
Assessing the Role of Gaseous Chlorine Dioxide in Modulating the Postharvest Ripening of Keitt Mangoes through the Induction of Ethylene Biosynthesis
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mango Fruit
2.2. Procedure and Experimental Design
2.2.1. Gas ClO2 Preparation
2.2.2. Design of the Experiment
2.2.3. Observed Response
2.3. Fresh Weight Loss
2.4. Quality Attributes
2.4.1. Sensory Analysis
2.4.2. Color Measurement
2.4.3. Firmness
2.4.4. Titratable Acidity, Total Soluble Solids, and Total Starch Content
2.4.5. Total Soluble Sugar
2.4.6. Total Carotenoids
2.5. Residual ClO2 Analyses
2.6. Ethylene Production, Respiration Rate, and Abscisic Acid Content
2.7. RNA Sequencing (RNA-Seq) Analysis
2.8. Quantitative Real-Time PCR (RT-qPCR) Analysis
2.9. Statistical Analysis
3. Results
3.1. Physical Quality Characteristics of Mango Fruits
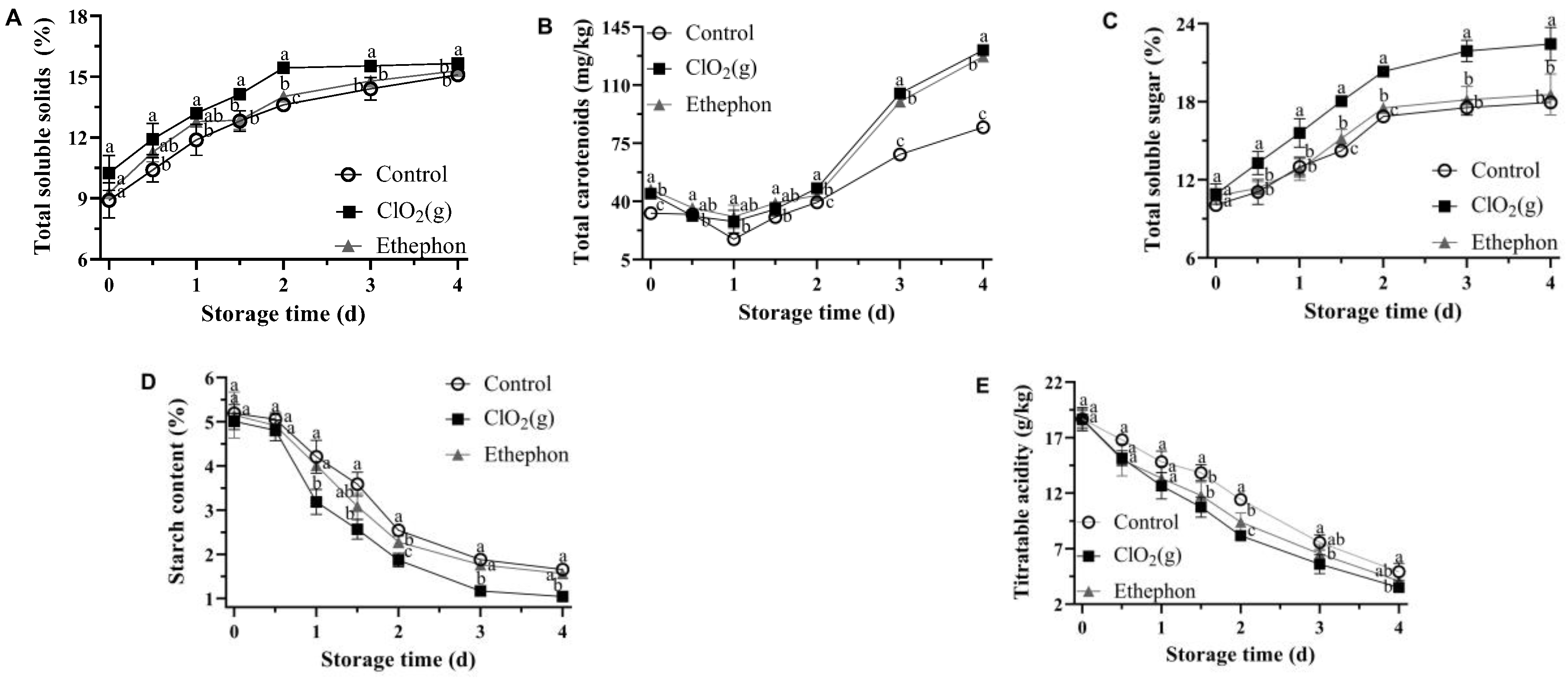
3.2. Biochemical Quality Attributes of Mango Fruits
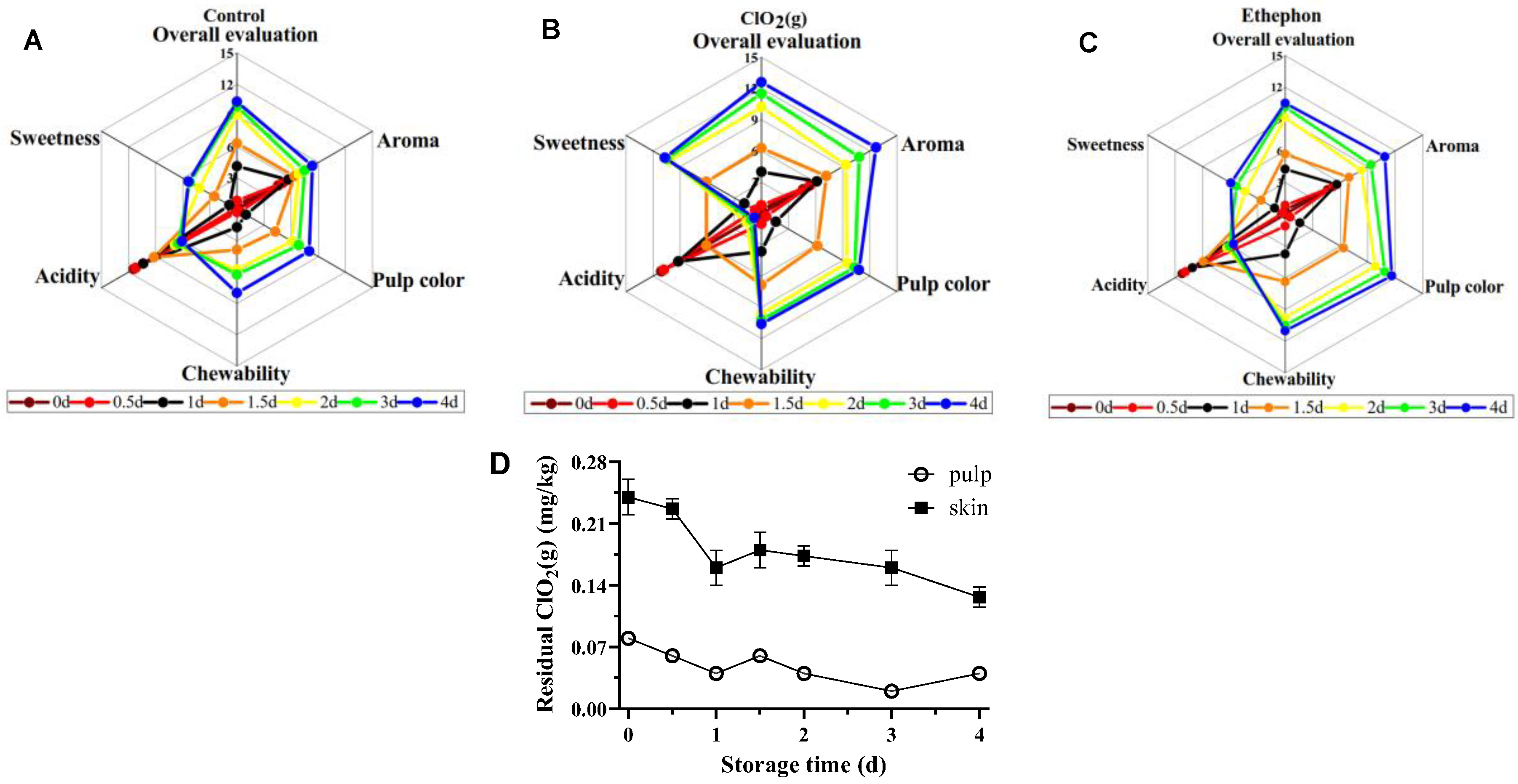
3.3. Sensory Profile and ClO2 Residues of Mangoes Fruits
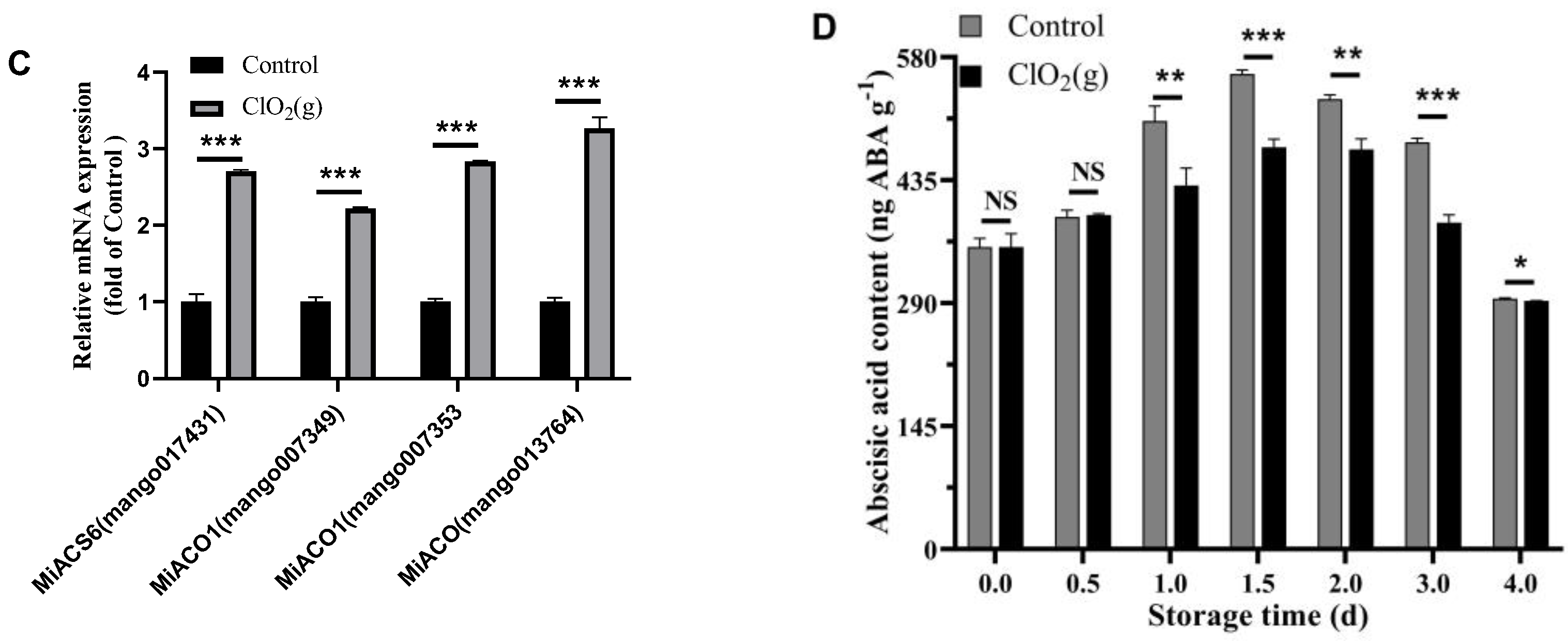
3.4. Respiration Rate and Ethylene Production
3.5. Transcriptomic Profile of Ripe Mango
4. Discussion
5. Conclusions
6. Future Outlook
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Busatto, N.; Vittani, L.; Farneti, B.; Khomenko, I.; Caffini, M.; Faccini, S.; Boschetti, M.; Costa, F. Physiological and molecular characterization of the late ripening stages in Mangifera indica cv Keitt. Postharvest Biol. Technol. 2022, 182, 111746. [Google Scholar] [CrossRef]
- Ali, S.; Akbar Anjum, M.; Sattar Khan, A.; Nawaz, A.; Ejaz, S.; Khaliq, G.; Iqbal, S.; Ullah, S.; Naveed Ur Rehman, R.; Moaaz Ali, M.; et al. Carboxymethyl cellulose coating delays ripening of harvested mango fruits by regulating softening enzymes activities. Food Chem. 2022, 380, 131804. [Google Scholar] [CrossRef] [PubMed]
- Sivakumar, D.; Jiang, Y.; Yahia, E.M. Maintaining mango (Mangifera indica L.) fruit quality during the export chain. Food Rev. Int. 2011, 44, 1254–1263. [Google Scholar] [CrossRef]
- Ma, X.; Zheng, B.; Ma, Y.; Xu, W.; Wu, H.; Wang, S. Carotenoid accumulation and expression of carotenoid biosynthesis genes in mango flesh during fruit development and ripening. Sci. Hortic. 2018, 237, 201–206. [Google Scholar] [CrossRef]
- Baiea, M.H.M.; El-Gioushy, S.F.; El-Badawy, H.E.M. Efficacy of kaolin and screen duo spraying on fruit sunburn, yield and fruit quality of Keitt mango fruits. J. Plant Prod. 2018, 9, 1013–1020. [Google Scholar] [CrossRef]
- Zhang, W.; Dong, P.; Lao, F.; Liu, J.; Liao, X.; Wu, J. Characterization of the major aroma-active compounds in Keitt mango juice: Comparison among fresh, pasteurization and high hydrostatic pressure processing juices. Food Chem. 2019, 289, 215–222. [Google Scholar] [CrossRef]
- Zhou, W. Study on Optimal Fertilization Technology for Improving Quality and Effectiveness of Late Mango of Panzhihua. Master Degree, Hainan University, Haikou, China, 2021; pp. 1–2. [Google Scholar]
- Lebrun, M.; Plotto, A.; Goodner, K.; Ducamp, M.N.; Baldwin, E. Discrimination of mango fruit maturity by volatiles using the electronic nose and gas chromatography. Postharvest Biol. Technol. 2008, 48, 122–131. [Google Scholar] [CrossRef]
- Yan, H.; Wang, R.; Ji, N.; Cao, S.; Ma, C.; Li, J.; Wang, G.; Huang, Y.; Lei, J.; Ba, L. Preparation, Shelf, and Eating Quality of Ready-to-Eat “Guichang” Kiwifruit: Regulation by Ethylene and 1-MCP. Front. Chem. 2022, 10, 934032. [Google Scholar] [CrossRef]
- Zaharah, S.S.; Singh, Z.; Symons, G.M.; Reid, J.B. Role of brassinosteroids, ethylene, abscisic acid, and indole-3-acetic acid in mango fruit ripening. J. Plant Growth Regul. 2012, 31, 363–372. [Google Scholar] [CrossRef]
- Alexander, L.; Grierson, D. Ethylene biosynthesis and action in tomato: A model for climacteric fruit ripening. J. Exp. Bot. 2002, 53, 2039–2055. [Google Scholar] [CrossRef]
- Forlani, S.; Masiero, S.; Mizzotti, C. Fruit ripening: The role of hormones, cell wall modifications, and their relationship with pathogens. J. Exp. Bot. 2019, 70, 2993–3006. [Google Scholar] [CrossRef]
- Shu, P.; Li, Y.; Sheng, J.; Shen, L. SlMAPK3 positively regulates the ethylene production of postharvest tomato fruits and is involved in ethylene-mediated cold tolerance. J. Agric. Food Chem. 2023, 71, 6003–6013. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tang, M.; Liu, M.; Su, D.; Chen, J.; Gao, Y.; Bouzayen, M.; Li, Z. The molecular regulation of ethylene in fruit ripening. Small Methods 2020, 4, 1900485. [Google Scholar] [CrossRef]
- Sabuz, A.A.; Chowdhury, M.G.F.; Molla, M.M.; Khan, M.H.H.; Miaruddin, M. Effect of ethephon on ripening and postharvest quality of mango. Bangladesh J. Agric. Sci. Technol. 2019, 44, 453–467. [Google Scholar] [CrossRef]
- Zhang, Y.; Long, Y.; Liu, Y.; Yang, M.; Wang, L.; Liu, X.; Zhang, Y.; Chen, Q.; Li, M.; Lin, Y.; et al. MAPK5 and MAPK10 overexpression influences strawberry fruit ripening, antioxidant capacity and resistance to Botrytis cinerea. Planta 2021, 255, 19. [Google Scholar] [CrossRef] [PubMed]
- Pilati, S.; Bagagli, G.; Sonego, P.; Moretto, M.; Brazzale, D.; Castorina, G.; Simoni, L.; Tonelli, C.; Guella, G.; Engelen, K.; et al. Abscisic acid is a major regulator of grape berry ripening onset: New insights into ABA signaling network. Front. Plant Sci. 2017, 8, 1093. [Google Scholar] [CrossRef]
- Wu, S.; Wu, D.; Song, J.; Zhang, Y.; Tan, Q.; Yang, T.; Yang, J.; Wang, S.; Xu, J.; Xu, W.; et al. Metabolomic and transcriptomic analyses reveal new insights into the role of abscisic acid in modulating mango fruit ripening. Hort. Res. 2022, 9, uhac102. [Google Scholar] [CrossRef]
- Zaharah, S.S.; Singh, Z.; Symons, G.M.; Reid, J.B. Mode of action of abscisic acid in triggering ethylene biosynthesis and softening during ripening in mango fruit. Postharvest Biol. Technol. 2013, 75, 37–44. [Google Scholar] [CrossRef]
- Adaskaveg, J.A.; Blanco-Ulate, B. Targeting ripening regulators to develop fruit with high quality and extended shelf life. Curr. Opin. Biotechnol. 2023, 79, 102872. [Google Scholar] [CrossRef]
- Zhao, P.; Ndayambaje, J.P.; Liu, X.; Xia, X. Microbial spoilage of fruits: A review on causes and prevention methods. Food Rev. Int. 2022, 38, 225–246. [Google Scholar] [CrossRef]
- Sun, X.; Baldwin, E.; Bai, J. Applications of gaseous chlorine dioxide on postharvest handling and storage of fruits and vegetables—A review. Food Control 2019, 95, 18–26. [Google Scholar] [CrossRef]
- Guo, Q.; Wu, B.; Peng, X.; Wang, J.; Li, Q.; Jin, J.; Ha, Y. Effects of chlorine dioxide treatment on respiration rate and ethylene synthesis of postharvest tomato fruit. Postharvest Biol. Technol. 2014, 93, 9–14. [Google Scholar] [CrossRef]
- Sun, X.; Baldwin, E.; Plotto, A.; Narciso, J.; Ference, C.; Ritenour, M.; Harrison, K.; Gangemi, J.; Bai, J. Controlled-release of chlorine dioxide in a perforated packaging system to extend the storage life and improve the safety of grape tomatoes. J. Vis. Exp. 2017, 122, e55400. [Google Scholar] [CrossRef]
- Jiang, T.; Cheng, C.; Wang, H.; Liu, B.; Zhang, X.; Tian, M.; Li, C.; Fang, T.; Chen, T. Novel gaseous chlorine dioxide treatment system for improving the safety and quality of table grapes during cold storage. LWT 2022, 172, 114232. [Google Scholar] [CrossRef]
- Chi, Z. Study on Inactivation of Inoculated Rot of Pathogens on Grape and Preservation by Chlorine Dioxide Gas. Master Degree, Sun Yat-Sen University, Guangzhou, China, 2010; pp. 48–49. [Google Scholar]
- Silué, Y.; Nindjin, C.; Cissé, M.; Kouamé, K.A.; Amani, N.G.; Mbéguié-A-Mbéguié, D.; Lopez-Lauri, F.; Tano, K. Hexanal application reduces postharvest losses of mango (Mangifera indica L. variety “Kent”) over cold storage whilst maintaining fruit quality. Postharvest Biol. Technol. 2022, 189, 111930. [Google Scholar] [CrossRef]
- Dien, D.C.; Mochizuki, T.; Yamakawa, T. Effect of various drought stresses and subsequent recovery on proline, total soluble sugar and starch metabolisms in Rice (Oryza sativa L.) varieties. Plant Prod. Sci. 2019, 22, 530–545. [Google Scholar] [CrossRef]
- González-Casado, S.; Martín-Belloso, O.; Elez-Martínez, P.; Soliva-Fortuny, R. Enhancing the carotenoid content of tomato fruit with pulsed electric field treatments: Effects on respiratory activity and quality attributes. Postharvest Biol. Technol. 2018, 137, 113–118. [Google Scholar] [CrossRef]
- Tinoco, R.; Hernandez-Saavedra, D.; Ochoa, J.L.; Vazquez-Duhalt, R. Biochemical method for chlorine dioxide determination. Anal. Biochem. 1996, 241, 18–22. [Google Scholar] [CrossRef]
- Razzaq, K.; Singh, Z.; Khan, A.S.; Khan, S.A.K.U.; Ullah, S. Role of 1-MCP in regulating ‘Kensington Pride’ mango fruit softening and ripening. Plant Growth Regul. 2016, 78, 401–411. [Google Scholar] [CrossRef]
- Cárdenas-Barboza, L.C.; Paredes-Córdoba, A.C.; Serna-Cock, L.; Guancha-Chalapud, M.; Torres-León, C. Quality of Physalis peruviana fruits coated with pectin and pectin reinforced with nanocellulose from P. peruviana calyces. Heliyon 2021, 7, e07988. [Google Scholar] [CrossRef]
- Nunes, M.C.N.; Emond, J.P.; Brecht, J.K.; Dea, S.; Proulx, E. Quality curves for mango fruit (cv. Tommy Atkins and Palmer) stored at chilling and nonchilling temperatures. J. Food Qual. 2007, 30, 104–120. [Google Scholar] [CrossRef]
- Mishra, P.; Woltering, E.; Harchioui, N.E. Improved prediction of ‘Kent’ mango firmness during ripening by near-infrared spectroscopy supported by interval partial least square regression. Infrared Phys. Technol. 2020, 110, 103459. [Google Scholar] [CrossRef]
- Ngamchuachit, P.; Sivertsen, H.K.; Mitcham, E.J.; Barrett, D.M. Influence of cultivar and ripeness stage at the time of fresh-cut processing on instrumental and sensory qualities of fresh-cut mangos. Postharvest Biol. Technol. 2015, 106, 11–20. [Google Scholar] [CrossRef]
- Ibarra-Garza, I.P.; Ramos-Parra, P.A.; Hernández-Brenes, C.; Jacobo-Velázquez, D.A. Effects of postharvest ripening on the nutraceutical and physicochemical properties of mango (Mangifera indica L. cv Keitt). Postharvest Biol. Technol. 2015, 103, 45–54. [Google Scholar] [CrossRef]
- Padda, M.S.; Amarante, C.V.T.; Garcia, R.M.; Slaughter, D.C.; Mitcham, E.J. Methods to analyze physico-chemical changes during mango ripening: A multivariate approach. Postharvest Biol. Technol. 2011, 62, 267–274. [Google Scholar] [CrossRef]
- Rosalie, R.; Lechaudel, M.; Dhuique-Mayer, C.; Dufosse, L.; Joas, J. Antioxidant and enzymatic responses to oxidative stress induced by cold temperature storage and ripening in mango (Mangifera indica L. cv. ‘Cogshall’) in relation to carotenoid content. J. Plant Physiol. 2018, 224–225, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Xin, Q.; Zhang, M.; Chen, J.; Lu, Q.; Zhou, X.; Li, X.; Zhang, W.; Feng, W.; Pei, H.; et al. Research progress on mango post-harvest ripening physiology and the regulatory technologies. Foods 2022, 12, 173. [Google Scholar] [CrossRef]
- U.S. Environmental Protection Agency. National Primary Drinking Water Regulations. Available online: https://www.epa.gov/ground-water-and-drinking-water/national-primary-drinking-water-regulations#one (accessed on 22 November 2023).
- Gill, P.P.S.; Jawandha, S.K.; Kaur, N.; Singh, N. Physico-chemical changes during progressive ripening of mango (Mangifera indica L.) cv. Dashehari under different temperature regimes. J. Food Sci. Technol. 2017, 54, 1964–1970. [Google Scholar] [CrossRef]
- Abu, M.; Abbey, L.D.; Amey, N.K. Relation of harvesting time on physicochemical properties of Haden, Kent, Palmer and Keitt mango varieties for export and local markets. J. Hortic. Postharvest Res. 2021, 4, 87–100. [Google Scholar] [CrossRef]
- Maoka, T. Carotenoids as natural functional pigments. J. Nat. Med. 2020, 74, 1–16. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Q.R.; Zhang, M.H.; You, Y.; Wang, Y.; Wang, Y.H. Enhancement of carotenoid biosynthesis in Phaffia rhodozyma PR106 under stress conditions. Biosci. Biotechnol. Biochem. 2019, 83, 2375–2385. [Google Scholar] [CrossRef] [PubMed]
- Vasquez-Caicedo, A.L.; Sruamsiri, P.; Carle, R.; Neidhart, S. Accumulation of all-trans-beta-carotene and its 9-cis and 13-cis stereoisomers during postharvest ripening of nine Thai mango cultivars. J. Agric. Food Chem. 2005, 53, 4827–4835. [Google Scholar] [CrossRef] [PubMed]
- Kyaing, M.S.; Thandar, S.; Myint, M.M.; Wai, K.P.; Htwe, H.T.P.; Nyein, C.M.; Han, J.; Naing, A.H. Characterization of Fruit Quality Traits and Biochemical Properties in Different Myanmar Mango Cultivars during Ripening Stages. Int. J. Plant Biol. 2023, 14, 14–27. [Google Scholar] [CrossRef]
- Wang, H.; Wang, J.; Mujumdar, A.S.; Jin, X.; Liu, Z.L.; Zhang, Y.; Xiao, H.W. Effects of postharvest ripening on physicochemical properties, microstructure, cell wall polysaccharides contents (pectin, hemicellulose, cellulose) and nanostructure of kiwifruit (Actinidia deliciosa). Food Hydrocoll. 2021, 118, 106808. [Google Scholar] [CrossRef]
- Huang, X.Y.; Wang, C.K.; Zhao, Y.W.; Sun, C.H.; Hu, D.G. Mechanisms and regulation of organic acid accumulation in plant vacuoles. Hortic. Res. 2021, 8, 227. [Google Scholar] [CrossRef] [PubMed]
- Medlicott, A.P.; Thompson, A.K. Analysis of sugars and organic acids in ripening of mango fruit (Mangifera indica L. var Keitt) by high-performance liquid chromatography. J. Sci. Food Agric. 1985, 36, 561–566. [Google Scholar] [CrossRef]
- Hernandez-Munoz, P.; Almenar, E.; Valle, V.D.; Velez, D.; Gavara, R. Effect of chitosan coating combined with postharvest calcium treatment on strawberry (Fragariaxananassa) quality during refrigerated storage. Food Chem. 2008, 110, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Lufu, R.; Ambaw, A.; Opara, U.L. Water loss of fresh fruit: Influencing pre-harvest, harvest and postharvest factors. Sci. Hortic. 2020, 272, 109519. [Google Scholar] [CrossRef]
- Robinson, J.E.; Browne, K.M.; Burton, W.G. Storage characteristics of some vegetables and soft fruits. Ann. Appl. Biol. 1975, 81, 399–408. [Google Scholar] [CrossRef]
- Pujol, M.; Garcia-Mas, J. Regulation of climacteric fruit ripening in melon, recent advances and future challenges. J. Exp. Bot. 2023, 74, 6224–6236. [Google Scholar] [CrossRef]
- Bleecker, A.B.; Kende, H. Ethylene: A gaseous signal molecule in plants. Annu. Rev. Cell Dev. Biol. 2000, 16, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Chen, H.; Ma, B.; Zhang, W.; Chen, S.; Zhang, J. The role of ethylene in plants under salinity stress. Front. Plant Sci. 2015, 6, 1059. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Pirrello, J.; Chervin, C.; Roustan, J.P.; Bouzayen, M. Ethylene Control of Fruit Ripening: Revisiting the Complex Network of Transcriptional Regulation. Plant Physiol. 2015, 169, 2380–2390. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Yuan, B.; Leng, P. The role of ABA in triggering ethylene biosynthesis and ripening of tomato fruit. J. Exp. Bot. 2009, 60, 1579–1588. [Google Scholar] [CrossRef]
- Chai, Y.M.; Jia, H.F.; Li, C.L.; Dong, Q.H.; Shen, Y.Y. FaPYR1 is involved in strawberry fruit ripening. J. Exp. Bot. 2011, 62, 5079–5089. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, D.; Liu, B.; Wu, S.; Li, C.; Fang, T.; Tian, M. Assessing the Role of Gaseous Chlorine Dioxide in Modulating the Postharvest Ripening of Keitt Mangoes through the Induction of Ethylene Biosynthesis. Foods 2024, 13, 316. https://doi.org/10.3390/foods13020316
Zhang D, Liu B, Wu S, Li C, Fang T, Tian M. Assessing the Role of Gaseous Chlorine Dioxide in Modulating the Postharvest Ripening of Keitt Mangoes through the Induction of Ethylene Biosynthesis. Foods. 2024; 13(2):316. https://doi.org/10.3390/foods13020316
Chicago/Turabian StyleZhang, Dongwei, Binxiong Liu, Shaoyi Wu, Changcheng Li, Ting Fang, and Meiling Tian. 2024. "Assessing the Role of Gaseous Chlorine Dioxide in Modulating the Postharvest Ripening of Keitt Mangoes through the Induction of Ethylene Biosynthesis" Foods 13, no. 2: 316. https://doi.org/10.3390/foods13020316
APA StyleZhang, D., Liu, B., Wu, S., Li, C., Fang, T., & Tian, M. (2024). Assessing the Role of Gaseous Chlorine Dioxide in Modulating the Postharvest Ripening of Keitt Mangoes through the Induction of Ethylene Biosynthesis. Foods, 13(2), 316. https://doi.org/10.3390/foods13020316