1. Introduction
Shellfish allergy affects 0.5–2.5% of the global population and is defined as an immune response resulting from exposure to crustacean or molluscan tissues [
1,
2]. Allergens in shellfish are typically muscle-related proteins, such as tropomyosin (TM), arginine kinase (AK) and myosin light chain (MLC), in addition to hemolymph-associated allergens, such as hemocyanin (HC) [
3,
4]. Shellfish allergy has one of the highest rates of food-induced anaphylaxis, affecting 42% of shellfish-allergic adults and 12–20% of children [
5]. There is currently no effective treatment for shellfish allergy; thus, management requires strict elimination of the offending foods [
6]. However, avoiding the consumption of shellfish may not be enough to avoid allergic reactions, as certain allergens in shellfish have been considered cross-reactive with other arthropods, such as dust mites [
7].
In Western food cultures, typically, the tail muscle of crustaceans is consumed, although, in large-clawed crustaceans (e.g., lobsters, crayfish and crabs), the muscle tissue from the claw is also eaten. In eastern cultures (e.g., Asia-Pacific), additional body parts are also consumed (e.g., hepatopancreas and gonads) or used whole in broths and soups [
8]. Research into the differential distribution of allergenicity among crustacean tissues has been poorly investigated within the current literature despite cultural differences in shellfish consumption [
4,
9]. The few studies that have investigated differential tissue allergenicity highlighted that patients may display allergic sensitivity after consuming shrimp cephalothorax tissues but test negative in commercial diagnostic tests using shrimp extracts that are usually based on tail muscle [
9,
10,
11]. For improved clinical diagnosis of shellfish allergy, it is important to understand how different types of tissues differ in diversity and abundance of allergenic proteins. Cooking has also been shown to change the protein composition of tissues by degrading heat-labile proteins. As several allergens are heat-stable, cooking can increase the proportion of heat-stable allergens and thereby increase the allergenicity of tissues consumed [
12]. The effect of heat treatment on crustacean allergens in different tissues, however, has not been fully explored.
The Australian Redclaw crayfish (
Cherax quadricarinatus; termed redclaw hereafter) is a freshwater crayfish endemic to northern Australia. The species has been cultured in Australia since the early 1990s and is now farmed through Southeast Asia, China and South America [
13]. Redclaw is a member of the crustacea order Astacidea, in contrast to the more commonly consumed shrimps and crabs (order Dendrobranchiata and Brachyura, respectively). Despite redclaw being commonly eaten as food, no allergens have been officially characterised from the species, and there are limited studies on the cross-sensitisation between redclaw and other edible shellfish. However, redclaw have been demonstrated to elicit IgE-binding and cross-sensitivity with other related crustacea [
14]. Cross-sensitisation among crustaceans and between crustaceans and molluscs is often observed, with approximately 80–100% of some crustacean allergens being cross-reactive between other crustacean species, in particular the pan-allergen TM [
14]. In contrast, cross-sensitisation between molluscs and crustaceans is often much lower, often less than 20% [
15]. As many allergenic proteins in shellfish are functionally important and evolutionary conserved, redclaw allergens may cross-react and elicit IgE binding from patients allergic to other crustacean species.
Due to a lack of knowledge on the allergenicity of this species, this study characterised the redclaw allergome, determined the distribution and proportion of allergens among three different body parts, and evaluated how cooking affected the heat stability of allergens as well as the allergen proportion. Finally, this study examined the potential of redclaw allergens to cross-react with sera from shrimp-allergic patients.
4. Discussion
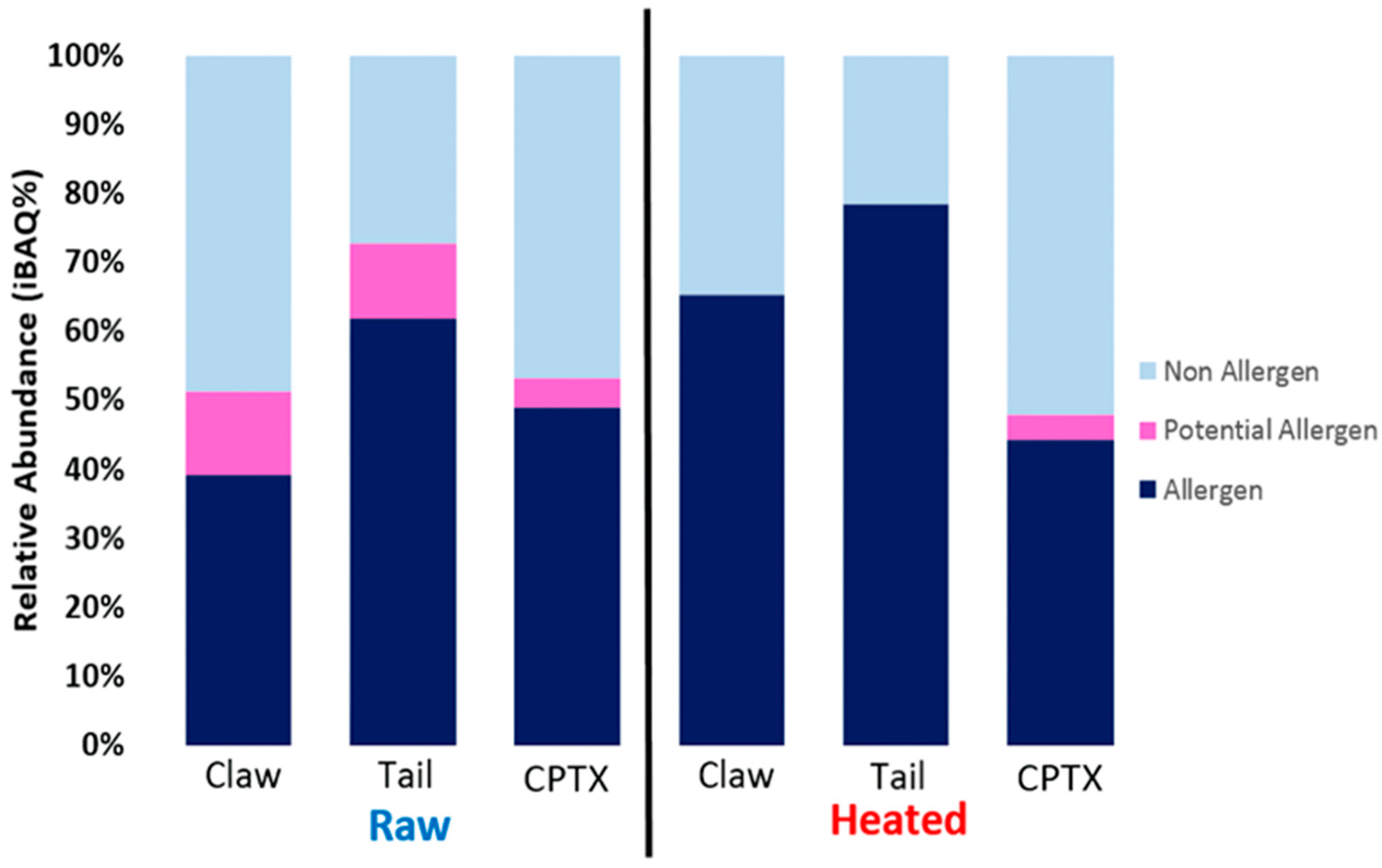
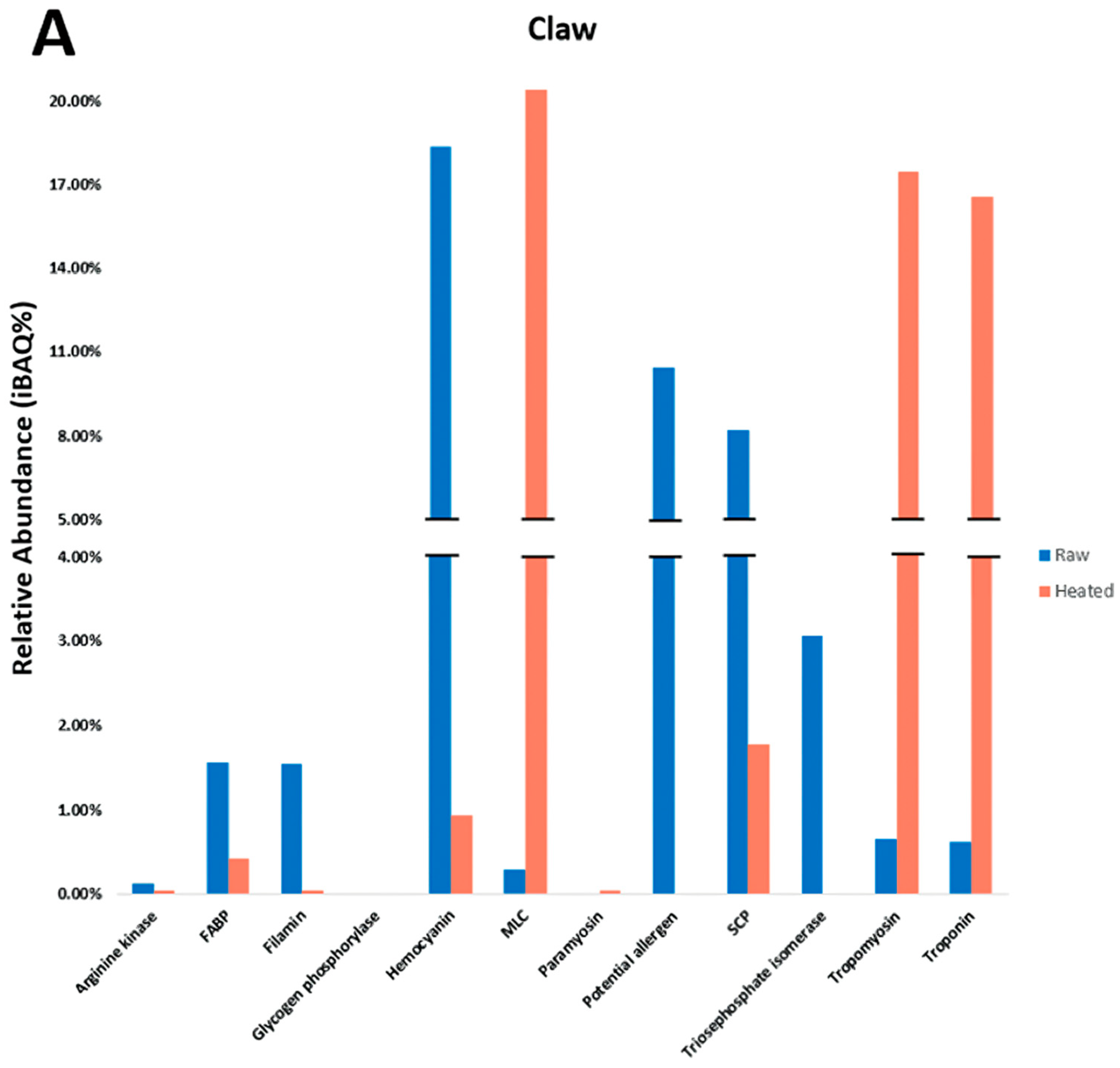
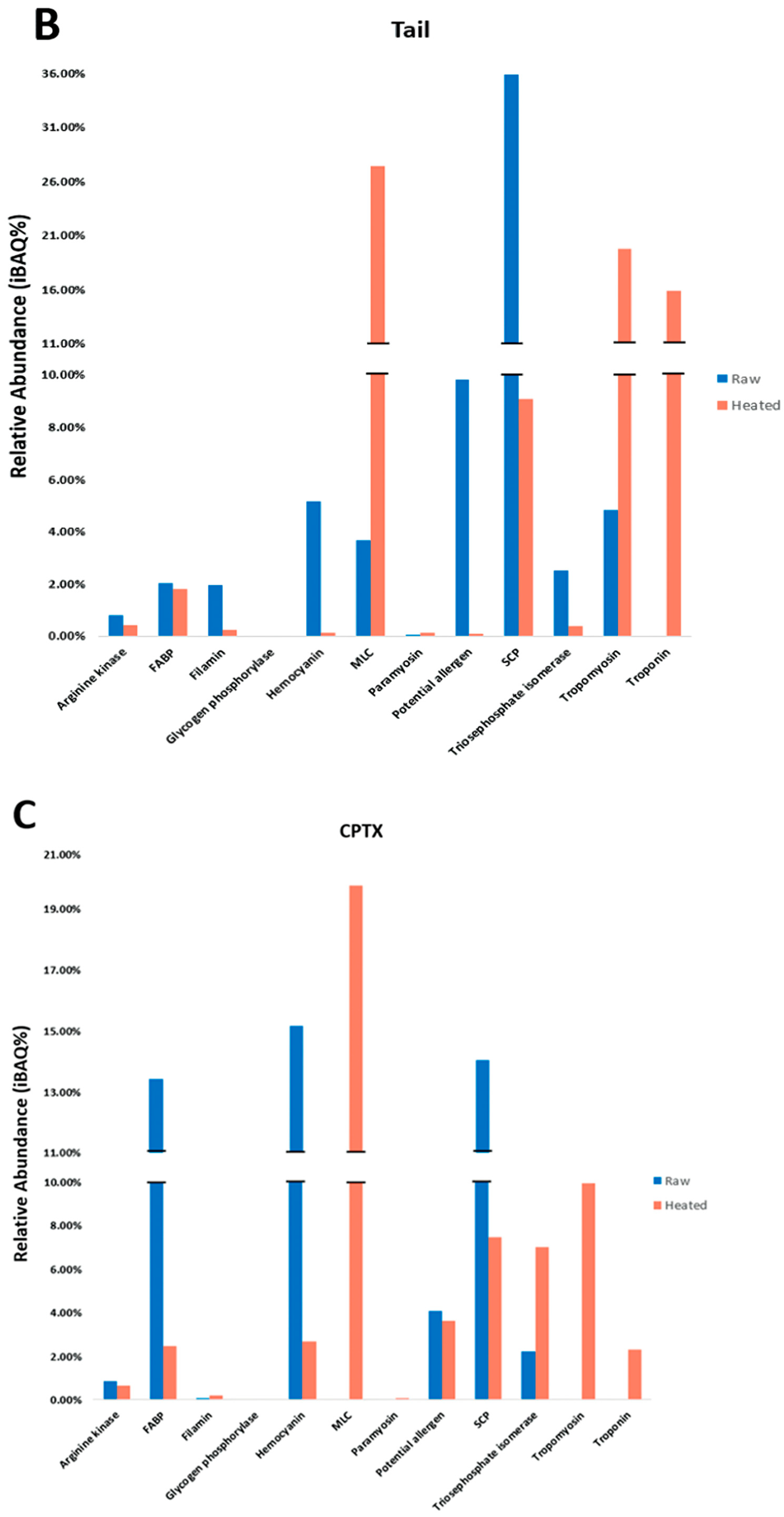
The allergenic differences among various shellfish tissues remain under-investigated. This lack of knowledge relating to tissue-specific allergens may result in missing PAs and hinder the effective diagnosis of a shellfish allergy in patients. In our study, tissue-specific differences in allergens were examined among three body parts and indicated that extracts from the redclaw claw and tail are most similar in allergen content compared to the extracts from the cephalothorax. Key allergens in the claw/tail included HC, MLC, SCP, TM and troponin, as well as other PAs that have evidence of IgE-binding in patients but have not been registered in the WHO/IUIS. In contrast, primary allergens in cephalothorax tissues included FABP, HC, SCP and MLC. Additionally, it was found that the relative abundance (iBAQ%) of predominant allergens varies amongst body parts. Finally, the results from SDS-PAGE, immunoblot and LC/MS indicate that previously described heat-stable allergens are only partially heat-stable, and allergenicity is reduced upon heating. For example, studies into HC regard it as being heat-stable [
25,
26], whereas others suggest that allergenicity is reduced upon heating [
27]. However, reductions are different depending on the tissue type and allergen. The results suggest evidence for allergen isoforms with varying heat stability in the different tissues, although there are limited studies that have investigated the presence of allergen isoforms in shellfish. In those conducted, HC appears to exhibit many isoforms, with 12 isoforms found in tiger shrimp
P. monodon [
11]. HC isoforms display unique sequence diversity and distinctive tissue expression patterns. As a result, this study may suggest that allergen isoforms also have varying allergenicity. However, the differences between allergen isoforms have not been investigated in the context of altered allergenicity. The findings of the previous study are supported by isoform expression studies in crabs (
Cancer magister) and shrimp (
P. monodon) [
28,
29]. Different isoforms were expressed under different conditions (e.g., cold stress, hypoxia, etc.) [
28]. In shellfish, the differences between isoform allergenicity have not been investigated and are required for improved component-resolved diagnosis.
Although studies determining allergenicity between shellfish tissues are limited, the initial evidence of tissue-based differences supports the findings from this current study. It was reported that shellfish-sensitive patients may test negative to conventional IgE tests using shellfish tail muscle extract but have allergic symptoms upon consuming the cephalothorax [
10]. However, when performing a prick-to-prick test using cephalothorax extract instead, patients tested positive [
10]. A subsequent study found that HC in shrimp cephalothorax triggered anaphylaxis in some individuals. Patients with a history of anaphylaxis to shrimp but negative IgE test results underwent prick-to-prick tests using
L. vannamei tail and cephalothorax. The patients tested negative for the tail but positive for the cephalothorax [
9]. Immunoblot analysis using patient sera found two IgE-binding bands at 75 kDa that were identified as HC [
9]. These studies implicate HC as a major allergen in cephalothorax due to its high abundance and potential to cause patient anaphylaxis [
9]. The presence of this HC was confirmed in redclaw, which is taxonomically in a different group to crabs and shrimps.
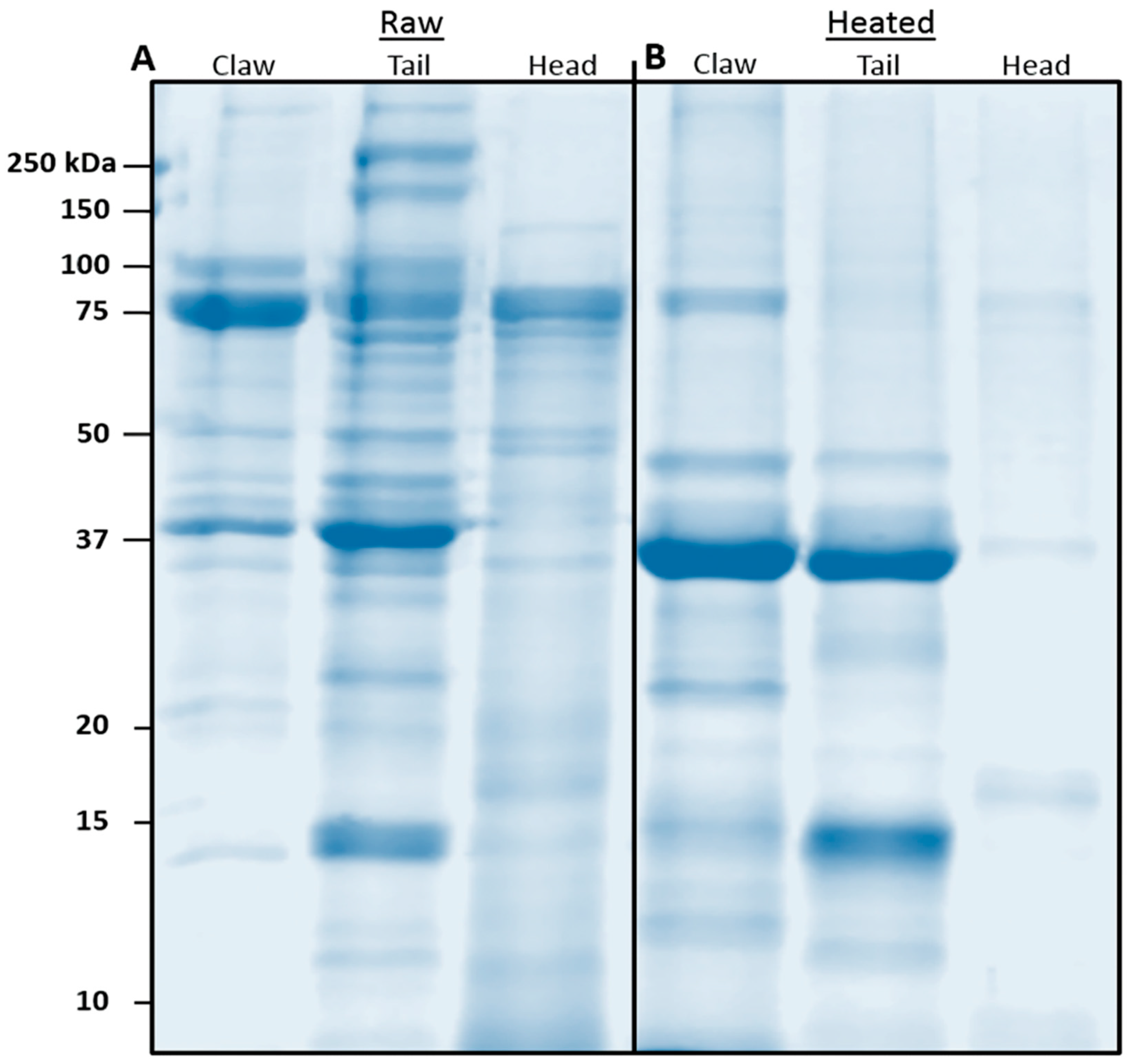
Determining the presence of allergens in raw and heated shellfish is important for consumption and food processing. Heat processing removes many heat-labile proteins but increases the proportion of heat-stable proteins. As detailed in
Section 3.3, centrifugation results in the precipitation of insoluble, denatured proteins and allergens in heated extracts. This results in some differences between the heated tissue and the heated extract, as the extract no longer contains insoluble protein, while it remains in heated tissue. However, a standardised treatment of protein extracts allows for a comparison between raw and heated extracts derived from claw, tail and cephalothorax. Heating typically increases the patient’s recognition of stable allergens and enhances IgE-binding [
9]. In redclaw, TM and MLC were present in raw tissue at relatively low amounts but, upon heating, became predominantly abundant allergens. Antibody detection of TM has previously been shown to increase upon heating in many shellfish species, which may also be due to conformational changes in the secondary structure [
11,
30].
TM, as the most reported major shellfish allergen, remains focal in heat-processing studies [
12], with most other allergenic proteins often overlooked. In this present study, TM, troponin and MLC, proportionally against other proteins, increased when heated, while HC, SCP and PA decreased. However, HC has been described in previous studies to be heat-stable [
25]. In redclaw, the proportion of HC was reduced in all extracts upon heating. However, HC still demonstrated antibody binding upon heating. Similar to HC, SCP is considered to be heat-stable [
31,
32]. However, our results indicated that SCP binding decreases upon heating but is not completely removed. Instead, bands of SCP are found at high molecular weights, which are likely to be SCP aggregates of degraded fragments. SCP aggregates also demonstrate antibody-binding epitopes to the shrimp-specific anti-SCP antibody. This may indicate that perhaps SCP allergenicity decreases upon heating. Other PAs (allergens in similar species) have not been characterised in shellfish; thus, their heat stability is unknown. Research into characterising PAs and determining their allergenicity is required. The current study found that heat stability involves certain allergen isotypes, and some allergens are only partially heat-stable. In our study, redclaw of the same age were sourced from the same hatchery and thereby are standardised for possible protein variations due to age or environmental conditions. Other studies indicate that allergenic protein isoforms may be upregulated or downregulated depending on factors such as age or the moult cycle [
28,
29]. As a result, our current data indicates that in age-matched specimens, there is potential for varying heat stability between allergen isoforms, and many allergens are not strictly heat-stable or labile as previously assumed.
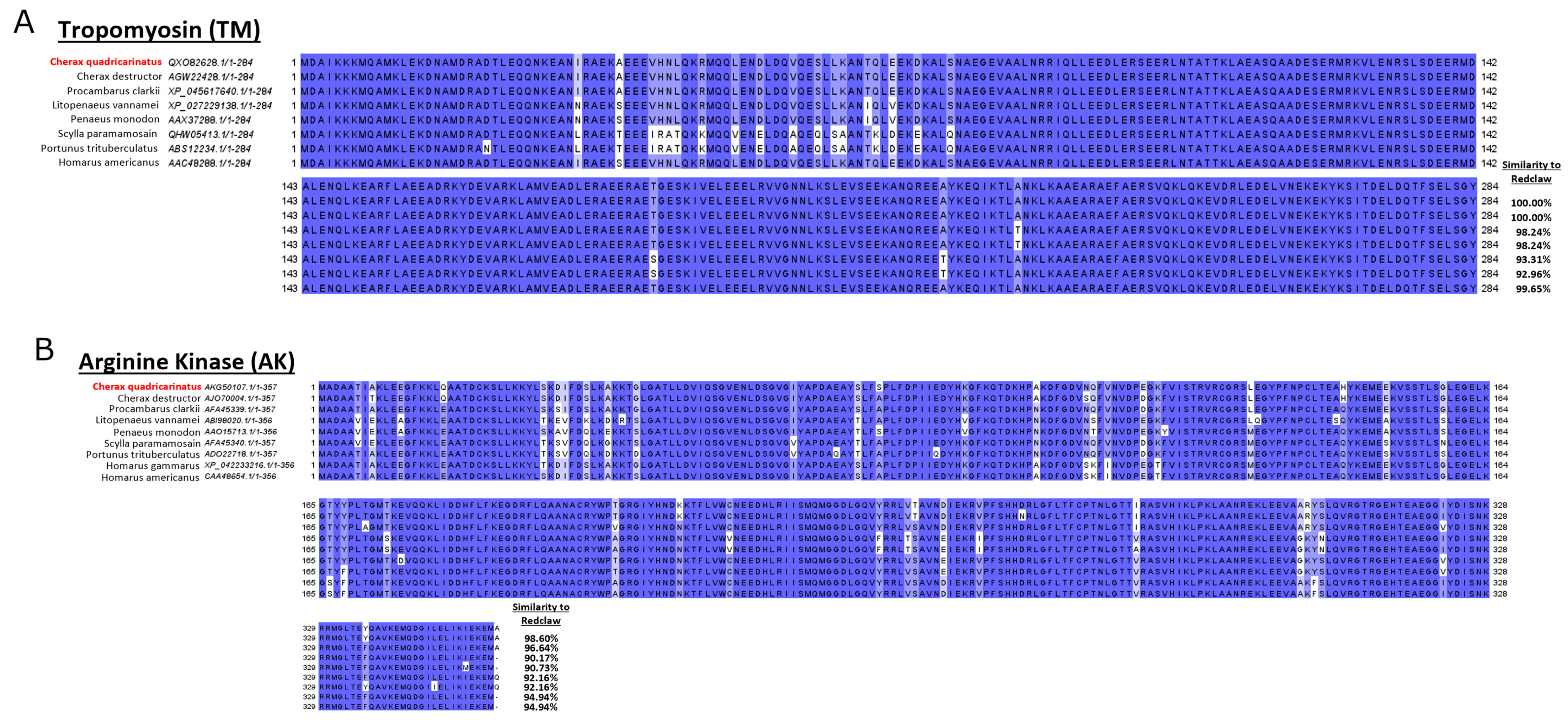
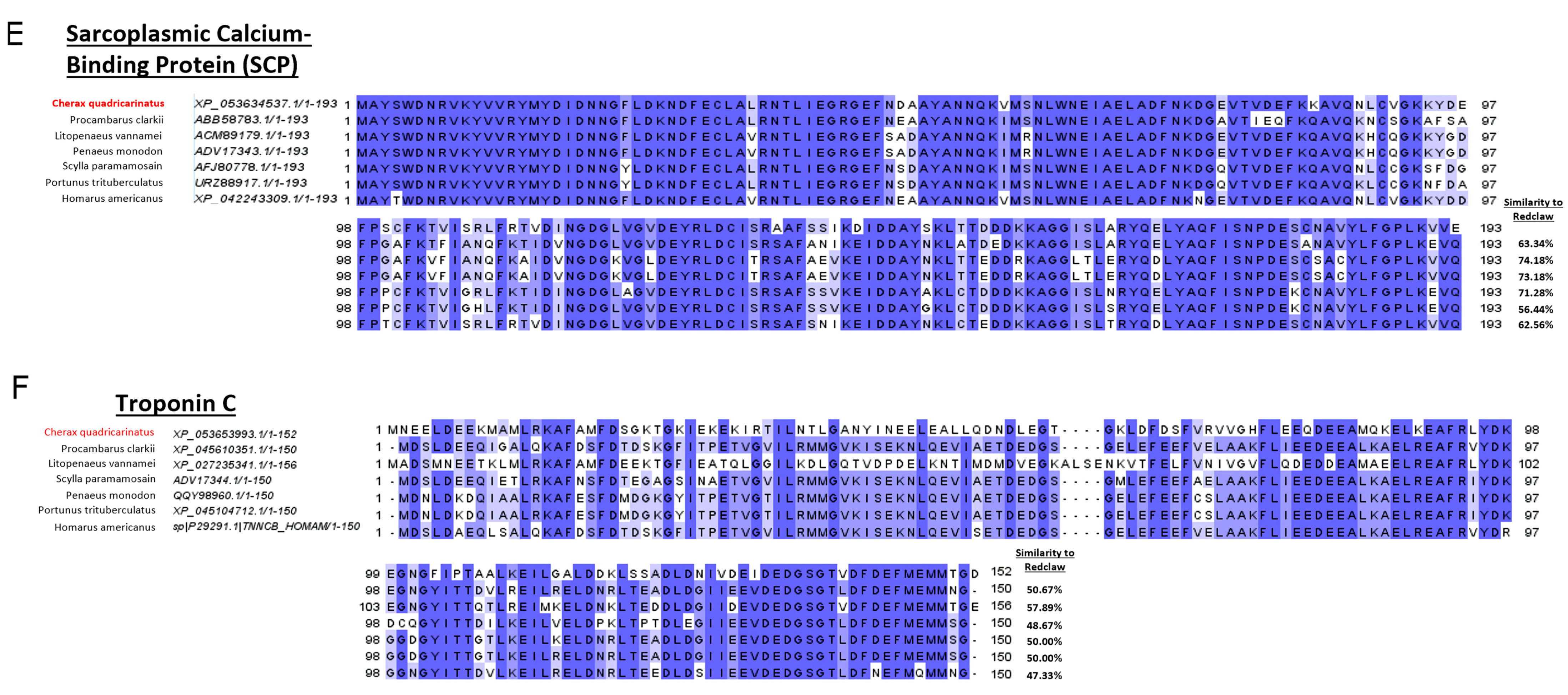
A comparison of amino acid sequences sourced from the protein NCBI database indicates that redclaw TM, AK, HC and SCP are similar (some even identical) to other crustaceans. In contrast, MLC varies depending on the species that redclaw is compared against. For example, redclaw MLC was similar to penaeid shrimp but divergent to crab MLC and even to that of the crayfish
Procambarus clarkii (
Figure 7). However, more research into the similarities between the IgE-binding epitopes of redclaw and other crustacean species is needed to determine if these sequence differences are likely to affect allergenicity. IgE-binding epitopes may be highly conserved regions in the amino acid sequence with little to no sequence changes between species. Specifically, TM, MLC and SCP are all highly cross-reactive proteins between most shellfish and potentially some insect species [
33,
34,
35,
36,
37]. TM is a major invertebrate pan-allergen due to containing high sequence similarities among crustaceans, molluscs, arachnids and insects [
37]. Thus, patients sensitive to TM are likely sensitive to many arthropod species. In contrast, MLC and SCP have been scarcely investigated compared to TM. Shrimp MLC contains high sequence similarities to insects such as the cockroach [
36]. Meanwhile, SCP sequences are similar between shrimp and crayfish species but considerably different from molluscs [
34,
38]. Previous SCP sequence studies indicate that SCP may not be as highly conserved as MLC and TM, which supports the present results, as SCP has a lower sequence similarity between redclaw and shrimp species.
Our study demonstrates that adult and paediatric patients were sensitised to various proteins in different redclaw tissues. MLC and SCP from
L. vannamei were found to be of increased importance in paediatrics, as the frequency of recognition of these allergens is over double that of adults (SCP: 70% vs. 31% in adults, and MLC: 59% vs. 21% in adults). Paediatric patients have also demonstrated increased epitope recognition of many known shrimp allergens. The frequency of allergen recognition is greater in paediatrics, with recognition being almost twice that of adults [
39].
When exposing redclaw extracts to shrimp-sensitive sera, TM was found to be a common IgE-binding protein in both adults and paediatrics. This is reflected in the literature, as adult and paediatric populations were shown to have high recognition frequencies, with up to 61% and 94% to TM from
L. vannamei [
39]. This suggests that TM is associated with the persistence of shellfish allergy into adulthood [
36,
39]. Interestingly, in this present study, adult sera recognised HC (75 kDa) in raw extracts more frequently than in paediatrics. However, three paediatric sera displayed binding to HC. Increased binding in adults may signify that sensitivity and recognition of HC become more prevalent with age. Studies have also found that HC is likely cross-reactive with shellfish and insects, including house dust mites (HDM) and cockroaches. In a study by Kamath et al., 2017 [
40], over 60% of HDM-sensitive infants reacted to HC from Turkish narrow-clawed crayfish (
Astacus leptodactylus). This could indicate that crayfish HC shares cross-reactive epitopes with HDM HC [
40]. In tropical regions (e.g., northern Australia and Southeast Asia), allergic sensitisation is often presented to the tropical mite (
Blomia tropicalis) [
41]. However,
B. tropicalis allergens are still not well characterised.
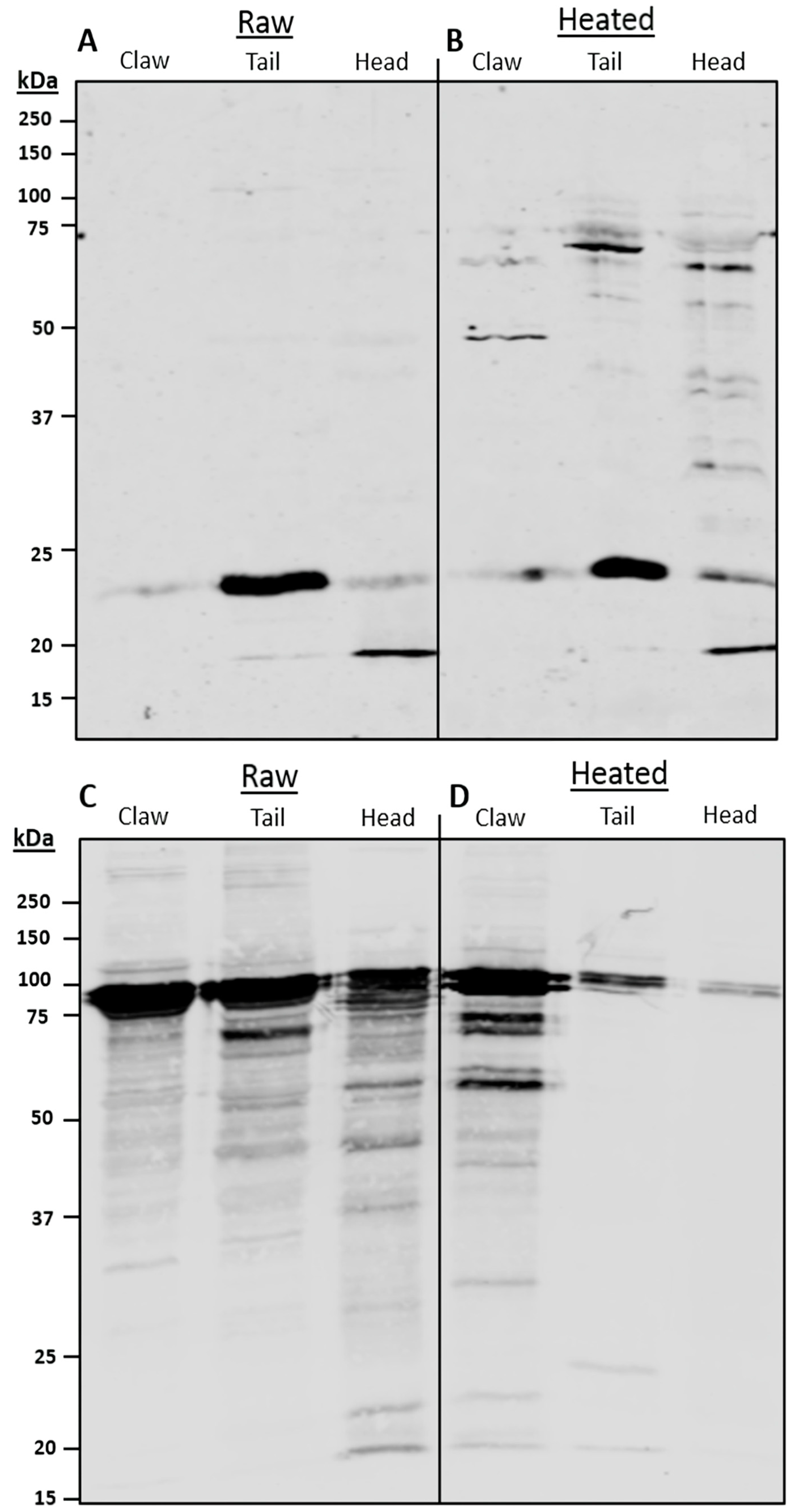
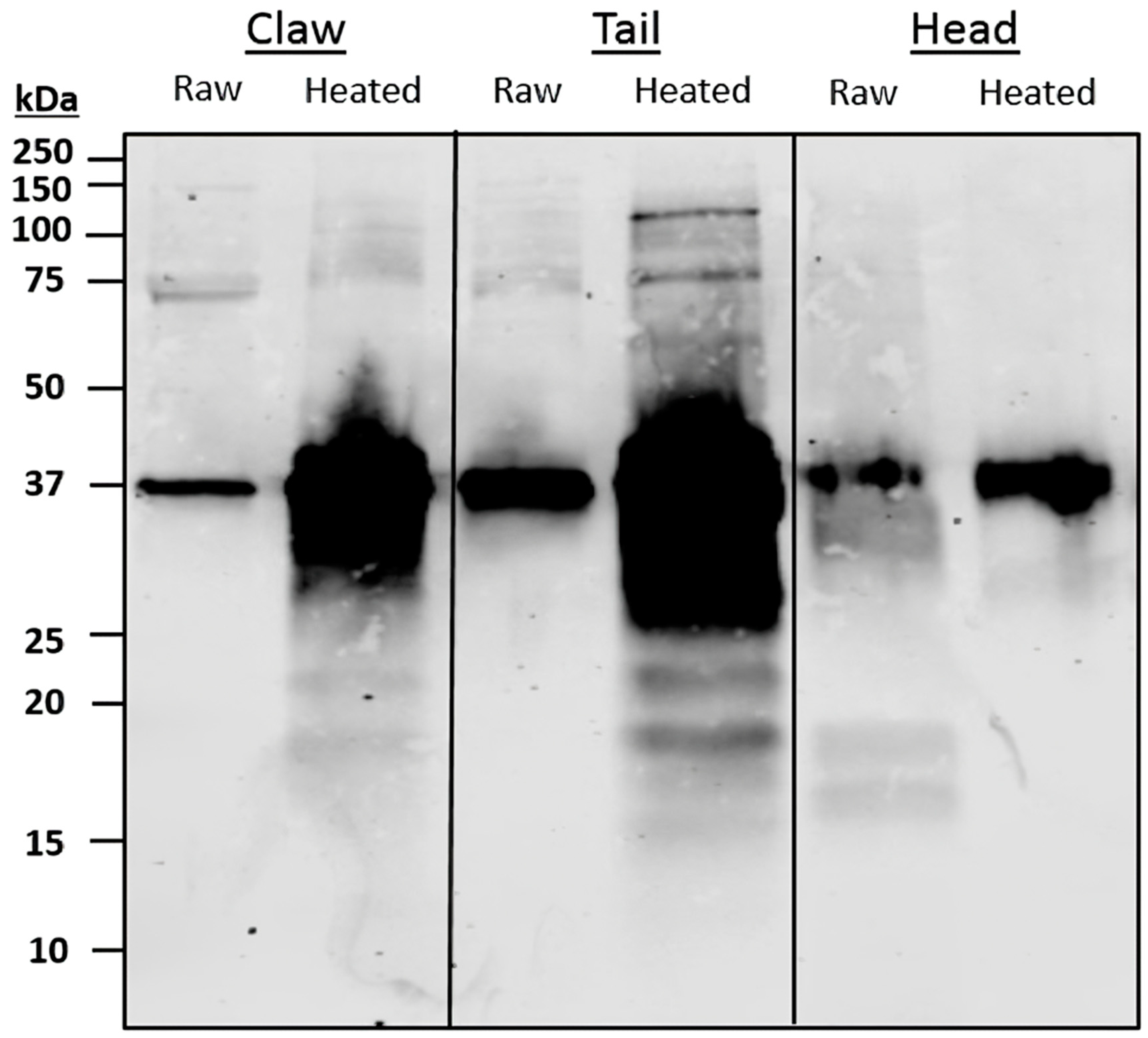
Serum IgE-binding patterns and identified allergens vary among body parts. SDS-PAGE and immunoblot, using pooled sera, display similar binding patterns between claw and tail extracts, with a large region at 37 kDa present in all extracts. The 37 kDa band, identified as TM, was found primarily in the claw and tail, as compared to the cephalothorax. These results support previous findings, as TM is considered a major protein involved in muscle contraction [
37]. Therefore, TM would be more present in the claw and tail, as these body parts comprise primarily fast and slow-contracting muscles. The results from the immunoblot and LC/MS indicate that HC is more abundant in the claw and cephalothorax, with similar relative abundance levels in the 75 kDa bands. These results are supported by the literature, in which HC was found to be synthesised in the hepatopancreas; thus, higher levels of certain HC isoforms were predominantly found in cephalothorax tissue [
42]. The results for claw also indicate a greater presence of IgE-binding to HC. Despite this, there is little information regarding allergenicity and the presence of HC in claw tissue.
The results from the pooled serum immunoblot and LC/MS analysis identified the main IgE-binding proteins. The most allergenic isoform of TM is TM isoform X17, predominantly found at 37 kDa in heated extracts, as well as in lower abundances at 37 kDa in raw extracts. This suggests TM isoform X17 is a protein found in all muscle tissues and seems to be the predominant allergenic form of TM in redclaw. Differences in predominant TM sequences between redclaw, as well as other various crayfish and shrimp species, have been documented [
14]. Redclaw TM IgE-binding epitopes were found to contain no alteration in the conserved regions compared to TM from
Scylla serrata, L. vannamei,
Exopalaemon modestus,
Macrobrachium rosenbergii,
Macrobrachium lanchesteri, and P. clarkii [
14]. This indicates that patients sensitive to other crustaceans are likely sensitive to redclaw, as TM is extremely conserved amongst shellfish [
14].
The LC/MS results indicate that in heated tails, MHC is the most abundant protein in the IgE-binding band. However, MHC is not a registered WHO/IUIS allergen despite being part of the same complex as MLC. Recent studies indicate MHC allergenicity in patients with fish allergy [
43] and increasing evidence in shellfish allergy. MHC is a potential allergen in
P. monodon [
44,
45] and is cross-reactive with HDM MHC [
44]. Studies on banana shrimp (
Fenneropenaeus merguiensis) have also found evidence of MHC allergenicity in shrimp-sensitive patients [
46]. The results from this study may also indicate the allergenicity of MHC. It is worth noting that despite this, this binding may come from the second most abundant protein present in these bands, TM isoform X17. As shown above, TM isoform X17 seems to be a major allergen isoform that causes intense IgE-binding in shrimp-allergic patients. However, inhibition studies are required to determine the protein responsible for this binding.


 ,
, 













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}