

The Microbial Diversity on the Surface of Smear-Ripened Cheeses and Its Impact on Cheese Quality and Safety

Abstract
:1. Introduction
1.1. Cheese Varieties
1.2. Smear-Ripened Cheese
2. The Manufacture of Smear-Ripened Cheese
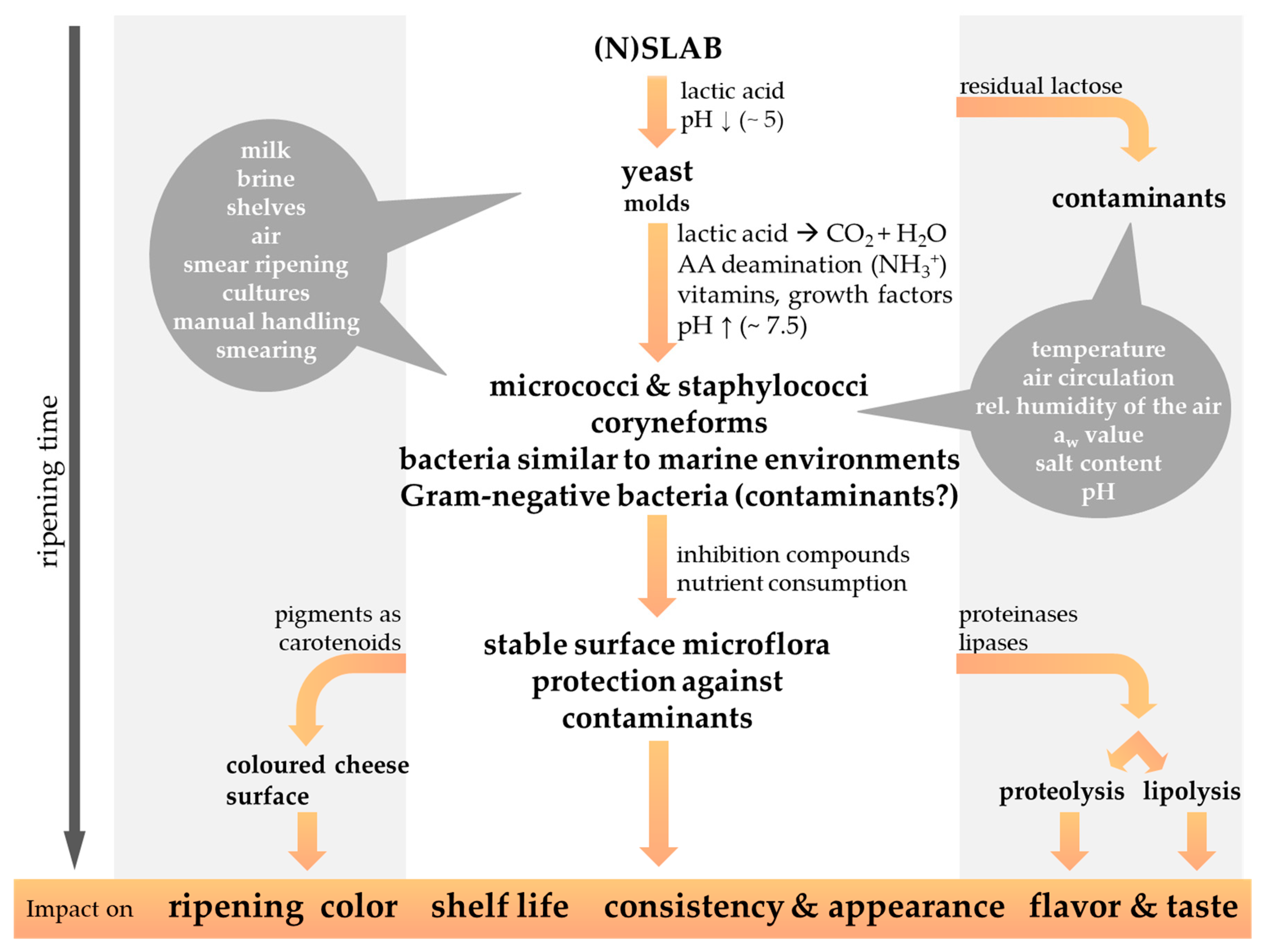
3. The Ripening of Smear-Ripened Cheese

4. The Microbiology of Smear-Ripened Cheese
4.1. The Microbiota of the Cheese Interior
4.2. The Smear Microbiota on the Cheese Surface

{kind=link}
{kind=link}
{kind=link}
| Phylum Order | Family | Genus | Atmospheric Conditions | % of Smear Microbiota a, b | Contribution to Cheese Ripening | References |
|---|---|---|---|---|---|---|
| Firmicutes | ||||||
| Bacillales | Staphylococcaceae | Staphylococcus | facultative anaerobic | 5–18% a,b | proteases, lipases, growth factors, flavor compounds, bacteriocins | [23,32,33,47,60,62,65,91,100,143,144,161,172,173] |
| Lactobacillales | Carnobacteriaceae | Alkalibacterium | facultative anaerobic | not known | lactic acid | [64,91,161,172,173] |
| Lactobacillales | Carnobacteriaceae | Marinilactibacillus | facultative anaerobic | 2–5% a,b | lactic acid, flavor compounds | [64,91,100,160,162,171,172] |
| Lactobacillales | Aerococcaceae | Facklamia | facultative anaerobic | 0.6% a | lipases | [92,172,173] |
| Lactobacillales | Lactobacillaceae | Lactobacillus | anaerobic | 0.2% a | >proteases, lipases, lactic acid | [65,78,92,172,173] |
| Lactobacillales | Enterococcaceae | Enterococcus | facultative anaerobic | <1% a,b | proteases, lipases, flavor compounds, bacteriocins | [32,81,100,133,134,135] |
| Actinobacteria | ||||||
| Actinomycetales | Micrococcaceae | Arthrobacter | aerobic | 0.1–5% a,b | proteinases, lipases, flavor compounds, pigments, bacteriocins | [32,60,65,89,92,100,125,143,172] |
| Actinomycetales | Micrococcaceae | Micrococcus | aerobic | 0.1–20% b | proteinases, lipases, pigments | [32,143,174] |
| Actinomycetales | Brevibacteriaceae | Brevibacterium | aerobic | 1–30% a,b | proteases, lipases, flavor compounds, pigments, growth factors, bacteriocins | [11,16,17,23,47,60,61,65,78,89,91,92,99,100,144,161,172,173] |
| Actinomycetales | Dermabacteraceae | Brachybacterium | facultative anaerobic | 3.6–7.3% a | flavor compounds, pigments | [11,16,17,32,60,92,157,172,173] |
| Actinomycetales | Corynebacteriaceae | Corynebacterium | facultative anaerobic | 1.2–70% a,b | esterases, lipases, flavor compounds | [16,23,60,65,89,91,100,143,144,172,173] |
| Actinomycetales | Microbacteriaceae | Microbacterium | facultative anaerobic | 0.9% a | flavor compounds, lactic acid, pigments, growth factors | [16,60,172,173] |
| Actinomycetales | Microbacteriaceae | Leucobacter | aerobic | 0.08% a | not known | [60,173] |
| Proteobacteria | ||||||
| Oceanospirillales | Halomonadaceae | Halomonas | facultative anaerobic | 2.5–29.1% a,b | >not known | [46,47,60,92,143,144,161,172,173] |
| Enterobacterales | Enterobacteriaceae | Enterobacter | facultative anaerobic | 0.02% a | proteases, flavor compounds, pigments | [60,172,173] |
| Enterobacterales | Enterobacteriaceae | Proteus | facultative anaerobic | 6.3% b | proteinases, lipases, flavor compounds | [146,158,172,173] |
| Pseudomonadales | Moraxellaceae | Psychrobacter | aerobic | 3–9.9% a,b | >flavor compounds | [46,60,65,89,92,100,143,144,158,159,161,171,173] |
| Alteromonadales | Pseudoalteromonadaceae | Pseudoalteromonas | aerobic | 3.5–3.8% a | extracellular enzymes, pigments | [60,65,92,100] |
5. Cheese Smear Defects
6. Conclusions and Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Curry, A. The milk revolution. Nature 2013, 500, 20–22. [Google Scholar] [CrossRef] [PubMed]
- Gerbault, P.; Roffet-Salque, M.; Evershed, R.P.; Thomas, M.G. How long have adult humans been consuming milk? IUBMB Life 2013, 65, 983–990. [Google Scholar] [CrossRef] [PubMed]
- Salque, M.; Bogucki, P.I.; Pyzel, J.; Sobkowiak-Tabaka, I.; Grygiel, R.; Szmyt, M.; Evershed, R.P. Earliest evidence for cheese making in the sixth millennium bc in northern Europe. Nature 2013, 493, 522–525. [Google Scholar] [CrossRef] [PubMed]
- Fox, P.F.; McSweeney, P.L.H. Cheese: An Overview. In Cheese: Chemistry, Physics and Microbiology; Patrick, F., Fox, P.F., McSweeney, P.L.H., Cogan, T.M., Guinee, T.P., Eds.; Elsevier: Amsterdam, The Netherlands, 2004; Volume 1, pp. 1–18. [Google Scholar]
- O’Brien, N.M.; O’Connor, T.P. Nutritional aspects of cheese. In Cheese: Chemistry, Physics and Microbiology; Fox, P.F., McSweeney, P.L.H., Cogan, T.M., Guinee, T.P., Eds.; Elsevier: Amsterdam, The Netherlands, 2004; Volume 1, pp. 573–581. [Google Scholar]
- Quigley, L.; O’Sullivan, O.; Stanton, C.; Beresford, T.P.; Ross, R.P.; Fitzgerald, G.F.; Cotter, P.D. The complex microbiota of raw milk. FEMS Microbiol. Rev. 2013, 37, 664–698. [Google Scholar] [CrossRef] [PubMed]
- Pepi, M.; Focardi, S. The Microbiology of Cheese and Dairy Products is a Critical Step in Ensuring Health, Quality and Typicity. Corpus J. Dairy Vet. Sci. 2022, 3, 9. [Google Scholar]
- Codex Alimentarius Commision. Codex Alimentarius: Milk and Milk Products; Joint World Health Organization and Food and Agriculture Organization of the United Nations: Rome, Italy, 2011; Volume 2. [Google Scholar]
- Donnelly, C.W. Cheese and Microbes; American Society of Microbiology: Washington, DC, USA, 2014. [Google Scholar] [CrossRef]
- McSweeney, P.L.H.; Ottogalli, G.; Fox, P.F. Diversity of cheese varieties: An overview. In Cheese: Chemistry, Physics and Microbiology; Fox, P.F., McSweeney, P.L.H., Cogan, T.M., Guinee, T.P., Eds.; Elsevier: Amsterdam, The Netherlands, 2004; Volume 2, pp. 1–23. [Google Scholar]
- Brennan, N.M.; Cogan, T.M.; Loessner, M.; Scherer, S. Bacterial surface-ripened cheeses. In Cheese: Chemistry, Physics and Microbiology; Fox, P.F., McSweeney, P.L.H., Cogan, T.M., Guinee, T.P., Eds.; Elsevier: Amsterdam, The Ntherlands, 2004; Volume 2, pp. 199–225. [Google Scholar]
- Bokulich, N.A.; Mills, D.A. Facility-specific “house” microbiome drives microbial landscapes of artisan cheesemaking plants. Appl. Environ. Microbiol. 2013, 79, 5214–5223. [Google Scholar] [CrossRef] [PubMed]
- Mounier, J.; Goerges, S.; Gelsomino, R.; Vancanneyt, M.; Vandemeulebroecke, K.; Hoste, B.; Brennan, N.; Scherer, S.; Swings, J.; Fitzgerald, G. Sources of the adventitious microflora of a smear ripened cheese. J. Appl. Microbiol. 2006, 101, 668–681. [Google Scholar] [CrossRef]
- Settanni, L.; Busetta, G.; Puccio, V.; Licitra, G.; Franciosi, E.; Botta, L.; Di Gerlando, R.; Todaro, M.; Gaglio, R. In-Depth Investigation of the Safety of Wooden Shelves Used for Traditional Cheese Ripening. Appl. Environ. Microbiol. 2021, 87, e01524-21. [Google Scholar] [CrossRef]
- Vermote, L.; Verce, M.; De Vuyst, L.; Weckx, S. Amplicon and shotgun metagenomic sequencing indicates that microbial ecosystems present in cheese brines reflect environmental inoculation during the cheese production process. Int. Dairy J. 2018, 87, 44–53. [Google Scholar] [CrossRef]
- Bockelmann, W. Secondary cheese starter cultures. In Technology of Cheesemaking; Law, B.A., Tamime, A.Y., Eds.; Blackwell Publishing Ltd.: Oxford, UK, 2010; Volume 14, pp. 193–230. [Google Scholar]
- Bockelmann, W.; Hoppe-Seyler, T. The surface flora of bacterial smear-ripened cheeses from cow’s and goat’s milk. Int. Dairy J. 2001, 11, 307–314. [Google Scholar] [CrossRef]
- Ritschard, J.S.; Van Loon, H.; Amato, L.; Meile, L.; Schuppler, M. High Prevalence of Enterobacterales in the Smear of Surface-Ripened Cheese with Contribution to Organoleptic Properties. Foods 2022, 11, 361. [Google Scholar] [CrossRef] [PubMed]
- Ryser, L.T.; Arias-Roth, E.; Perreten, V.; Irmler, S.; Bruggmann, R. Genetic and Phenotypic Diversity of Morganella morganii Isolated From Cheese. Front. Microbiol. 2021, 12, 738492. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, C.W. Growth and Survival of Microbial Pathogens in Cheese. In Cheese: Chemistry, Physics and Microbiology; Patrick, F., Fox, P.F., McSweeney, P.L.H., Timothy, P.G., Eds.; Elsevier: Amsterdam, The Netherlands, 2004; Volume 1, pp. 541–559. [Google Scholar]
- Mounier, J.; Monnet, C.; Jacques, N.; Antoinette, A.; Irlinger, F. Assessment of the microbial diversity at the surface of Livarot cheese using culture-dependent and independent approaches. Int. J. Food Microbiol. 2009, 133, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Reps, A. Bacterial Surface-Ripened Cheeses. In Cheese: Chemistry, Physics and Microbiology; Fox, P.F., Ed.; Springer US: New York, NY, USA, 1993; pp. 137–172. [Google Scholar] [CrossRef]
- Bertuzzi, A.S.; Walsh, A.M.; Sheehan, J.J.; Cotter, P.D.; Crispie, F.; McSweeney, P.L.H.; Kilcawley, K.N.; Rea, M.C. Omics-Based Insights into Flavor Development and Microbial Succession within Surface-Ripened Cheese. MSystems 2018, 3, e00211-17. [Google Scholar] [CrossRef] [PubMed]
- Bockelmann, W.; Willems, K.; Neve, H.; Heller, K. Cultures for the ripening of smear cheeses. Int. Dairy J. 2005, 15, 719–732. [Google Scholar] [CrossRef]
- Falardeau, J.; Yildiz, E.; Yan, Y.F.; Castellarin, S.D.; Wang, S.Y. Microbiome and Physicochemical Features Associated with Differential Listeria monocytogenes Growth in Soft, Surface-Ripened Cheeses. Appl. Environ. Microbiol. 2023, 89, e0200422. [Google Scholar] [CrossRef]
- Hammer, P.; Bockelmann, W.; Hoffmann, W. Fate of Listeria innocua during production and ripening of smeared hard cheese made from raw milk. J. Dairy Sci. 2017, 100, 7846–7856. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Bré, J.-M.; Guéguen, M.; Vernoux, J.-P.; Desmasures, N. Reduced growth of Listeria monocytogenes in two model cheese microcosms is not associated with individual microbial strains. Food Microbiol. 2013, 33, 30–39. [Google Scholar] [CrossRef]
- Gaulin, C.; Ramsay, D.; Bekal, S. Widespread listeriosis outbreak attributable to pasteurized cheese, which led to extensive cross-contamination affecting cheese retailers, Quebec, Canada, 2008. J. Food Prot. 2012, 75, 71–78. [Google Scholar] [CrossRef]
- Gérard, A.; El-Hajjaji, S.; Niyonzima, E.; Daube, G.; Sindic, M. Prevalence and survival of in various types of cheese—A review. Int. J. Dairy Technol. 2018, 71, 825–843. [Google Scholar] [CrossRef]
- Makino, S.I.; Kawamoto, K.; Takeshi, K.; Okada, Y.; Yamasaki, M.; Yamamoto, S.; Igimi, S. An outbreak of food-borne listeriosis due to cheese in Japan, during 2001. Int. J. Food Microbiol. 2005, 104, 189–196. [Google Scholar] [CrossRef]
- Monnet, C.; Bleicher, A.; Neuhaus, K.; Sarthou, A.-S.; Leclercq-Perlat, M.-N.; Irlinger, F. Assessment of the anti-listerial activity of microfloras from the surface of smear-ripened cheeses. Food Microbiol. 2010, 27, 302–310. [Google Scholar] [CrossRef]
- Bockelmann, W. Smear-ripened cheeses. In Encyclopaedia of Dairy Sciences; Roginski, H., Fuquay, J.W., Fox, P.F., Eds.; Academic Press: London, UK, 2002; pp. 391–401. [Google Scholar]
- Rea, M.; Görges, S.; Gelsomino, R.; Brennan, N.; Mounier, J.; Vancanneyt, M.; Scherer, S.; Swings, J.; Cogan, T. Stability of the biodiversity of the surface consortia of Gubbeen, a red-smear cheese. J. Dairy Sci. 2007, 90, 2200–2210. [Google Scholar] [CrossRef]
- Goerges, S.; Mounier, J.; Rea, M.C.; Gelsomino, R.; Heise, V.; Beduhn, R.; Cogan, T.M.; Vancanneyt, M.; Scherer, S. Commercial ripening starter microorganisms inoculated into cheese milk do not successfully establish themselves in the resident microbial ripening consortia of a South German red smear cheese. Appl. Environ. Microbiol. 2008, 74, 2210–2217. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.Q.; Zhao, G.; Gu, L.Y.; Solem, C. A novel approach for accelerating smear development on bacterial smear-ripened cheeses reduces ripening time and inhibits the growth of Listeria and other unwanted microorganisms on the rind. LWT Food Sci. Technol. 2022, 170, 114109. [Google Scholar] [CrossRef]
- Desmasures, N.; Bora, N.; Dodd, C.; Ward, A.C. Smear ripened cheeses. In Diversity, Dynamics and Functional Role of Actinomycetes on European Smear Ripened Cheeses; Bora, N., Dodd, C., Desmasures, N., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- Ardö, Y.; Berthier, F.; Hartmann, K.; Eugster-Meier, E.; Fröhlich-Wyder, M.-T.; Jakob, E.; Wechsler, D. Bacterial Surface-Ripened (Smear) Cheeses. In Global Cheesemaking Technology: Cheese Quality and Characteristics, 1st ed.; Papademas, P., Bintsis, T., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2018. [Google Scholar]
- Lücke, F.-K.; Zangerl, P. Food safety challenges associated with traditional foods in German-speaking regions. Food Control 2014, 43, 217–230. [Google Scholar] [CrossRef]
- Bennett, R.J.; Johnston, K.A. General aspects of cheese technology. In Cheese: Chemistry, Physics and Microbiology; Fox, P.F., McSweeney, P.L.H., Cogan, T.M., Guinee, T.P., Eds.; Elsevier: Amsterdam, The Netherlands, 2004; Volume 2, pp. 23–50. [Google Scholar]
- Montel, M.-C.; Buchin, S.; Mallet, A.; Delbès–Paus, C.; Vuitton, D.A.; Desmasures, N.; Berthier, F. Traditional cheeses: Rich and diverse microbiota with associated benefits. Int. J. Food Microbiol. 2014, 177, 136–154. [Google Scholar] [CrossRef] [PubMed]
- Samelis, J.; Lianou, A.; Kakouri, A.; Delbes, C.; Rogelj, I.; Bogovic-Matijasic, B.; Montel, M.C. Changes in the microbial composition of raw milk induced by thermization treatments applied prior to traditional Greek hard cheese processing. J. Food Prot. 2009, 72, 783–790. [Google Scholar] [CrossRef]
- Home, D.S.; Banks, J.M. Rennet-induced Coagulation of Milk. In Cheese: Chemistry, Physics and Microbiology; Fox, P.F., McSweeney, P.L.H., Cogan, T.M., Guinee, T.P., Eds.; Elsevier: Amsterdam, The Netherlands, 2004; Volume 1, pp. 47–70. [Google Scholar]
- Dejmek, P.; Walstra, P. The syneresis of rennet-coagulated curd. In Cheese: Chemistry, Physics and Microbiology; Fox, P.F., McSweeney, P.L.H., Cogan, T.M., Guinee, T.P., Eds.; Elsevier: Amsterdam, The Netherlands, 2004; Volume 1, pp. 71–103. [Google Scholar]
- Guinee, T.P.; Fox, P.F. Salt in Cheese: Physical, Chemical and Biological Aspects. In Cheese: Chemistry, Physics and Microbiology; Fox, P.F., McSweeney, P.L.H., Cogan, T.M., Guinee, T.P., Eds.; Elsevier: Amsterdam, The Netherlands, 2004; Volume 1, pp. 207–259. [Google Scholar]
- Feurer, C.; Vallaeys, T.; Corrieu, G.; Irlinger, F. Does smearing inoculum reflect the bacterial composition of the smear at the end of the ripening of a French soft, red-smear cheese? J. Dairy Sci. 2004, 87, 3189–3197. [Google Scholar] [CrossRef]
- Irlinger, F.; Monnet, C. Temporal differences in microbial composition of Epoisses cheese rinds during ripening and storage. J. Dairy Sci. 2021, 104, 7500–7508. [Google Scholar] [CrossRef]
- Quijada, N.M.; Mann, E.; Wagner, M.; Rodríguez-Lázaro, D.; Hernández, M.; Schmitz-Esser, S. Autochthonous facility-specific microbiota dominates washed-rind Austrian hard cheese surfaces and its production environment. Int. J. Food Microbiol. 2018, 267, 54–61. [Google Scholar] [CrossRef]
- Winkler, H.; Bobst, C.; Amrein, R. Käsepflege ohne Farbe. ALP Forum 2004, 5, 1–7. [Google Scholar]
- Mounier, J.; Coton, M.; Irlinger, F.; Landaud, S.; Bonnarme, P. Smear-Ripened Cheeses. In Cheese: Chemistry, Physics and Microbiology, 4th ed.; Elsevier: Amsterdam, The Netherlands, 2017; Volume 1–2, pp. 955–996. [Google Scholar] [CrossRef]
- De Buyser, M.-L.; Dufour, B.; Maire, M.; Lafarge, V. Implication of milk and milk products in food-borne diseases in France and in different industrialised countries. Int. J. Food Microbiol. 2001, 67, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Honish, L.; Predy, G.; Hislop, N.; Chui, L.; Kowalewska-Grochowska, K.; Trottier, L.; Kreplin, C.; Zazulak, I. An outbreak of E. coli O157: H7 hemorrhagic colitis associated with unpasteurized Gouda cheese. Can. J. Public Health 2005, 96, 182–184. [Google Scholar] [CrossRef]
- McCollum, J.; Williams, N.; Beam, S.; Cosgrove, S.; Ettestad, P.; Ghosh, T.; Kimura, A.; Nguyen, L.; Stroika, S.; Vogt, R. Multistate outbreak of Escherichia coli O157: H7 infections associated with in-store sampling of an aged raw-milk gouda cheese, 2010. J. Food Prot. 2012, 75, 1759–1765. [Google Scholar] [CrossRef]
- Zottola, E.A.; Smith, L.B. Pathogens in cheese. Food Microbiol. 1991, 8, 171–182. [Google Scholar] [CrossRef]
- Sutthiwong, N.; Fouillaud, M.; Dufossé, L. The Influence of pH, NaCl, and the Deacidifying Yeasts Debaryomyces hansenii and Kluyveromyces marxianus on the Production of Pigments by the Cheese-Ripening Bacteria Arthrobacter arilaitensis. Foods 2018, 7, 190. [Google Scholar] [CrossRef]
- Sutthiwong, N.; Sukdee, P.; Lekhavat, S.; Dufosse, L. Identification of Red Pigments Produced by Cheese-Ripening Bacterial Strains of Glutamicibacter arilaitensis Using HPLC. Dairy 2021, 2, 396–409. [Google Scholar] [CrossRef]
- Wolfe, B.E.; Dutton, R.J. Towards an Ecosystem Approach to Cheese Microbiology. Microbiol. Spectr. 2013, 1, 311–321. [Google Scholar] [CrossRef]
- Jany, J.-L.; Barbier, G. Culture-independent methods for identifying microbial communities in cheese. Food Microbiol. 2008, 25, 839–848. [Google Scholar] [CrossRef]
- Justé, A.; Thomma, B.P.H.J.; Lievens, B. Recent advances in molecular techniques to study microbial communities in food-associated matrices and processes. Food Microbiol. 2008, 25, 745–761. [Google Scholar] [CrossRef]
- Quigley, L.; O’Sullivan, O.; Beresford, T.P.; Ross, R.P.; Fitzgerald, G.F.; Cotter, P.D. Molecular approaches to analysing the microbial composition of raw milk and raw milk cheese. Int. J. Food Microbiol. 2011, 150, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, B.E.; Button, J.E.; Santarelli, M.; Dutton, R.J. Cheese rind communities provide tractable systems for in situ and in vitro studies of microbial diversity. Cell 2014, 158, 422–433. [Google Scholar] [CrossRef] [PubMed]
- Bockelmann, W.; Krusch, U.; Engel, G.; Klijn, N.; Smit, G.; Heller, K. The microflora of Tilsit cheese. Part 1. Variability of the smear flora. Food/Nahrung 1997, 41, 208–212. [Google Scholar] [CrossRef]
- Brennan, N.M.; Ward, A.C.; Beresford, T.P.; Fox, P.F.; Goodfellow, M.; Cogan, T.M. Biodiversity of the bacterial flora on the surface of a smear cheese. Appl. Environ. Microbiol. 2002, 68, 820–830. [Google Scholar] [CrossRef]
- Mounier, J.; Gelsomino, R.; Goerges, S.; Vancanneyt, M.; Vandemeulebroecke, K.; Hoste, B.; Scherer, S.; Swings, J.; Fitzgerald, G.F.; Cogan, T.M. Surface microflora of four smear-ripened cheeses. Appl. Environ. Microbiol. 2005, 71, 6489–6500. [Google Scholar] [CrossRef] [PubMed]
- Roth, E.; Schwenninger, S.M.; Eugster-Meier, E.; Lacroix, C. Facultative anaerobic halophilic and alkaliphilic bacteria isolated from a natural smear ecosystem inhibit Listeria growth in early ripening stages. Int. J. Food Microbiol. 2011, 147, 26–32. [Google Scholar] [CrossRef]
- O’Sullivan, D.J.; Cotter, P.D.; O’Sullivan, O.; Giblin, L.; McSweeney, P.L.; Sheehan, J.J. Temporal and spatial differences in microbial composition during the manufacture of a continental-type cheese. Appl. Environ. Microbiol. 2015, 81, 2525–2533. [Google Scholar] [CrossRef]
- Cotter, P.D.; Beresford, T.P. Microbiome Changes During Ripening. In Cheese: Chemistry, Physics and Microbiology, 4th ed.; Elsevier: Amsterdam, The Netherlands, 2017; Volume 1–2, pp. 389–409. [Google Scholar] [CrossRef]
- Vacheyrou, M.; Normand, A.C.; Guyot, P.; Cassagne, C.; Piarroux, R.; Bouton, Y. Cultivable microbial communities in raw cow milk and potential transfers from stables of sixteen French farms. Int. J. Food Microbiol. 2011, 146, 253–262. [Google Scholar] [CrossRef]
- Verdier-Metz, I.; Gagne, G.; Bornes, S.; Monsallier, F.; Veisseire, P.; Delbès-Paus, C.; Montel, M.-C. Cow teat skin, a potential source of diverse microbial populations for cheese production. Appl. Environ. Microbiol. 2012, 78, 326–333. [Google Scholar] [CrossRef]
- Goldberg, J.J.; Wildman, E.E.; Pankey, J.W.; Kunkel, J.R.; Howard, D.B.; Murphy, B.M. The influence of intensively managed rotational grazing, traditional continuous grazing, and confinement housing on bulk tank milk quality and udder health. J. Dairy Sci. 1992, 75, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Hogan, J.S.; Smith, K.L.; Hoblet, K.H.; Todhunter, D.A.; Schoenberger, P.S.; Hueston, W.D.; Pritchard, D.E.; Bowman, G.L.; Heider, L.E.; Brockett, B.L.; et al. Bacterial counts in bedding materials used on nine commercial dairies. J. Dairy Sci. 1989, 72, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Marchand, S.; De Block, J.; De Jonghe, V.; Coorevits, A.; Heyndrickx, M.; Herman, L. Biofilm formation in milk production and processing environments; Influence on milk quality and safety. Compr. Rev. Food Sci. Food Saf. 2012, 11, 133–147. [Google Scholar] [CrossRef]
- Cousin, M. Presence and activity of psychrotrophic microorganisms in milk and dairy products; a review. J. Food Prot. 1982, 45, 172–207. [Google Scholar] [CrossRef]
- Hantsis-Zacharov, E.; Halpern, M. Culturable Psychrotrophic Bacterial Communities in Raw Milk and Their Proteolytic and Lypolytic Traits. Appl. Environ. Microbiol. 2007, 73, 7162–7168. [Google Scholar] [CrossRef]
- O’Sullivan, D.; Cotter, P.; Giblin, L.; Sheehan, J.; McSweeney, P. Nucleic acid-based approaches to investigate microbial-related cheese quality defects. Front. Microbiol. 2013, 4, 1. [Google Scholar] [CrossRef] [PubMed]
- Fox, P.F.; Guinee, T.P.; Cogan, T.M.; McSweeney, P.L.H. Pathogens in Cheese and Foodborne Illnesses. In Fundamentals of Cheese Science, 2nd ed.; Fox, P.F., Guinee, T.P., Cogan, T.M., McSweeney, P.L.H., Eds.; Springer: New York, NY, USA, 2017. [Google Scholar] [CrossRef]
- Oliver, S.P.; Boor, K.J.; Murphy, S.C.; Murinda, S.E. Food safety hazards associated with consumption of raw milk. Foodborne Pathog. Dis. 2009, 6, 793–806. [Google Scholar] [CrossRef]
- Beresford, T.; Williams, A. The Microbiology of Cheese Ripening. In Cheese: Chemistry, Physics and Microbiology; Fox, P.F., McSweeney, P.L.H., Cogan, T.M., Guinee, T.P., Eds.; Elsevier: Amsterdam, The Netherlands, 2004; Volume 1, pp. 287–317. [Google Scholar]
- Beuvier, E.; Buchin, S. Raw milk cheeses. In Cheese: Chemistry, Physics and Microbiology; Fox, P.F., McSweeney, P.L.H., Cogan, T.M., Guinee, T.P., Eds.; Elsevier: Amsterdam, The Netherlands, 2004; Volume 1, pp. 319–345. [Google Scholar]
- Paxson, H. Post-pasteurian cultures: The microbiopolitics of raw-milk cheese in the United States. Cult. Anthropol. 2008, 23, 15–47. [Google Scholar] [CrossRef]
- Paxson, H. The Life of Cheese: Crafting Food and Value in America; University of California Press: Los Angeles, CA, USA, 2012. [Google Scholar]
- Callon, C.; Retureau, E.; Didienne, R.; Montel, M.-C. Microbial biodiversity in cheese consortia and comparative Listeria growth on surfaces of uncooked pressed cheeses. Int. J. Food Microbiol. 2014, 174, 98–109. [Google Scholar] [CrossRef]
- Ksontini, H.; Kachouri, F.; Hamdi, M. Dairy biofilm: Impact of microbial community on raw milk quality. J. Food Qual. 2013, 36, 282–290. [Google Scholar] [CrossRef]
- Didienne, R.; Defargues, C.; Callon, C.; Meylheuc, T.; Hulin, S.; Montel, M.-C. Characteristics of microbial biofilm on wooden vats (‘gerles’) in PDO Salers cheese. Int. J. Food Microbiol. 2012, 156, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Elias, S.; Banin, E. Multi-species biofilms: Living with friendly neighbors. FEMS Microbiol. Rev. 2012, 36, 990–1004. [Google Scholar] [CrossRef] [PubMed]
- Rendueles, O.; Ghigo, J.-M. Multi-species biofilms: How to avoid unfriendly neighbors. FEMS Microbiol. Rev. 2012, 36, 972–989. [Google Scholar] [CrossRef] [PubMed]
- Schoder, D.; Rossmanith, P.; Glaser, K.; Wagner, M. Fluctuation in contamination dynamics of L. monocytogenes in quargel (acid curd cheese) lots recalled during the multinational listeriosis outbreak 2009/2010. Int. J. Food Microbiol. 2012, 157, 326–331. [Google Scholar] [CrossRef] [PubMed]
- Parente, E.; Cogan, T.M. Starter cultures: General aspects. In Cheese: Chemistry, Physics and Microbiology; Fox, P.F., McSweeney, P.L.H., Cogan, T.M., Guinee, T.P., Eds.; Elsevier: Amsterdam, The Netherlands, 2004; Volume 1, pp. 123–147. [Google Scholar]
- Masoud, W.; Takamiya, M.; Vogensen, F.K.; Lillevang, S.; Al-Soud, W.A.; Sørensen, S.J.; Jakobsen, M. Characterization of bacterial populations in Danish raw milk cheeses made with different starter cultures by denaturating gradient gel electrophoresis and pyrosequencing. Int. Dairy J. 2011, 21, 142–148. [Google Scholar] [CrossRef]
- Dalmasso, A.; del Rio, M.D.S.; Civera, T.; Pattono, D.; Cardazzo, B.; Bottero, M.T. Characterization of microbiota in Plaisentif cheese by high-throughput sequencing. LWT Food Sci. Technol. 2016, 69, 490–496. [Google Scholar] [CrossRef]
- Dugat-Bony, E.; Garnier, L.; Denonfoux, J.; Ferreira, S.; Sarthou, A.S.; Bonnarme, P.; Irlinger, F. Highlighting the microbial diversity of 12 French cheese varieties. Int. J. Food Microbiol. 2016, 238, 265–273. [Google Scholar] [CrossRef]
- Ryssel, M.; Johansen, P.; Abu Al-Soud, W.; Sorensen, S.; Arneborg, N.; Jespersen, L. Microbial diversity and dynamics throughout manufacturing and ripening of surface ripened semi-hard Danish Danbo cheeses investigated by culture-independent techniques. Int. J. Food Microbiol. 2015, 215, 124–130. [Google Scholar] [CrossRef]
- Quigley, L.; O’Sullivan, O.; Beresford, T.P.; Ross, R.P.; Fitzgerald, G.F.; Cotter, P.D. High-Throughput Sequencing for detection of subpopulations of bacteria not previously associated with artisanal cheeses. Appl. Environ. Microbiol. 2012, 78, 5717–5723. [Google Scholar] [CrossRef]
- Monnet, C.; Loux, V.; Bento, P.; Gibrat, J.-F.; Straub, C.; Bonnarme, P.; Landaud, S.; Irlinger, F. Genome sequence of Corynebacterium casei UCMA 3821, isolated from a smear-ripened cheese. J. Bacteriol. 2012, 194, 738–739. [Google Scholar] [CrossRef]
- Mariani, C.; Briandet, R.; Chamba, J.F.; Notz, E.; Carnet-Pantiez, A.; Eyoug, R.; Oulahal, N. Biofilm ecology of wooden shelves used in ripening the French raw milk smear cheese Reblochon de Savoie. J. Dairy Sci. 2007, 90, 1653–1661. [Google Scholar] [CrossRef] [PubMed]
- Oulahal, N.; Adt, I.; Mariani, C.; Carnet-Pantiez, A.; Notz, E.; Degraeve, P. Examination of wooden shelves used in the ripening of a raw milk smear cheese by FTIR spectroscopy. Food Control 2009, 20, 658–663. [Google Scholar] [CrossRef]
- Baati, H.; Guermazi, S.; Gharsallah, N.; Sghir, A.; Ammar, E. Microbial community of salt crystals processed from Mediterranean seawater based on 16S rRNA analysis. Can. J. Microbiol. 2010, 56, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Dufossé, L.; Donadio, C.; Valla, A.; Meile, J.-C.; Montet, D. Determination of speciality food salt origin by using 16S rDNA fingerprinting of bacterial communities by PCR–DGGE: An application on marine salts produced in solar salterns from the French Atlantic Ocean. Food Control 2013, 32, 644–649. [Google Scholar] [CrossRef]
- Elevi Bardavid, R.; Khristo, P.; Oren, A. Interrelationships between Dunaliella and halophilic prokaryotes in saltern crystallizer ponds. Extremophiles 2008, 12, 5–14. [Google Scholar] [CrossRef]
- Sun, L.; D’Amico, D.J. Composition, Succession, and Source Tracking of Microbial Communities throughout the Traditional Production of a Farmstead Cheese. MSystems 2021, 26, 18. [Google Scholar] [CrossRef]
- Delcenserie, V.; Taminiau, B.; Delhalle, L.; Nezer, C.; Doyen, P.; Crevecoeur, S.; Roussey, D.; Korsak, N.; Daube, G. Microbiota characterization of a Belgian protected designation of origin cheese, Herve cheese, using metagenomic analysis. J. Dairy Sci. 2014, 97, 6046–6056. [Google Scholar] [CrossRef]
- Dolci, P.; Barmaz, A.; Zenato, S.; Pramotton, R.; Alessandria, V.; Cocolin, L.; Rantsiou, K.; Ambrosoli, R. Maturing dynamics of surface microflora in Fontina PDO cheese studied by culture dependent and independent methods. J. Appl. Microbiol. 2009, 106, 278–287. [Google Scholar] [CrossRef]
- Rademaker, J.; Peinhopf, M.; Rijnen, L.; Bockelmann, W.; Noordman, W. The surface microflora dynamics of bacterial smear-ripened Tilsit cheese determined by T-RFLP DNA population fingerprint analysis. Int. Dairy J. 2005, 15, 785–794. [Google Scholar] [CrossRef]
- Fox, P.F.; Cogan, T.M. Factors that Affect the Quality of Cheese. In Cheese: Chemistry, Physics and Microbiology; Fox, P.F., McSweeney, P.L.H., Cogan, T.M., Guinee, T.P., Eds.; Elsevier: Amsterdam, The Netherlands, 2004; Volume 1, pp. 583–608. [Google Scholar]
- Bintsis, T. Yeasts in different types of cheese. Aims Microbiol. 2021, 7, 447–470. [Google Scholar] [CrossRef]
- Monnet, C.; Dugat-Bony, E.; Swennen, D.; Beckerich, J.M.; Irlinger, F.; Fraud, S.; Bonnarme, P. Investigation of the Activity of the Microorganisms in a Reblochon-Style Cheese by Metatranscriptomic Analysis. Front. Microbiol. 2016, 7, 536. [Google Scholar] [CrossRef] [PubMed]
- Prillinger, H.; Molnár, O.; Eliskases-Lechner, F.; Lopandic, K. Phenotypic and genotypic identification of yeasts from cheese. Antonie Van Leeuwenhoek 1999, 75, 267–283. [Google Scholar] [CrossRef] [PubMed]
- Corsetti, A.; Rossi, J.; Gobbetti, M. Interactions between yeasts and bacteria in the smear surface-ripened cheeses. Int. J. Food Microbiol. 2001, 69, 1–10. [Google Scholar] [CrossRef]
- Mounier, J. Microbial Interactions in Smear-Ripened Cheeses. In Diversity, Dynamics and Functional Role of Actinomycetes on European Smear Ripened Cheeses; Bora, N., Dodd, C., Desmasures, N., Eds.; Springer: Cham, Switzerland, 2015. [Google Scholar] [CrossRef]
- Bertuzzi, A.S.; Kilcawley, K.N.; Sheehan, J.J.; O’Sullivan, M.G.; Kennedy, D.; McSweeney, P.L.H.; Rea, M.C. Use of smear bacteria and yeasts to modify flavour and appearance of Cheddar cheese. Int. Dairy J. 2017, 72, 44–54. [Google Scholar] [CrossRef]
- Ruaro, A.; Andrighetto, C.; Torriani, S.; Lombardi, A. Biodiversity and characterization of indigenous coagulase-negative staphylococci isolated from raw milk and cheese of North Italy. Food Microbiol. 2013, 34, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Delbès, C.; Ali-Mandjee, L.; Montel, M.C. Monitoring bacterial communities in raw milk and cheese by culture-dependent and-independent 16S rRNA gene-based analyses. Appl. Environ. Microbiol. 2007, 73, 1882–1891. [Google Scholar] [CrossRef] [PubMed]
- Hoppe-Seyler, T.S.; Jaeger, B.; Bockelmann, W.; Noordman, W.H.; Geis, A.; Heller, K.J. Molecular identification and differentiation of Staphylococcus species and strains of cheese origin. Syst. Appl. Microbiol. 2004, 27, 211–218. [Google Scholar] [CrossRef]
- Meugnier, H.; Bes, M.; Vernozy-Rozand, C.; Mazuy, C.; Brun, Y.; Freney, J.; Fleurette, J. Identification and ribotyping of Staphylococcus xylosus and Staphylococcus equorum strains isolated from goat milk and cheese. Int. J. Food Microbiol. 1996, 31, 325–331. [Google Scholar] [CrossRef]
- Place, R.B.; Hiestand, D.; Burri, S.; Teuber, M. Staphylococcus succinus subsp. casei subsp. nov., a dominant isolate from a surface ripened cheese. Syst. Appl. Microbiol. 2002, 25, 353–359. [Google Scholar] [CrossRef]
- Place, R.B.; Hiestand, D.; Gallmann, H.R.; Teuber, M. Staphylococcus equorum subsp. linens, subsp. nov., a starter culture component for surface ripened semi-hard cheeses. Syst. Appl. Microbiol. 2003, 26, 30–37. [Google Scholar] [CrossRef]
- Eliskases-Lechner, F.; Ginzinger, W. The bacterial flora of surface-ripened cheeses with special regard to coryneforms. Le Lait 1995, 75, 571–584. [Google Scholar] [CrossRef]
- Bockelmann, W.; Hoppe Seyler, T.; Krusch, U.; Hoffmann, W.; Heller, K. The microflora of Tilsit cheese. Part 2. Development of a surface smear starter culture. Food/Nahrung 1997, 41, 213–218. [Google Scholar] [CrossRef]
- Rattray, F.P.; Fox, P.F. Aspects of enzymology and biochemical properties of Brevibacterium linens relevant to cheese ripening: A review. J. Dairy Sci. 1999, 82, 891–909. [Google Scholar] [CrossRef] [PubMed]
- Hoppe-Seyler, T.-S.; Jaeger, B.; Bockelmann, W.; Geis, A.; Heller, K.-J. Molecular identification and differentiation of Brevibacterium species and strains. Syst. Appl. Microbiol. 2007, 30, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Mounier, J.; Rea, M.C.; O’Connor, P.M.; Fitzgerald, G.F.; Cogan, T.M. Growth characteristics of Brevibacterium, Corynebacterium, Microbacterium, and Staphylococcus spp. isolated from surface-ripened cheese. Appl. Environ. Microbiol. 2007, 73, 7732–7739. [Google Scholar] [CrossRef] [PubMed]
- Motta, A.; Brandelli, A. Properties and antimicrobial activity of the smear surface cheese coryneform bacterium Brevibacterium linens. Eur. Food Res. Technol. 2008, 227, 1299–1306. [Google Scholar] [CrossRef]
- Irlinger, F.; Mounier, J. Microbial interactions in cheese: Implications for cheese quality and safety. Curr. Opin. Biotechnol. 2009, 20, 142–148. [Google Scholar] [CrossRef]
- Busse, H.J. Review of the taxonomy of the genus Arthrobacter, emendation of the genus Arthrobacter sensu lato, proposal to reclassify selected species of the genus Arthrobacter in the novel genera Glutamicibacter gen. nov., Paeniglutamicibacter gen. nov., Pseudoglutamicibacter gen. nov.,. Paenarthrobacter gen. nov. and Pseudarthrobacter gen. nov., and emended description of Arthrobacter roseus. Int. J. Syst. Evol. Microbiol. 2016, 66, 9–37. [Google Scholar] [CrossRef]
- Irlinger, F.; Bimet, F.; Delettre, J.; Lefèvre, M.; Grimont, P.A.D. Arthrobacter bergerei sp. nov. and Arthrobacter arilaitensis sp. nov., novel coryneform species isolated from the surfaces of cheeses. Int. J. Syst. Evol. Microbiol. 2005, 55, 457–462. [Google Scholar] [CrossRef]
- Monnet, C.; Loux, V.; Gibrat, J.-F.; Spinnler, E.; Barbe, V.; Vacherie, B.; Gavory, F.; Gourbeyre, E.; Siguier, P.; Chandler, M. The Arthrobacter arilaitensis Re117 genome sequence reveals its genetic adaptation to the surface of cheese. PLoS ONE 2010, 5, e15489. [Google Scholar] [CrossRef]
- Giuffrida, D.; Sutthiwong, N.; Dugo, P.; Donato, P.; Cacciola, F.; Girard-Valenciennes, E.; Le Mao, Y.; Monnet, C.; Fouillaud, M.; Caro, Y.; et al. Characterisation of the C50 carotenoids produced by strains of the cheese-ripening bacterium. Int. Dairy J. 2016, 55, 10–16. [Google Scholar] [CrossRef]
- Sutthiwong, N.; Dufossé, L. Production of carotenoids by Arthrobacter arilaitensis strains isolated from smear-ripened cheeses. FEMS Microbiol. Lett. 2014, 360, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Sutthiwong, N.; Caro, Y.; Milhau, C.; Valla, A.; Fouillaud, M.; Dufossé, L. Arthrobacter arilaitensis strains isolated from ripened cheeses: Characterization of their pigmentation using spectrocolorimetry. Food Res. Int. 2014, 65, 184–192. [Google Scholar] [CrossRef]
- Boldyreva, E.M.; Hill, A.; Griffiths, M.; Marcone, M. The quality and safety of washed-rind cheeses with a focus on antilisterial protection. Int. Dairy J. 2016, 55, 26–37. [Google Scholar] [CrossRef]
- Brennan, N.M.; Brown, R.; Goodfellow, M.; Ward, A.C.; Beresford, T.P.; Simpson, P.J.; Fox, P.F.; Cogan, T.M. Corynebacterium mooreparkense sp. nov. and Corynebacterium casei sp. nov., isolated from the surface of a smear-ripened cheese. Int. J. Syst. Evol. Microbiol. 2001, 51, 843–852. [Google Scholar] [CrossRef]
- Monnet, C.; Correia, K.; Sarthou, A.-S.; Irlinger, F. Quantitative detection of Corynebacterium casei in cheese by Real-Time PCR. Appl. Environ. Microbiol. 2006, 72, 6972–6979. [Google Scholar] [CrossRef]
- Walter, F.; Albersmeier, A.; Kalinowski, J.; Rückert, C. Complete genome sequence of Corynebacterium casei LMG S-19264T (=DSM 44701T), isolated from a smear-ripened cheese. J. Biotechnol. 2014, 189, 76–77. [Google Scholar] [CrossRef]
- Favaro, L.; Basaglia, M.; Casella, S.; Hue, I.; Dousset, X.; Dora Gombossy de Melo, F.B.; Todorov, S.D. Bacteriocinogenic potential and safety evaluation of non-starter Enterococcus faecium strains isolated from home made white brine cheese. Food Microbiol. 2014, 38, 228–239. [Google Scholar] [CrossRef]
- Moraes, P.M.; Perin, L.M.; Todorov, S.D.; Silva, A.; Franco, B.D.G.M.; Nero, L.A. Bacteriocinogenic and virulence potential of Enterococcus isolates obtained from raw milk and cheese. J. Appl. Microbiol. 2012, 113, 318–328. [Google Scholar] [CrossRef]
- Ogier, J.-C.; Serror, P. Safety assessment of dairy microorganisms: The Enterococcus genus. Int. J. Food Microbiol. 2008, 126, 291–301. [Google Scholar] [CrossRef]
- EFSA Panel on Biological Hazards (BIOHAZ). Scientific Opinion on the maintenance of the list of QPS biological agents intentionally added to food and feed (2013 update). EFSA J. 2013, 11, 3449–3557. [Google Scholar]
- EFSA Panel on Biological Hazards (BIOHAZ). Scientific Opinion: Statement on the update of the list of QPS-recommended biological agents intentionally added to food or feed as notified to EFSA 1: Suitability of taxonomic units notified to EFSA until October 2014. EFSA J. 2014, 12, 3938–3979. [Google Scholar]
- Ladero, V.; Fernández, M.; Calles-Enríquez, M.; Sánchez-Llana, E.; Cañedo, E.; Martín, M.C.; Alvarez, M.A. Is the production of the biogenic amines tyramine and putrescine a species-level trait in enterococci? Food Microbiol. 2012, 30, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Sarantinopoulos, P.; Andrighetto, C.; Georgalaki, M.D.; Rea, M.C.; Lombardi, A.; Cogan, T.M.; Kalantzopoulos, G.; Tsakalidou, E. Biochemical properties of enterococci relevant to their technological performance. Int. Dairy J. 2001, 11, 621–647. [Google Scholar] [CrossRef]
- Chaves-López, C.; De Angelis, M.; Martuscelli, M.; Serio, A.; Paparella, A.; Suzzi, G. Characterization of the Enterobacteriaceae isolated from an artisanal Italian ewe’s cheese (Pecorino Abruzzese). J. Appl. Microbiol. 2006, 101, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Coton, M.; Delbès-Paus, C.; Irlinger, F.; Desmasures, N.; Le Fleche, A.; Stahl, V.; Montel, M.C.; Coton, E. Diversity and assessment of potential risk factors of Gram-negative isolates associated with French cheeses. Food Microbiol. 2011, 29, 88–98. [Google Scholar] [CrossRef]
- Deetae, P.; Mounier, J.; Bonnarme, P.; Spinnler, H.; Irlinger, F.; Helinck, S. Effects of Proteus vulgaris growth on the establishment of a cheese microbial community and on the production of volatile aroma compounds in a model cheese. J. Appl. Microbiol. 2009, 107, 1404–1413. [Google Scholar] [CrossRef]
- Kothe, C.I.; Bolotin, A.; Kraïem, B.F.; Dridi, B.; Renault, P.; Team, F.M. Unraveling the world of halophilic and halotolerant bacteria in cheese by combining cultural, genomic and metagenomic approaches. Int. J. Food Microbiol. 2021, 358, 109312. [Google Scholar] [CrossRef]
- Raymond-Fleury, A.; Lessard, M.H.; Chamberland, J.; Pouliot, Y.; Dugat-Bony, E.; Turgeon, S.L.; St-Gelais, D.; Labrie, S. Analysis of Microbiota Persistence in Quebec’s Terroir Cheese Using a Metabarcoding Approach. Microorganisms 2022, 10, 1381. [Google Scholar] [CrossRef]
- Tornadijo, M.; García, M.; Fresno, J.; Carballo, J. Study of Enterobacteriaceae during the manufacture and ripening of San Simón cheese. Food Microbiol. 2001, 18, 499–509. [Google Scholar] [CrossRef]
- Larpin-Laborde, S.; Imran, M.; Bonaïti, C.; Bora, N.; Gelsomino, R.; Goerges, S.; Irlinger, F.; Goodfellow, M.; Ward, A.C.; Vancanneyt, M.; et al. Surface microbial consortia from Livarot, a French smear-ripened cheese. Can. J. Microbiol. 2011, 57, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Leriche, F.; Bordessoules, A.; Fayolle, K.; Karoui, R.; Laval, K.; Leblanc, L.; Dufour, E. Alteration of raw-milk cheese by Pseudomonas spp.: Monitoring the sources of contamination using fluorescence spectroscopy and metabolic profiling. J. Microbiol. Methods 2004, 59, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Ogier, J.-C.; Lafarge, V.; Girard, V.; Rault, A.; Maladen, V.; Gruss, A.; Leveau, J.-Y.; Delacroix-Buchet, A. Molecular fingerprinting of dairy microbial ecosystems by use of Temporal Temperature and Denaturing Gradient Gel Electrophoresis. Appl. Environ. Microbiol. 2004, 70, 5628–5643. [Google Scholar] [CrossRef] [PubMed]
- Kersters, K.; Vos, P.; Gillis, M.; Swings, J.; Vandamme, P.; Stackebrandt, E. Introduction to the Proteobacteria. In The Prokaryotes; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 3–37. [Google Scholar] [CrossRef]
- O’Hara, C.M.; Brenner, F.W.; Miller, J.M. Classification, identification and clinical significance of Proteus, Providencia and Morganella. Clin. Microbiol. Rev. 2000, 13, 534–546. [Google Scholar] [CrossRef] [PubMed]
- Amador, P.; Fernandes, R.; Prudêncio, C.; Brito, L. Resistance to β-lactams in bacteria isolated from different types of Portuguese cheese. Int. J. Mol. Sci. 2009, 10, 1538–1551. [Google Scholar] [CrossRef]
- Arslan, S.; Özdemir, F. Extended spectrum beta-lactamases in Escherichia coli strains isolated from homemade white cheeses: Prevalence and antibiotic susceptibility. World J. Microbiol. Biotechnol. 2008, 24, 2361–2364. [Google Scholar] [CrossRef]
- Hammad, A.M.; Ishida, Y.; Shimamoto, T. Prevalence and molecular characterization of ampicillin-resistant Enterobacteriaceae isolated from traditional Egyptian Domiati Cheese. J. Food Prot. 2009, 72, 624–630. [Google Scholar] [CrossRef]
- Loizzo, M.R.; Menichini, F.; Picci, N.; Puoci, F.; Spizzirri, U.G.; Restuccia, D. Technological aspects and analytical determination of biogenic amines in cheese. Trends Food Sci. Technol. 2013, 30, 38–55. [Google Scholar] [CrossRef]
- Marino, M.; Maifreni, M.; Moret, S.; Rondinini, G. The capacity of Enterobacteriaceae species to produce biogenic amines in cheese. Lett. Appl. Microbiol. 2000, 31, 169–173. [Google Scholar] [CrossRef]
- Pircher, A.; Bauer, F.; Paulsen, P. Formation of cadaverine, histamine, putrescine and tyramine by bacteria isolated from meat, fermented sausages and cheeses. Eur. Food Res. Technol. 2007, 226, 225–231. [Google Scholar] [CrossRef]
- Deetae, P.; Bonnarme, P.; Spinnler, H.; Helinck, S. Production of volatile aroma compounds by bacterial strains isolated from different surface-ripened French cheeses. Appl. Microbiol. Biotechnol. 2007, 76, 1161–1171. [Google Scholar] [CrossRef] [PubMed]
- Deetae, P.; Spinnler, H.-E.; Bonnarme, P.; Helinck, S. Growth and aroma contribution of Microbacterium foliorum, Proteus vulgaris and Psychrobacter sp. during ripening in a cheese model medium. Appl. Microbiol. Biotechnol. 2009, 82, 169–177. [Google Scholar] [CrossRef]
- Irlinger, F.; In Yung, S.A.Y.; Sarthou, A.-S.; Delbès-Paus, C.; Montel, M.-C.; Coton, E.; Coton, M.; Helinck, S. Ecological and aromatic impact of two Gram-negative bacteria (Psychrobacter celer and Hafnia alvei) inoculated as part of the whole microbial community of an experimental smear soft cheese. Int. J. Food Microbiol. 2012, 153, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, M.; Kodama, K.; Yasuda, H.; Okamoto-Kainuma, A.; Koizumi, K.; Yamasato, K. Presence of halophilic and alkaliphilic lactic acid bacteria in various cheeses. Lett. Appl. Microbiol. 2007, 44, 308–313. [Google Scholar] [CrossRef]
- Schornsteiner, E.; Mann, E.; Bereuter, O.; Wagner, M.; Schmitz-Esser, S. Cultivation-independent analysis of microbial communities on Austrian raw milk hard cheese rinds. Int. J. Food Microbiol. 2014, 180, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Toffin, L.; Zink, K.; Kato, C.; Pignet, P.; Bidault, A.; Bienvenu, N.; Birrien, J.-L.; Prieur, D. Marinilactibacillus piezotolerans sp. nov., a novel marine lactic acid bacterium isolated from deep sub-seafloor sediment of the Nankai Trough. Int. J. Syst. Evol. Microbiol. 2005, 55, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Maoz, A.; Mayr, R.; Scherer, S. Temporal stability and biodiversity of two complex antilisterial cheese-ripening microbial consortia. Appl. Environ. Microbiol. 2003, 69, 4012–4018. [Google Scholar] [CrossRef]
- Bachmann, H.; Bobst, C.; Bütikofer, U.; Casey, M.; Dalla Torre, M.; Fröhlich-Wyder, M.; Fürst, M. Occurrence and significance of Fusarium domesticum alias Anticollanti on smear-ripened cheeses. LWT Food Sci. Technol. 2005, 38, 399–407. [Google Scholar] [CrossRef]
- Hymery, N.; Vasseur, V.; Coton, M.; Mounier, J.; Jany, J.-L.; Barbier, G.; Coton, E. Filamentous fungi and mycotoxins in cheese: A review. Compr. Rev. Food Sci. Food Saf. 2014, 13, 437–456. [Google Scholar] [CrossRef]
- Ropars, J.; Cruaud, C.; Lacoste, S.; Dupont, J. A taxonomic and ecological overview of cheese fungi. Int. J. Food Microbiol. 2012, 155, 199–210. [Google Scholar] [CrossRef]
- Bachmann, H.; Bobst, C.; Buetikofer, U.; Torre, M.; Froehlich-Wyder, M.T.; Furst, M. Sticky cheese smear and natural white mould. Milchwissenschaft 2003, 58, 140–144. [Google Scholar]
- Mounier, J.; Monnet, C.; Vallaeys, T.; Arditi, R.; Sarthou, A.S.; Helias, A.; Irlinger, F. Microbial interactions within a cheese microbial community. Appl. Environ. Microbiol. 2008, 74, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Monnet, C.; Back, A.; Irlinger, F. Growth of aerobic ripening bacteria at the cheese surface is limited by the availability of iron. Appl. Environ. Microbiol. 2012, 78, 3185–3192. [Google Scholar] [CrossRef] [PubMed]
- Cogan, T.M.; Goerges, S.; Gelsomino, R.; Larpin, S.; Hohenegger, M.; Bora, N.; Jamet, E.; Rea, M.C.; Mounier, J.; Vancanneyt, M.; et al. Biodiversity of the Surface Microbial Consortia from Limburger, Reblochon, Livarot, Tilsit, and Gubbeen Cheeses. Microbiol. Spectr. 2014, 2, 219–250. [Google Scholar] [CrossRef] [PubMed]
- Korena, K.; Krzyzankova, M.; Florianova, M.; Karasova, D.; Babak, V.; Strakova, N.; Juricova, H. Microbial Succession in the Cheese Ripening Process-Competition of the Starter Cultures and the Microbiota of the Cheese Plant Environment. Microorganisms 2023, 11, 1735. [Google Scholar] [CrossRef] [PubMed]
- Amato, L.; Ritschard, J.S.; Kurtz, O.; Arias-Roth, E.; Lacroix, C.; Schuppler, M.; Meile, L. Microbial composition of defect smear—A problem evolving during foil-prepacked storage of red-smear cheeses. Int. Dairy J. 2012, 27, 77–85. [Google Scholar] [CrossRef]
- Ritschard, J.S.; Amato, L.; Kumar, Y.; Müller, B.; Meile, L.; Schuppler, M. The role of the surface smear microbiome in the development of defective smear on surface-ripened red-smear cheese. Aims Microbiol. 2018, 4, 621–641. [Google Scholar] [CrossRef]
- Law, B.A. Cheese ripening and cheese flavour technology. In Technology of Cheesemaking; Barry, A.L., Tamime, A.Y., Eds.; Society of Dairy Technology Joined with Wiley-Blackwell: Appleby in Westmorland, UK, 1999; pp. 163–192. [Google Scholar]
- McSweeney, P. Cheese Problems Solved; Elsevier: Amsterdam, The Netherlands, 2007. [Google Scholar]
- Picque, D.; Leclercq-Perlat, M.N.; Guillemin, H.; Cattenoz, T.; Corrieu, G.; Montel, M.C. Impact of packaging on the quality of Saint-Nectaire cheese. Int. Dairy J. 2011, 21, 987–993. [Google Scholar] [CrossRef]
- Imran, M.; Desmasures, N. Safety and Quality Aspects of Smear Ripened Cheeses. In Diversity, Dynamics and Functional Role of Actinomycetes on European Smear Ripened Cheeses; Bora, N., Dodd, C., Desmasures, N., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2015. [Google Scholar] [CrossRef]
- Storari, M.; Kulli, S.; Wüthrich, D.; Bruggmann, R.; Berthoud, H.; Arias-Roth, E. Genomic approach to studying nutritional requirements of Clostridium tyrobutyricum and other Clostridia causing late blowing defects. Food Microbiol. 2016, 59, 213–223. [Google Scholar] [CrossRef]
- Doyle, C.J.; Gleeson, D.; Jordan, K.; Beresford, T.P.; Ross, R.P.; Fitzgerald, G.F.; Cotter, P.D. Anaerobic sporeformers and their significance with respect to milk and dairy products. Int. J. Food Microbiol. 2015, 197, 77–87. [Google Scholar] [CrossRef]
- Tansman, G.F.; Kindstedt, P.S.; Hughest, J.M. Crystallization and demineralization phenomena in washed-rind cheese. J. Dairy Sci. 2017, 100, 8694–8704. [Google Scholar] [CrossRef] [PubMed]
- Dalsgaard, T.K.; Sørensen, J.; Bakman, M.; Nebel, C.; Albrechtsen, R.; Vognsen, L.; Nielsen, J.H. Light-induced protein and lipid oxidation in low-fat cheeses: Whey proteins as antioxidants. Dairy Sci. Technol. 2011, 91, 171–183. [Google Scholar] [CrossRef]
- Juric, M.; Bertelsen, G.; Mortensen, G.; Petersen, M.A. Light-induced colour and aroma changes in sliced, modified atmosphere packaged semi-hard cheeses. Int. Dairy J. 2003, 13, 239–249. [Google Scholar] [CrossRef]
- Kristensen, D.; Orlien, V.; Mortensen, G.; Brockhoff, P.; Skibsted, L.H. Light-induced oxidation in sliced Havarti cheese packaged in modified atmosphere. Int. Dairy J. 2000, 10, 95–103. [Google Scholar] [CrossRef]
- Mortensen, G.; Bertelsen, G.; Mortensen, B.K.; Stapelfeldt, H. Light-induced changes in packaged cheeses—A review. Int. Dairy J. 2004, 14, 85–102. [Google Scholar] [CrossRef]
- Jonnala, B.R.Y.; McSweeney, P.L.H.; Cotter, P.D.; Sheehan, J.J. Recreating pink defect in cheese with different strains of Thermus bacteria. Int. J. Dairy Technol. 2021, 74, 700–708. [Google Scholar] [CrossRef]
- Guzzon, R.; Carafa, I.; Tuohy, K.; Cervantes, G.; Vernetti, L.; Barmaz, A.; Larcher, R.; Franciosi, E. Exploring the microbiota of the red-brown defect in smear-ripened cheese by 454-pyrosequencing and its prevention using different cleaning systems. Food Microbiol. 2017, 62, 160–168. [Google Scholar] [CrossRef]
- Kamelamela, N.; Zalesne, M.; Morimoto, J.; Robbat, A.; Wolfe, B.E. Indigo- and indirubin-producing strains of Proteus and Psychrobacter are associated with purple rind defect in a surface-ripened cheese. Food Microbiol. 2018, 76, 543–552. [Google Scholar] [CrossRef]
- Rodriguez, J.; Lobato, C.; Vazquez, L.; Mayo, B.; Florez, A.B. Prodigiosin-Producing Serratia marcescens as the Causal Agent of a Red Colour Defect in a Blue Cheese. Foods 2023, 12, 2388. [Google Scholar] [CrossRef]
- Di Renzo, T.; Reale, A.; Nazzaro, S.; Siano, F.; Addeo, F.; Picariello, G. Shotgun proteomics for the identification of yeasts responsible for pink/red discoloration in commercial dairy products. Food Res. Int. 2023, 169, 112945. [Google Scholar] [CrossRef]
- Kurtzman, C.; Fell, J.W.; Boekhout, T. The Yeasts: A Taxonomic Study; Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Daly, D.F.M.; McSweeney, P.L.H.; Sheehan, J.J. Pink discolouration defect in commercial cheese: A review. Dairy Sci. Technol. 2012, 92, 439–453. [Google Scholar] [CrossRef]
- Mayo, B.; Rodríguez, J.; Vázquez, L.; Flórez, A.B. Microbial Interactions within the Cheese Ecosystem and Their Application to Improve Quality and Safety. Foods 2021, 10, 602. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, N.M.; O’Connor, T.P.; O’Callaghan, J.; Dobson, A.D.W. Toxins in cheese. In Cheese: Chemistry, Physics and Microbiology; Patrick, F., Fox, P.L.H.M.T.M.C., Timothy, P.G., Eds.; Elsevier: Amsterdam, The Netherlands, 2004; Volume 1, pp. 561–571. [Google Scholar]
- Haasum, I.; Nielsen, P.V. Physiological characterization of common fungi associated with cheese. J. Food Sci. 1998, 63, 157–161. [Google Scholar] [CrossRef]
- Cerqueira, M.A.; Lima, Á.M.; Souza, B.W.S.; Teixeira, J.A.; Moreira, R.A.; Vicente, A.A. Functional polysaccharides as edible coatings for cheese. J. Agric. Food Chem. 2009, 57, 1456–1462. [Google Scholar] [CrossRef] [PubMed]
- Cerqueira, M.A.; Sousa-Gallagher, M.J.; Macedo, I.; Rodriguez-Aguilera, R.; Souza, B.W.S.; Teixeira, J.A.; Vicente, A.A. Use of galactomannan edible coating application and storage temperature for prolonging shelf-life of “Regional” cheese. J. Food Eng. 2010, 97, 87–94. [Google Scholar] [CrossRef]
- Yilmaz, F.; Dagdemir, E. The effects of beeswax coating on quality of Kashar cheese during ripening. Int. J. Food Sci. Technol. 2012, 47, 2582–2589. [Google Scholar] [CrossRef]
- Khoshgozaran, S.; Azizi, M.; Bagheripoor-Fallah, N. Evaluating the effect of modified atmosphere packaging on cheese characteristics: A review. Dairy Sci. Technol. 2012, 92, 1–24. [Google Scholar] [CrossRef]
- Amato, L.; Ritschard, J.S.; Sprecher, I.; Lacroix, C.; Schuppler, M.; Meile, L. Effect of packaging materials, environmental factors and rRNA transcriptional activity of surface microflora on red-smear cheese defect development. Int. Dairy J. 2015, 41, 50–57. [Google Scholar] [CrossRef]


| Soft and Semi-Soft Cheese | Semi-Hard Cheese | Hard Cheese |
|---|---|---|
| Brick (US) | Appenzeller (CH) | Bergkäse (AT, CH, DE) |
| Epoisses (FR) | Danbo (DK) | Comté (FR) |
| Langres (FR) | Havarti (DK) | Esrom (DK) |
| Limburger (DE) | Jurakäse (CH) | Gruyère (CH, FR) |
| Livarot (FR) | Kenhem (NL) | |
| Maroilles (FR) | Mutschli (CH) | Ripened acid curd cheese |
| Mont d’Or (FR) | Raclette (CH, FR) | Harzer, Mainzer (DE) |
| Münster (DE, FR) | Ridder (NO) | Quargel (CZ, DE, AT) |
| Port de Salut (FR) | Salers (FR) | |
| Romadur (DE, BE) | St. Paulin (Suisse) (CH, FR) | Smear-ripened cheese with mold |
| Saint Nectaire (FR) | Tête de Moine (CH) | Chaumes (FR) |
| Trappiste (FR) | Tilsit (CH, DE) | Pont l’Évêque (FR) |
| Vacherin Mont-d’Or (CH) | Vacherin Fribourgeois (CH) | Reblochon (de Savoie) (FR) |
| Winzerkäse (CH) | Taleggio (IT) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ritschard, J.S.; Schuppler, M. The Microbial Diversity on the Surface of Smear-Ripened Cheeses and Its Impact on Cheese Quality and Safety. Foods 2024, 13, 214. https://doi.org/10.3390/foods13020214
Ritschard JS, Schuppler M. The Microbial Diversity on the Surface of Smear-Ripened Cheeses and Its Impact on Cheese Quality and Safety. Foods. 2024; 13(2):214. https://doi.org/10.3390/foods13020214
Chicago/Turabian StyleRitschard, Jasmine S., and Markus Schuppler. 2024. "The Microbial Diversity on the Surface of Smear-Ripened Cheeses and Its Impact on Cheese Quality and Safety" Foods 13, no. 2: 214. https://doi.org/10.3390/foods13020214
APA StyleRitschard, J. S., & Schuppler, M. (2024). The Microbial Diversity on the Surface of Smear-Ripened Cheeses and Its Impact on Cheese Quality and Safety. Foods, 13(2), 214. https://doi.org/10.3390/foods13020214