
Improving the Storage Stability of Soy Protein Isolate through Annealing



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Soy Protein Isolate
2.3. Annealing Treatments
2.4. Solubility
2.5. Particle Size Distribution
2.6. Molecular Weight Distribution
2.7. Surface Free and Total Free Sulfhydryl Groups
2.8. Sulfhydryl and Disulfide Content
2.9. Surface Hydrophobicity
2.10. Intrinsic Fluorescence Spectroscopy
2.11. SDS-PAGE Analysis
2.12. Statistical Analysis
3. Results and Discussion
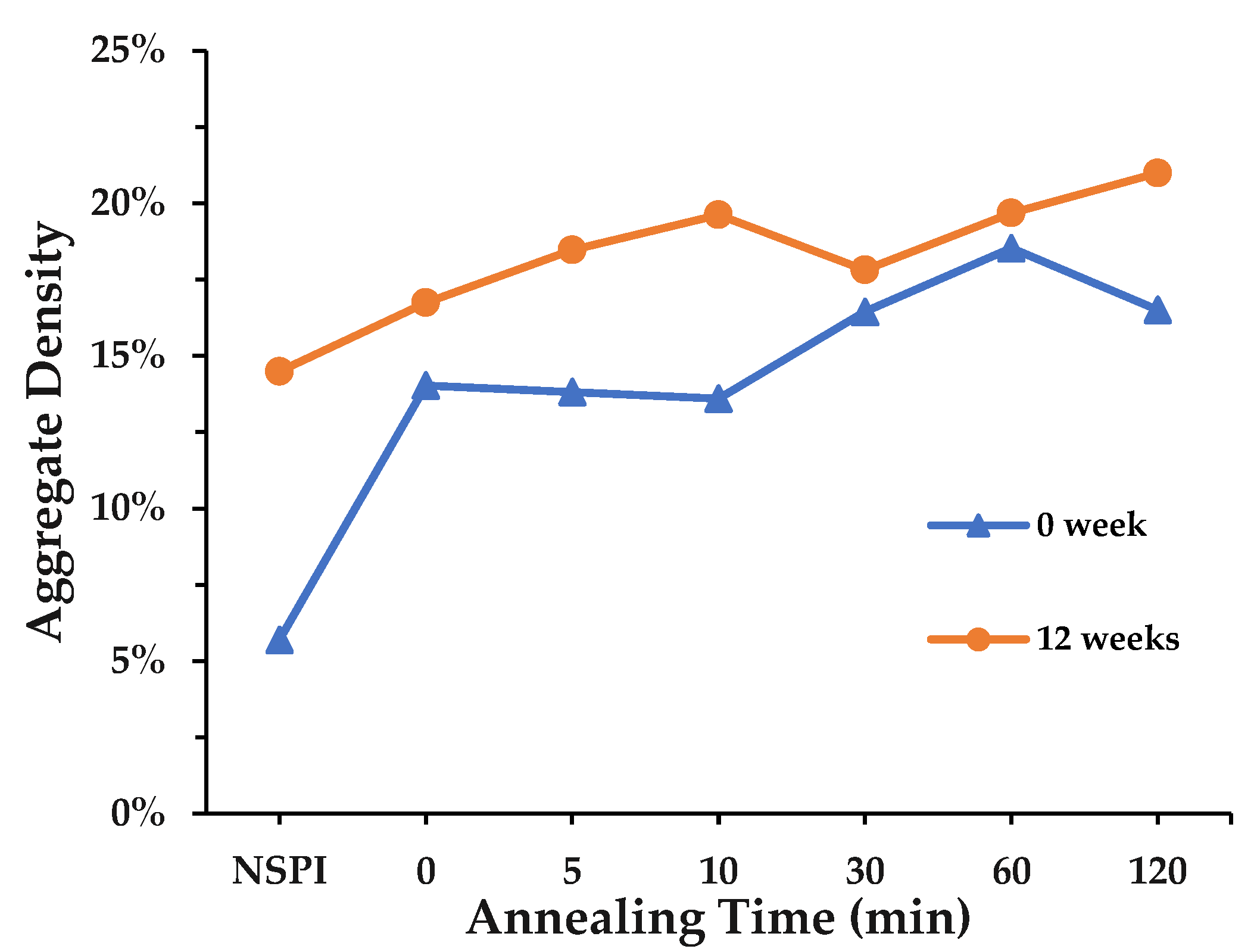
3.1. Solubility
3.2. Particle Size Distribution
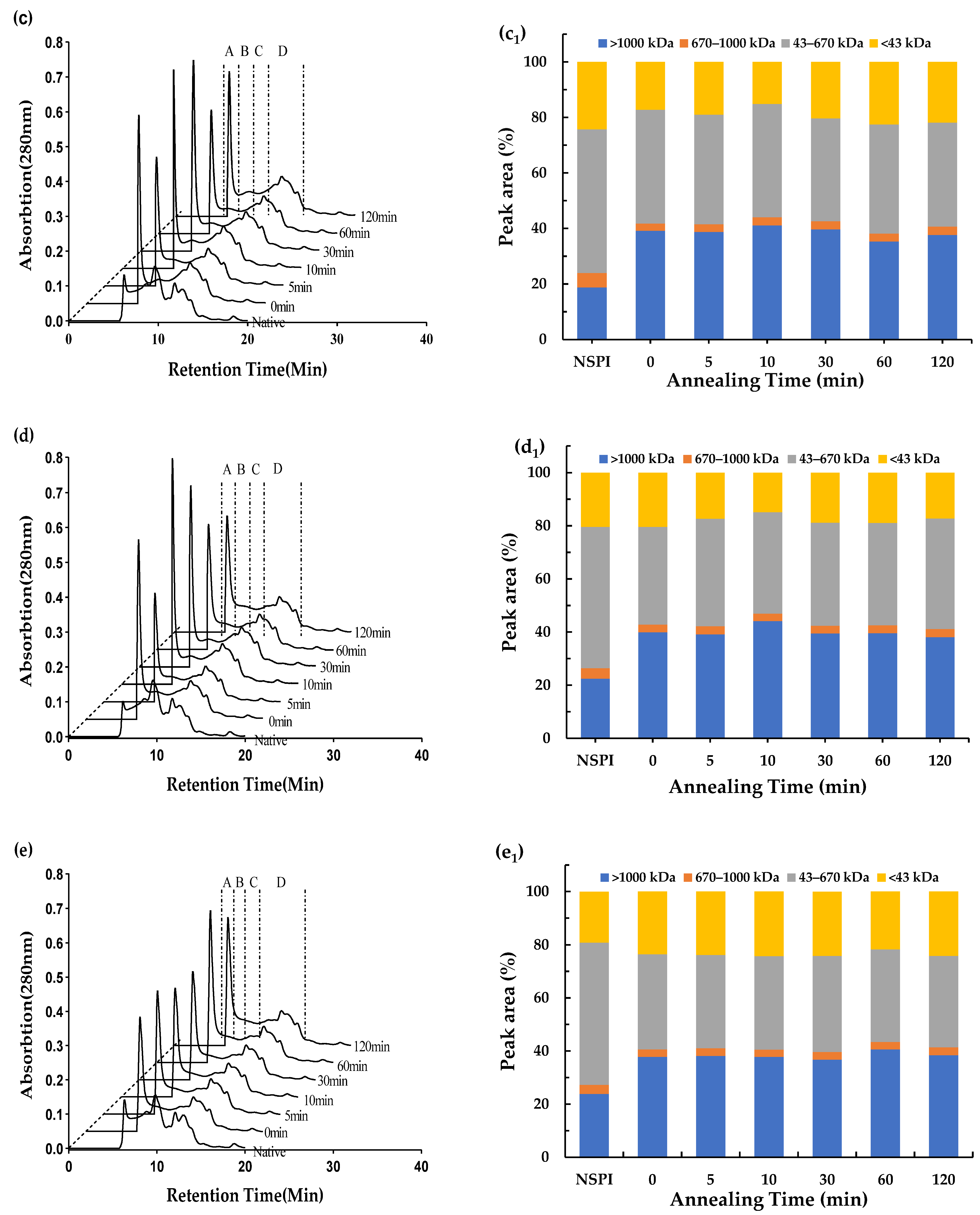
3.3. Molecular Weight Distribution
3.4. Sulfhydryl Groups Contents
3.5. Surface Hydrophobicity
3.6. Intrinsic Fluorescence Spectrum
3.7. SDS–PAGE
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A




References
- Guyony, V.; Fayolle, F.; Jury, V. High moisture extrusion of vegetable proteins for making fibrous meat analogs: A review. Food Rev. Int. 2022, 39, 4262–4287. [Google Scholar] [CrossRef]
- Zhang, Z.Y.; Zhang, L.J.; He, S.D.; Li, X.J.; Jin, R.S.; Liu, Q.; Chen, S.G.; Sun, H.J. High-moisture Extrusion Technology Application in the Processing of Textured Plant Protein Meat Analogues: A Review. Food Rev. Int. 2022, 39, 4873–4908. [Google Scholar] [CrossRef]
- Martins, V.B.; Netto, F.M. Physicochemical and functional properties of soy protein isolate as a function of water activity and storage. Food Res. Int. 2006, 39, 145–153. [Google Scholar] [CrossRef]
- Silva-Pinto, M.d.; Lajolo, F.M.; Genovese, M.I. Effect of storage temperature and water activity on the content and profile of isoflavones, antioxidant activity, and in vitro protein digestibility of soy protein isolates and defatted soy flours. J. Agric. Food Chem. 2005, 53, 6340–6346. [Google Scholar] [CrossRef] [PubMed]
- Hua, Y.F.; Cui, S.W.; Wang, Q.; Mine, Y.; Poysa, V. Heat induced gelling properties of soy protein isolates prepared from different defatted soybean flours. Food Res. Int. 2005, 38, 377–385. [Google Scholar] [CrossRef]
- Feng-Xian, G.; Youling, L.X.; Fang, Q.; Hua-Jun, J.; Xiao-Lin, H.; Jie, C. Examination of the causes of instability of soy protein isolate during storage through probing of the heat-induced aggregation. J. Am. Oil Chem. Soc. 2015, 92, 1075–1084. [Google Scholar] [CrossRef]
- Boatright, W.L.; Hettiarachchy, N.S. Soy protein isolate solubility and surface hydrophobicity as affected by antioxidants. J. Food Sci. 1995, 60, 798–800. [Google Scholar] [CrossRef]
- Fengxian, G.; Luan, L.; Zhiyong, H.; Zong-Ping, Z. Storage stability of soy protein isolate powders containing soluble protein aggregates formed at varying pH. Food Sci. Nutr. 2020, 8, 5275–5283. [Google Scholar] [CrossRef]
- Franzen, K.L.; Kinsella, J.E. Functional properties of succinylated and acetylated soy protein. J. Agric. Food Chem. 1976, 24, 788–795. [Google Scholar] [CrossRef]
- Friedrich, J.P.; Eldridge, A.C. Production of Defatted Soybean Products by Supercritical Fluid Extraction. U.S. Patent 4 493 854, 1 February 1989. [Google Scholar]
- Garner, F.A.; Bates, J.F.; Mitchell, M.A. The strong influence of temper annealing conditions on the neutron-induced swelling of cold-worked austenitic steels. J. Nucl. Mater. 1992, 189, 201–209. [Google Scholar] [CrossRef]
- Coules, H.E. Contemporary approaches to reducing weld induced residual stress. Mater. Sci. Technol. 2013, 29, 4–18. [Google Scholar] [CrossRef]
- Xiong, X.S.; Hu, S.B.; Dang, N.Y.; Hu, K. Effect of stress-relief annealing on microstructure, texture and hysteresis curve of mechanically cut non-oriented Fe-Si steel. Mater. Charact. 2017, 132, 239–247. [Google Scholar] [CrossRef]
- Zhou, X.; Feng, J.C.; Yi, J.J.; Wang, L. Synergistic improvement of toughness of isotactic polypropylene: The introduction of high density polyethylene and annealing treatment. Mater. Des. 2013, 49, 502–510. [Google Scholar] [CrossRef]
- Chu, X.F.; Su, X.; Liu, M.D.; Li, L.; Li, T.H.; Qin, Y.C.; Lu, G.L.; Qi, L.; Liu, Y.H.; Lin, J.Z.; et al. Annealing synchronizes the 70S ribosome into a minimum-energy conformation. Proc. Natl. Acad. Sci. USA 2022, 119, e2111231119. [Google Scholar] [CrossRef] [PubMed]
- Su, C.Y.; Saleh, A.S.M.; Zhang, B.; Zhao, K.; Ge, X.Z.; Zhang, Q.; Li, W.H. Changes in structural, physicochemical, and digestive properties of normal and waxy wheat starch during repeated and continuous annealing. Carbohydr. Polym. 2020, 247, 116675. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.W.; Wang, Z.J.; He, Z.Y.; Zeng, M.M.; Qin, F.; Chen, J. Effects of soluble aggregates sizes on rheological properties of soybean protein isolate under high temperature. LWT-Food Sci. Technol. 2023, 182, 10. [Google Scholar] [CrossRef]
- Beveridge, T.; Toma, S.J.; Nakai, S. Determination of SH- and SS-groups in some food proteins using Ellman’s reagent. J. Food Sci. 1974, 39, 49–51. [Google Scholar] [CrossRef]
- Tang, C.; Ma, C. Effect of high pressure treatment on aggregation and structural properties of soy protein isolate. LWT-Food Sci. Technol. 2009, 42, 606–611. [Google Scholar] [CrossRef]
- Hu, H.; Li-Chan, E.C.Y.; Wan, L.; Tian, M.; Pan, S.Y. The effect of high intensity ultrasonic pre-treatment on the properties of soybean protein isolate gel induced by calcium sulfate. Food Hydrocoll. 2013, 32, 303–311. [Google Scholar] [CrossRef]
- Xu, X.; Liu, W.; Liu, C.; Luo, L.; Chen, J.; Luo, S.; McClements, D.J.; Wu, L. Effect of limited enzymatic hydrolysis on structure and emulsifying properties of rice glutelin. Food Hydrocoll. 2016, 61, 251–260. [Google Scholar] [CrossRef]
- Zhang, W.X.; Boateng, I.D.; Zhang, W.J.; Jia, S.F.; Wang, T.T.; Huang, L.R. Effect of ultrasound-assisted ionic liquid pretreatment on the structure and interfacial properties of soy protein isolate. Process Biochem. 2022, 115, 160–168. [Google Scholar] [CrossRef]
- Nik, A.M.; Tosh, S.M.; Woodrow, L.; Poysa, V.; Corredig, M. Effect of soy protein subunit composition and processing conditions on stability and particle size distribution of soymilk. LWT-Food Sci. Technol. 2009, 42, 1245–1252. [Google Scholar] [CrossRef]
- Zheng, Y.; Sun, M.; Sun, X.; Sun, C.; Fang, Y. The role of protein concentration in heat-induced particulation of soy proteins at different pHs: Structure and functional properties. Food Front. 2023, 4, 955–965. [Google Scholar] [CrossRef]
- He, Z.; Li, W.; Guo, F.; Li, W.; Zeng, M.; Chen, J. Foaming characteristics of commercial soy protein isolate as influenced by heat-induced aggregation. Int. J. Food Prop. 2015, 18, 1817–1828. [Google Scholar] [CrossRef]
- Shen, L.; Tang, C.H. Microfluidization as a potential technique to modify surface properties of soy protein isolate. Food Res. Int. 2012, 48, 108–118. [Google Scholar] [CrossRef]
- Xian’e, R.; Chunzhi, L.; Feng, Y.; Yongchun, H.; Chengdu, H.; Kunming, Z.; Liujuan, Y. Comparison of hydrodynamic and ultrasonic cavitation effects on soy protein isolate functionality. J. Food Eng. 2020, 265, 109697. [Google Scholar] [CrossRef]
- Remondetto, G.E.; Gonzalez, R.J.; Anon, M.C. Effects of simultaneous heat and reducing treatments on some structural characteristics of soy isolates obtained at pilot plant scale. Food Sci. Technol. Int. 2002, 8, 223–228. [Google Scholar] [CrossRef]
- Chuan-He, T. Thermal denaturation and gelation of vicilin-rich protein isolates from three Phaseolus legumes: A comparative study. LWT-Food Sci. Technol. 2008, 41, 1380–1388. [Google Scholar] [CrossRef]
- Wang, H.; Wang, N.; Chen, X.; Wu, Z.A.; Zhong, W.Y.; Yu, D.Y.; Zhang, H.W. Effects of moderate electric field on the structural properties and aggregation characteristics of soybean protein isolate. Food Hydrocoll. 2022, 133. [Google Scholar] [CrossRef]
- Wang, M.; Chen, X.; Guo, J.; Yang, J.; Wang, J.; Yang, X. Stabilization of foam and emulsion by subcritical water-treated soy protein: Effect of aggregation state. Food Hydrocoll. 2019, 87, 619–628. [Google Scholar] [CrossRef]
- Wagner, J.R.; Sorgentini, D.A.; Añón, M.C. Relation between solubility and surface hydrophobicity as an indicator of modifications during preparation processes of commercial and laboratory-prepared soy protein isolates. J. Agric. Food Chem. 2000, 48, 3159–3165. [Google Scholar] [CrossRef]
- Qi, B.; Zhao, C.; Jiang, L.; Wang, Z.; Li, Y. Effect of composition and secondary structure of soybean protein isolate on surface hydrophobicity. J. Chin. Inst. Food Sci. Technol. 2018, 18, 287–293. [Google Scholar] [CrossRef]
- Guo, Z.; Huang, Z.; Guo, Y.; Li, B.; Yu, W.; Zhou, L.; Jiang, L.; Teng, F.; Wang, Z. Effects of high-pressure homogenization on structural and emulsifying properties of thermally soluble aggregated kidney bean (Phaseolus vulgaris L.) proteins. Food Hydrocoll. 2021, 119, 106835. [Google Scholar] [CrossRef]
- Vetri, V.; Militello, V. Thermal induced conformational changes involved in the aggregation pathways of beta-lactoglobulin. Biophys. Chem. 2005, 113, 83–91. [Google Scholar] [CrossRef]
- Ye, Z.; Pingping, X.; Haihong, W.; Muhan, Z.; Zhilan, S.; Chong, S.; Daoying, W.; Jinxuan, C.; Weimin, X. Effects of different ultrasound power on physicochemical property and functional performance of chicken actomyosin. Int. J. Biol. Macromol. 2018, 113, 640–647. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhong, Q.X. Effects of Thermal Denaturation on Binding between Bixin and Whey Protein. J. Agric. Food Chem. 2012, 60, 7526–7531. [Google Scholar] [CrossRef] [PubMed]
- Vivian, J.T.; Callis, P.R. Mechanisms of tryptophan fluorescence shifts in proteins. Biophys. J. 2001, 80, 2093–2109. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.C.; Wang, T.; Wang, J.M.; Wu, D.; Wu, C.; Du, M. Enhancing the thermal stability of soy proteins by preheat treatment at lower protein Concentration (vol 306, 125593, 2020). Food Chem. 2020, 311, 125883. [Google Scholar] [CrossRef] [PubMed]
- Nishinari, K.; Fang, Y.; Guo, S.; Phillips, G.O. Soy proteins: A review on composition, aggregation and emulsification. Food Hydrocoll. 2014, 39, 301–318. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, S.; Wang, Z.; Zeng, M.; He, Z.; Chen, J. Improving the Storage Stability of Soy Protein Isolate through Annealing. Foods 2024, 13, 615. https://doi.org/10.3390/foods13040615
Zou S, Wang Z, Zeng M, He Z, Chen J. Improving the Storage Stability of Soy Protein Isolate through Annealing. Foods. 2024; 13(4):615. https://doi.org/10.3390/foods13040615
Chicago/Turabian StyleZou, Shenzhong, Zhaojun Wang, Maomao Zeng, Zhiyong He, and Jie Chen. 2024. "Improving the Storage Stability of Soy Protein Isolate through Annealing" Foods 13, no. 4: 615. https://doi.org/10.3390/foods13040615
APA StyleZou, S., Wang, Z., Zeng, M., He, Z., & Chen, J. (2024). Improving the Storage Stability of Soy Protein Isolate through Annealing. Foods, 13(4), 615. https://doi.org/10.3390/foods13040615