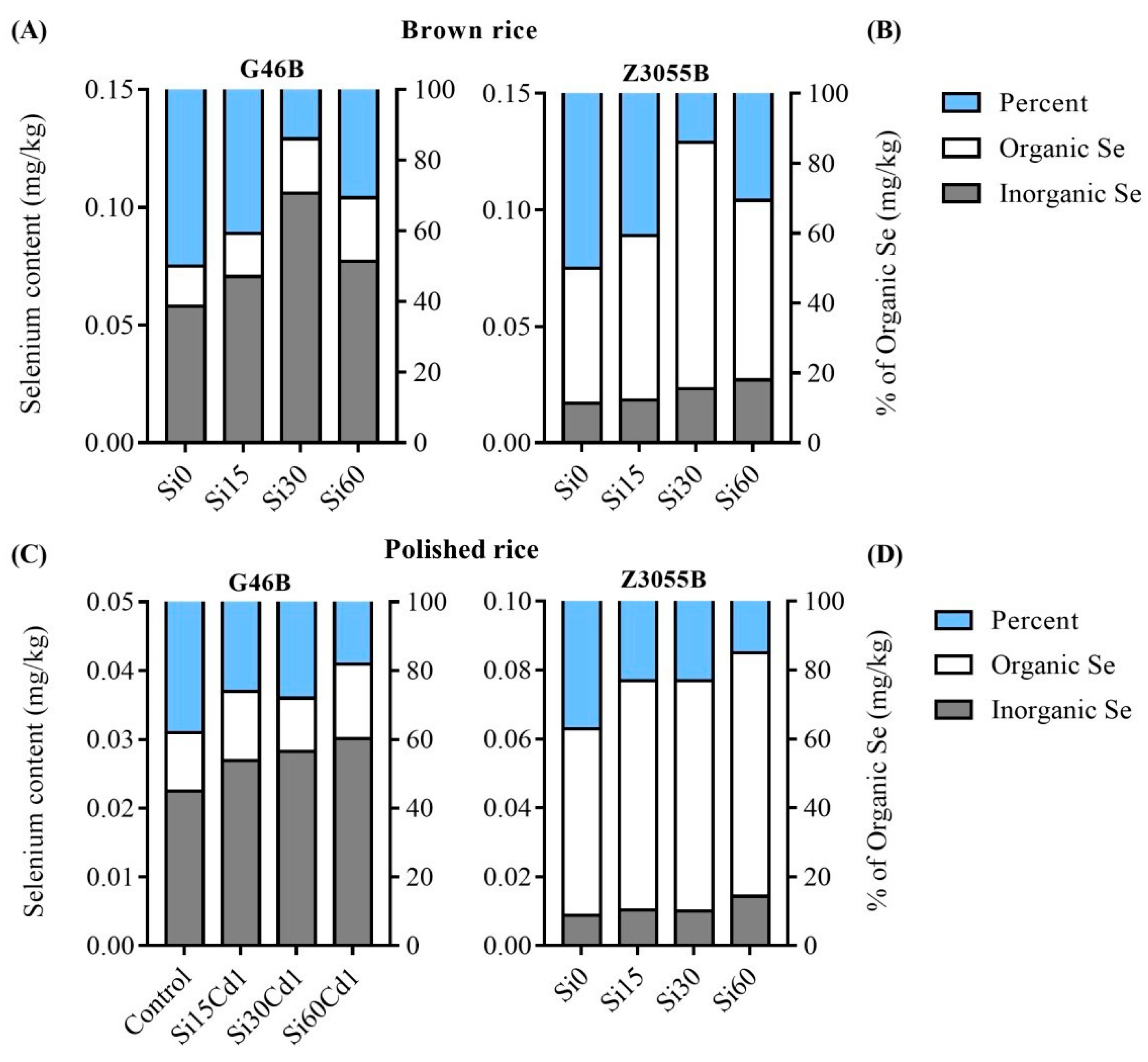
3.5. Distribution of Inorganic Se and Organic Se in Brown Rice of Two Genotypes
3.6. Distribution of Inorganic Se and Organic Se in Polished Rice of Two Genotypes of Rice
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lim, J.E.; Ahmad, M.; Usman, A.; Sang, S.L.; Jeon, W.T.; Oh, S.E.; Yang, J.E.; Yong, S.O. Effects of natural and calcined poultry waste on Cd, Pb and As mobility in contaminated soil. Environ. Earth Sci. 2013, 69, 11–20. [Google Scholar] [CrossRef]
- Mi, Y.; Cheng, M.; Yu, Q.; Si, Y. Foliar application of anthocyanin extract regulates cadmium accumulation and distribution in rice (Oryza sativa L.) at tillering and booting stages. Ecotoxicol. Environ. Saf. 2021, 224, 112647. [Google Scholar] [CrossRef]
- Sardar, K.; Ali, S.; Hameed, S.; Afzal, S.; Fatima, S.; Shakoor, M.B.; Bharwana, S.A.; Tauqeer, H.M. Heavy metals contamination and what are the impacts on living organisms. Green. J. Environ. Manag. Public Saf. 2013, 2, 172–179. [Google Scholar]
- Moulis, J.M. Cellular mechanisms of cadmium toxicity related to the homeostasis of essential metals. Biometals 2010, 23, 877–896. [Google Scholar] [CrossRef]
- El-Boshy, M.E.; Risha, E.F.; Abdelhamid, F.M.; Mubarak, M.S.; Hadda, T.B. Protective effects of selenium against cadmium induced hematological disturbances, immunosuppressive, oxidative stress and hepatorenal damage in rats. J. Trace Elem. Med. Biol. 2015, 29, 104–110. [Google Scholar] [CrossRef]
- Mozafariyan, M.; Shekari, L.; Hawrylak-Nowak, B.; Kamelmanesh, M.M. Protective Role of Selenium on Pepper Exposed to Cadmium Stress During Reproductive Stage. Biol. Trace Elem. Res. 2014, 160, 97–107. [Google Scholar] [CrossRef]
- Tang, X.; Li, Q.; Wu, M.; Lin, L.; Scholz, M. Review of remediation practices regarding cadmium-enriched farmland soil with particular reference to China. J. Environ. Manag. 2016, 181, 646–662. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Xu, J.; Huang, Q.; Tang, S.; Li, Y.; Li, F.; Zhou, D.; Wu, Z. Some deliberations on the issues of heavy metals in farmlands of China. Acta Pedol. Sin. 2013, 50, 186–194. [Google Scholar]
- Meharg, C.; Meharg, A.A. Silicon, the silver bullet for mitigating biotic and abiotic stress, and improving grain quality, in rice? Environ. Exp. Bot. 2015, 120, 8–17. [Google Scholar] [CrossRef] [Green Version]
- Borlaug, N. Feeding a Hungry World. Science 2007, 318, 359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.; Zeng, M.; Zhou, X.; Liao, B.-H.; Peng, P.-Q.; Hu, M.; Zhu, W.; Wu, Y.-J.; Zou, Z.-J. Heavy metal translocation and accumulation in iron plaques and plant tissues for 32 hybrid rice (Oryza sativa L.) cultivars. Plant Soil 2015, 386, 317–329. [Google Scholar] [CrossRef]
- Liu, C.; Li, F.; Luo, C.; Liu, X.; Wang, S.; Liu, T.; Li, X. Foliar application of two silica sols reduced cadmium accumulation in rice grains. J. Hazard. Mater. 2009, 161, 1466–1472. [Google Scholar] [CrossRef]
- Epstein, E. Silicon: Its manifold roles in plants. Ann. Appl. Biol. 2009, 155, 155–160. [Google Scholar] [CrossRef]
- Dallagnol, L.; Rodrigues, F.; Chaves, A.D.M.; Vale, F.; DaMatta, F. Photosynthesis and sugar concentration are impaired by the defective active silicon uptake in rice plants infected with Bipolaris oryzae. Plant Pathol. 2013, 62, 120–129. [Google Scholar] [CrossRef] [Green Version]
- Pan, D.; Liu, C.; Yi, J.; Li, X.; Li, F. Different effects of foliar application of silica sol on arsenic translocation in rice under low and high arsenite stress. J. Environ. Sci. 2021, 105, 22–32. [Google Scholar] [CrossRef]
- Shi, Q.; Bao, Z.; Zhu, Z.; Yong, H.; Qian, Q.; Yu, J. Silicon-mediated alleviation of Mn toxicity in Cucumis sativus in relation to activities of superoxide dismutase and ascorbate peroxidase. Phytochemistry 2005, 66, 1551–1559. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Zhou, J.; Liu, H.; Zhang, W.; Hu, Y.; Liang, J.; Zhou, J. Foliar spraying with silicon and selenium reduces cadmium uptake and mitigates cadmium toxicity in rice. Sci. Total Environ. 2018, 631–632, 1100–1108. [Google Scholar] [CrossRef] [PubMed]
- Schiavon, M.; Pilon-Smits, E.A.H. Selenium Biofortification and Phytoremediation Phytotechnologies: A Review. J. Environ. Qual. 2016, 46, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farooq, M.U.; Tang, Z.; Zeng, R.; Liang, Y.; Zhang, Y.; Zheng, T.; Ei, H.H.; Ye, X.; Jia, X.; Zhu, J. Accumulation, mobilization, and transformation of selenium in rice grain provided with foliar sodium selenite. J. Sci. Food Agric. 2019, 99, 2892–2900. [Google Scholar] [CrossRef]
- Liang, Y.; Farooq, M.U.; Rui, Z.; Tang, Z.; Zhu, J. Breeding of Selenium Rich Red Glutinous Rice, Protein Extraction and Analysis of the Distribution of Selenium in Grain. Int. J. Agric. Biol. 2018, 20, 1005–1011. [Google Scholar]
- Zeng, R.; Farooq, M.U.; Zhang, G.; Tang, Z.; Zheng, T.; Su, Y.; Hussain, S.; Liang, Y.; Ye, X.; Jia, X. Dissecting the potential of selenoproteins extracted from selenium-enriched rice on physiological, biochemical and anti-ageing effects in vivo. Biol. Trace Elem. Res. 2020, 196, 119–130. [Google Scholar] [CrossRef]
- Farooq, M.U.; Tang, Z.; Zheng, T.; Asghar, M.A.; Zeng, R.; Su, Y.; Ei, H.H.; Liang, Y.; Zhang, Y.; Ye, X. Cross-talk between cadmium and selenium at elevated cadmium stress determines the fate of selenium uptake in rice. Biomolecules 2019, 9, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ei, H.H.; Zheng, T.; Farooq, M.U.; Zeng, R.; Su, Y.; Huang, X.; Zhang, Y.; Liang, Y.; Tang, Z.; Ye, X. Evaluation on zinc and selenium nutrients in polished rice of rice genotypes under zinc biofortification. Biomed. J. Sci. Tech. Res. 2019, 21, 16205–16213. [Google Scholar]
- Liang, Y.; Farooq, M.U.; Hu, Y.; Tang, Z.; Zhang, Y.; Zeng, R.; Zheng, T.; Ei, H.H.; Ye, X.; Jia, X. Study on Stability and Antioxidant Activity of Red Anthocyanidin Glucoside Rich Hybrid Rice, its Nutritional and Physicochemical Characteristics. Food Sci. Technol. Res. 2018, 24, 687–696. [Google Scholar] [CrossRef] [Green Version]
- Saidi, I.; Chtourou, Y.; Djebali, W. Selenium alleviates cadmium toxicity by preventing oxidative stress in sunflower (Helianthus annuus) seedlings. J. Plant Physiol. 2014, 171, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Li, L.; Farooq, M.U.; Huang, X.; Zheng, T.; Zhang, Y.J.; Ei, H.H.; Panhwar, F.H.; Tang, Z.; Zeng, R.; et al. Rescue effects of Se-enriched rice on physiological and biochemical characteristics in cadmium poisoning mice. Environ. Sci. Pollut. Res. 2021, 28, 20023–20033. [Google Scholar] [CrossRef] [PubMed]
- Sharma, J.; Chakraverty, N. Mechanism of Plant Tolerance in Response to Heavy Metals. In Molecular Stress Physiology of Plants; Springer: New Delhi, India, 2013; pp. 289–308. [Google Scholar]
- Fodor, E.; Szabó-Nagy, A.; Erdei, L. The effects of cadmium on the fluidity and H+-ATPase activity of plasma membrane from sunflower and wheat roots. J. Plant Physiol. 1995, 147, 87–92. [Google Scholar] [CrossRef]
- Zhou, Z.; Zhang, B.; Liu, H.; Liang, X.; Ma, W.; Shi, Z.; Yang, S. Zinc effects on cadmium toxicity in two wheat varieties (Triticum aestivum L.) differing in grain cadmium accumulation. Ecotoxicol. Environ. Saf. 2019, 183, 109562. [Google Scholar] [CrossRef]
- Clemens, S.; Aarts, M.G.M.; Thomine, S.; Verbruggen, N. Plant science: The key to preventing slow cadmium poisoning. Trends Plant Sci. 2013, 18, 92–99. [Google Scholar] [CrossRef]
- Yan, B.F.; Nguyen, C.; Pokrovsky, O.S.; Candaudap, F.; Cornu, J.Y. Cadmium allocation to grains in durum wheat exposed to low Cd concentrations in hydroponics. Ecotoxicol. Environ. Saf. 2019, 184, 109592. [Google Scholar] [CrossRef]
- Ma, J.; Cai, H.; He, C.; Zhang, W.; Wang, L. A hemicellulose-bound form of silicon inhibits cadmium ion uptake in rice (Oryza sativa) cells. New Phytol. 2015, 206, 1063–1074. [Google Scholar] [CrossRef] [PubMed]
- Shobbar, Z.S.; Oane, R.; Gamuyao, R.; De Palma, J.; Malboobi, M.A.; Karimzadeh, G.; Javaran, M.J.; Bennett, J. Abscisic acid regulates gene expression in cortical fiber cells and silica cells of rice shoots. New Phytol. 2008, 178, 68–79. [Google Scholar] [CrossRef]
- Jozefczak, M.; Remans, T.; Vangronsveld, J.; Cuypers, A. Glutathione is a key player in metal-induced oxidative stress defenses. Int. J. Mol. Sci. 2012, 13, 3145–3175. [Google Scholar] [CrossRef] [Green Version]
- Weng, B.; Xie, X.; Weiss, D.J.; Liu, J.; Lu, H.; Yan, C. Kandelia obovata (S., L.) Yong tolerance mechanisms to Cadmium: Subcellular distribution, chemical forms and thiol pools. Mar. Pollut. Bull. 2012, 64, 2453–2460. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, J.; Ma, Y.; Cai, M.; Gao, F. Effects of silicon on cadmium accumulation and non-protein thiol content in the seedlings of two rice varieties under cadmium stress. J. Agro-Environ. Sci. 2018, 37, 1066–1071. [Google Scholar]
- Li, Q.; Guo, J.; Zhang, X.; Yu, H.; Huang, F.; Zhang, L.; Zhang, M.; Li, T. Changes of non-protein thiols in roots and organic acids in xylem sap involved in cadmium translocation of cadmium-safe rice line (Oryza Sative L.). Plant Soil 2019, 439, 475–486. [Google Scholar] [CrossRef]
- Wu, Z.; Wang, F.; Liu, S.; Du, Y.; Li, F.; Du, R.; Wen, D.; Zhao, J. Comparative responses to silicon and selenium in relation to cadmium uptake, compartmentation in roots, and xylem transport in flowering Chinese cabbage (Brassica campestris L. ssp. chinensis var. utilis) under cadmium stress. Environ. Exp. Bot. 2016, 131, 173–180. [Google Scholar] [CrossRef]
- Ishikawa, S.; Lshimaru, Y.; Igura, M.; Kuramata, M.; Abe, T.; Senoura, T.; Hase, Y.; Arao, T.; Nishizawa, N.K.; Nakanishi, H. Ion-beam irradiation, gene identification, and marker-assisted breeding in the development of low-cadmium rice. Proc. Natl. Acad. Sci. USA 2012, 109, 19166–19171. [Google Scholar] [CrossRef] [Green Version]
- Qian, Y.; Gallagher, F.J.; Feng, H.; Wu, M.; Zhu, Q. Vanadium uptake and translocation in dominant plant species on an urban coastal brownfield site. Sci. Total Environ. 2014, 476–477, 696–704. [Google Scholar] [CrossRef]
- Rizwan, M.; Meunier, J.D.; Miche, H.; Keller, C. Effect of silicon on reducing cadmium toxicity in durum wheat (Triticum turgidum L. cv. Claudio W.) grown in a soil with aged contamination. J. Hazard. Mater. 2012, 209–210, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Jie, M.; He, C.; Li, X.; Zhang, W.; Xu, F.; Lin, Y.; Wang, L. Inhibition of cadmium ion uptake in rice (Oryza sativa) cells by a wall-bound form of silicon. New Phytol. 2013, 200, 691–699. [Google Scholar] [CrossRef]
- Chen, Y.P.; Peng, Q.; Liang, D.L.; Song, W.W.; Yu, D.S. Effects of Nitrogen Application on Selenium Uptake, Translocation and Distribution in Winter Wheat. Huan Jing Ke Xue Huanjing Kexue 2017, 38, 825–831. [Google Scholar] [PubMed]
- Liang, Y.; Su, Y.; Li, L.; Huang, X.; Panhwar, F.H.; Zheng, T.; Tang, Z.; Ei, H.H.; Farooq, M.U.; Zeng, R. Quick selenium accumulation in the selenium-rich rice and its physiological responses in changing selenium environments. BMC Plant Biol. 2019, 19, 559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tobiasz, A.; Walas, S.; Filek, M.; Mrowiec, H.; Samsel, K.; Sieprawska, A.; Hartikainen, H. Effect of selenium on distribution of macro- and micro-elements to different tissues during wheat ontogeny. Biol. Plant. 2014, 58, 370–374. [Google Scholar] [CrossRef]
- Ueno, D.; Koyama, E.; Yamaji, N.; Ma, J.F. Physiological, genetic, and molecular characterization of a high-Cd-accumulating rice cultivar, Jarjan. J. Exp. Bot. 2011, 62, 2265–2272. [Google Scholar] [CrossRef] [Green Version]
- Filek, M.; Zembala, M.; Korna, A.; Walas, S.; Mrowiec, H.; Hartikainen, H. The uptake and translocation of macro- and microelements in rape and wheat seedlings as affected by selenium supply level. Plant Soil 2010, 336, 303–312. [Google Scholar] [CrossRef]
- Zhou, X.; Shi, W.; Yang, L. Genotypical differences and characteristics of Se uptake and accumulation in rice. Soils 2007, 39, 731–736. [Google Scholar]
- Zhang, L.; Hu, B.; Deng, K.; Gao, X.; Chu, C. NRT1.1B improves selenium concentrations in rice grains by facilitating selenomethinone translocation. Plant Biotechnol. J. 2019, 17, 1058–1068. [Google Scholar] [CrossRef] [Green Version]
- Li, H.F.; Lombi, E.; Stroud, J.L.; Mcgrath, S.P.; Zhao, F.J. Selenium Speciation in Soil and Rice: Influence of Water Management and Se Fertilization. J. Agric. Food Chem. 2010, 58, 11837–11843. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Content | Method |
|---|---|---|
| pH | 5.93 ± 0.066 | NY/T 1377-2007 |
| Organic matter (g/kg of soil) | 28.89 ± 2.233 | NY/T 1121.6-2006 |
| Total nitrogen (g/kg of soil) | 0.191 ± 0.012 | HJ 717-2014 |
| Total phosphorus (g/kg of soil) | 1.49 ± 0.015 | NY/T 88-1988 |
| Total potassium (g/kg of soil) | 10.3 ± 0.931 | NY/T 87-1988 |
| Total selenium (mg/kg of soil) | 0.22 ± 0.0298 | GB 5009.93-2017 |
| Total Cadmium (mg/kg of soil) | 0.085 ± 0.008 | GB/T17141-1997 |
| Interaction Effects | Plant Height (cm) | Effective Panicle Number | 1000-Grain Weight (g) | NPT Content | |||
|---|---|---|---|---|---|---|---|
| Roots (µmol/g) | Stem (µmol/g) | Leaves (µmol/g) | |||||
| Treatment (T) | Control | 116.157 | 6.667b | 246.379b | 1.788c | 1.016c | 1.437b |
| Si15Cd1 | 113.735 | 7.75ab | 252.697b | 1.909b | 1.225b | 1.478b | |
| Si30Cd1 | 114.75 | 7.578ab | 260.673a | 2.165a | 1.301a | 1.598a | |
| Si60Cd1 | 113.25 | 8.458a | 261.133a | 1.957b | 1.227b | 1.438b | |
| SEm± | 0.828 | 0.374 | 1.776 | 0.022 | 0.018 | 0.021 | |
| Genotypes | G46B | 116.417 | 7.854a | 254.565 | 1.71b | 0.892b | 1.234b |
| Z3055B | 112.542 | 7.375b | 255.867 | 2.199a | 1.493a | 1.742a | |
| SEm± | 0.585 | 0.264 | 1.256 | 0.016 | 0.013 | 0.015 | |
| Interaction | T | ns | * | ** | ** | ** | ** |
| G | ** | ns | ns | ** | ** | ** | |
| T × G | ns | ns | ns | ns | ** | * | |
| Interaction Effects | Cadmium Concentration (mg/kg) | |||||||
|---|---|---|---|---|---|---|---|---|
| Soil | Roots | Stem | Leaves | Husk | Brown Rice | Polished Rice | ||
| Treatment (T) | Control | 0.156d | 0.835a | 0.205a | 0.419a | 0.272a | 0.214a | 0.112a |
| Si15Cd1 | 0.187c | 0.708b | 0.167b | 0.316b | 0.268ab | 0.178bc | 0.098 | |
| Si30Cd1 | 0.22b | 0.689b | 0.140c | 0.264bc | 0.247ab | 0.170bc | 0.77c | |
| Si60Cd1 | 0.272a | 0.523c | 0.133c | 0.303c | 0.280b | 0.162c | 0.077c | |
| SEm± | 0.007 | 0.026 | 0.007 | 0.014 | 0.008 | 0.04 | 0.003 | |
| Genotypes (G) | G46B | 0.219a | 0.984a | 0.151b | 0.458a | 0.359a | 0.205a | 0.097a |
| Z3055B | 0.199b | 0.393b | 0.171a | 0.193b | 0.174b | 0.158b | 0.085b | |
| SEm± | 0.018 | 0.018 | 0.01 | 0.01 | 0.06 | 0.03 | 0.002 | |
| T | ** | ** | ** | ** | ns | ** | ** | |
| G | ** | * | ** | ** | ** | ** | * | |
| Interaction | T × G | ** | ns | * | ns | * | ns | ns |
| Interaction Effects | Selenium Content (mg/kg) | Organic Se Content (mg/kg) | Inorganic Se Content (mg/kg) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Soil | Roots | Stem | Leaves | Husk | Brown Rice | Polished Rice | Brown Rice | Polished Rice | Brown Rice | Polished Rice | ||
| Treatment (T) | Control | 0.272a | 0.674ab | 0.196c | 0.109c | 0.059c | 0.045c | 0.045c | 0.042c | 0.08c | 0.017c | 0.009 |
| Si15Cd1 | 0.249ab | 0.66ab | 0.281b | 0.157b | 0.067c | 0.055b | 0.055b | 0.05bc | 0.045b | 0.018c | 0.01 | |
| Si30Cd1 | 0.222bc | 0.714a | 0.337a | 0.217a | 0.093a | 0.057b | 0.057b | 0.072a | 0.048b | 0.021b | 0.009 | |
| Si60Cd1 | 0.208d | 0.644b | 0.266b | 0.219a | 0.082b | 0.064a | 0.064a | 0.058b | 0.054a | 0.024a | 0.01 | |
| SEm± | 0.009 | 0.016 | 0.01 | 0.009 | 0.003 | 0.00 | 0.00 | 0.033 | 0.001 | 0.001 | 0.001 | |
| Genotypes (G) | G46B | 0.33a | 0.93a | 0.185b | 0.086b | 0.052b | 0.037b | 0.037b | 0.027b | 0.027b | 0.019b | 0.009 |
| Z3055B | 0.145b | 0.416b | 0.355a | 0.265a | 0.099 | 0.075a | 0.075a | 0.054a | 0.065a | 0.021a | 0.01 | |
| SEm± | 0.006 | 0.011 | 0.007 | 0.006 | 0.002 | 0.001 | 0.001 | 0.002 | 0.001 | 0.001 | 0.001 | |
| T | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ns | |
| G | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ns | |
| Interaction | T × G | ns | ** | ** | ** | ** | ** | * | * | * | * | ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, Y.; Huang, X.; Li, L.; Muhammad, Z.A.; Li, M.; Zheng, T.; Guo, Z.; Zhang, Y.; Luo, D.; Ye, X.; et al. Comparative Responses of Silicon to Reduce Cadmium and Enrich Selenium in Rice Varieties. Foods 2023, 12, 1656. https://doi.org/10.3390/foods12081656
Su Y, Huang X, Li L, Muhammad ZA, Li M, Zheng T, Guo Z, Zhang Y, Luo D, Ye X, et al. Comparative Responses of Silicon to Reduce Cadmium and Enrich Selenium in Rice Varieties. Foods. 2023; 12(8):1656. https://doi.org/10.3390/foods12081656
Chicago/Turabian StyleSu, Yang, Xin Huang, Ling Li, Zahir Ahsan Muhammad, Meilin Li, Tengda Zheng, Zhe Guo, Yue Zhang, Dan Luo, Xiaoying Ye, and et al. 2023. "Comparative Responses of Silicon to Reduce Cadmium and Enrich Selenium in Rice Varieties" Foods 12, no. 8: 1656. https://doi.org/10.3390/foods12081656
APA StyleSu, Y., Huang, X., Li, L., Muhammad, Z. A., Li, M., Zheng, T., Guo, Z., Zhang, Y., Luo, D., Ye, X., Jia, X., Hussain Panhwar, F., Tun, M. T., & Zhu, J. (2023). Comparative Responses of Silicon to Reduce Cadmium and Enrich Selenium in Rice Varieties. Foods, 12(8), 1656. https://doi.org/10.3390/foods12081656