
Identification of Characteristic Bioactive Compounds in Silkie Chickens, Their Effects on Meat Quality, and Their Gene Regulatory Network

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statements
2.2. Birds and Breast Muscle Sample Collection
2.3. Nutritional Value
2.3.1. Metabolomic Analysis
2.3.2. Lipidomic Analysis
2.3.3. Melanin Content Measurement
2.4. Sensory Characteristics
2.4.1. pH Value and Meat Color
2.4.2. Water-Holding Capacity
2.4.3. Shear Force
2.4.4. Electronic Tongue Analysis
2.4.5. Volatile Flavor Profile Analysis
2.5. Transcriptome Analysis
2.6. Statistical Analysis
3. Results and Discussion
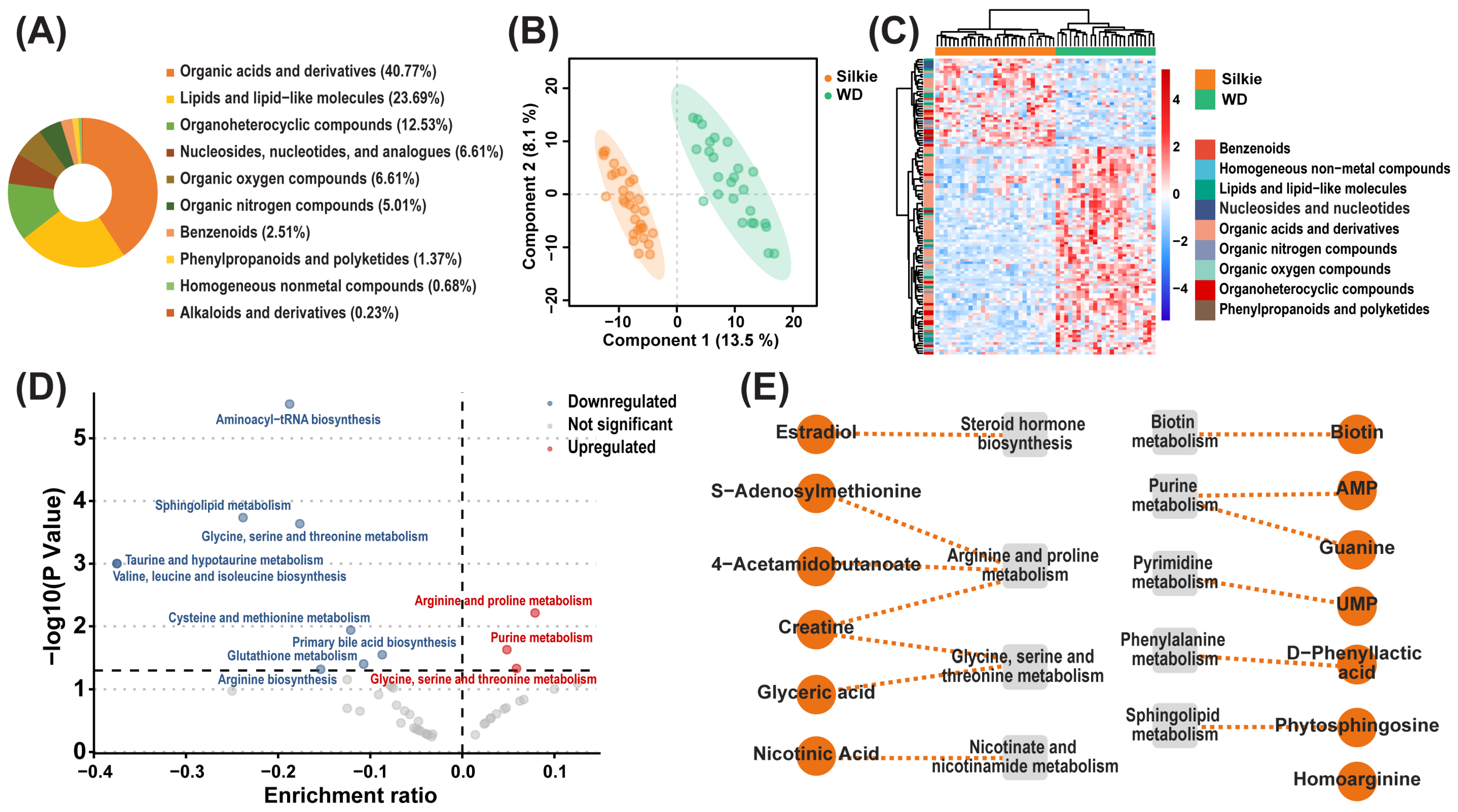
3.1. Identification of Characteristic Bioactive Metabolites
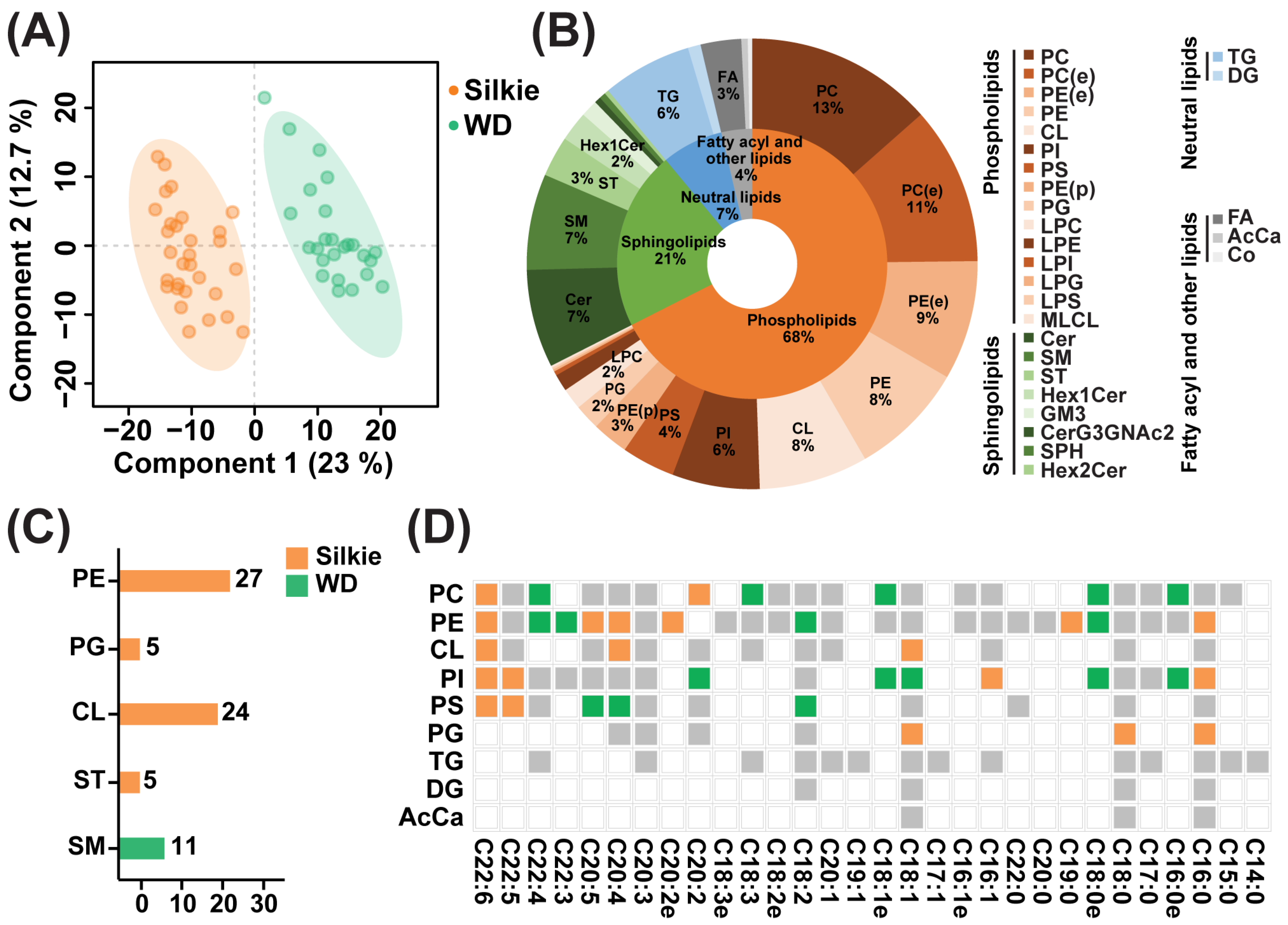
3.2. Identification of Characteristic Bioactive Lipids and Fatty Acids
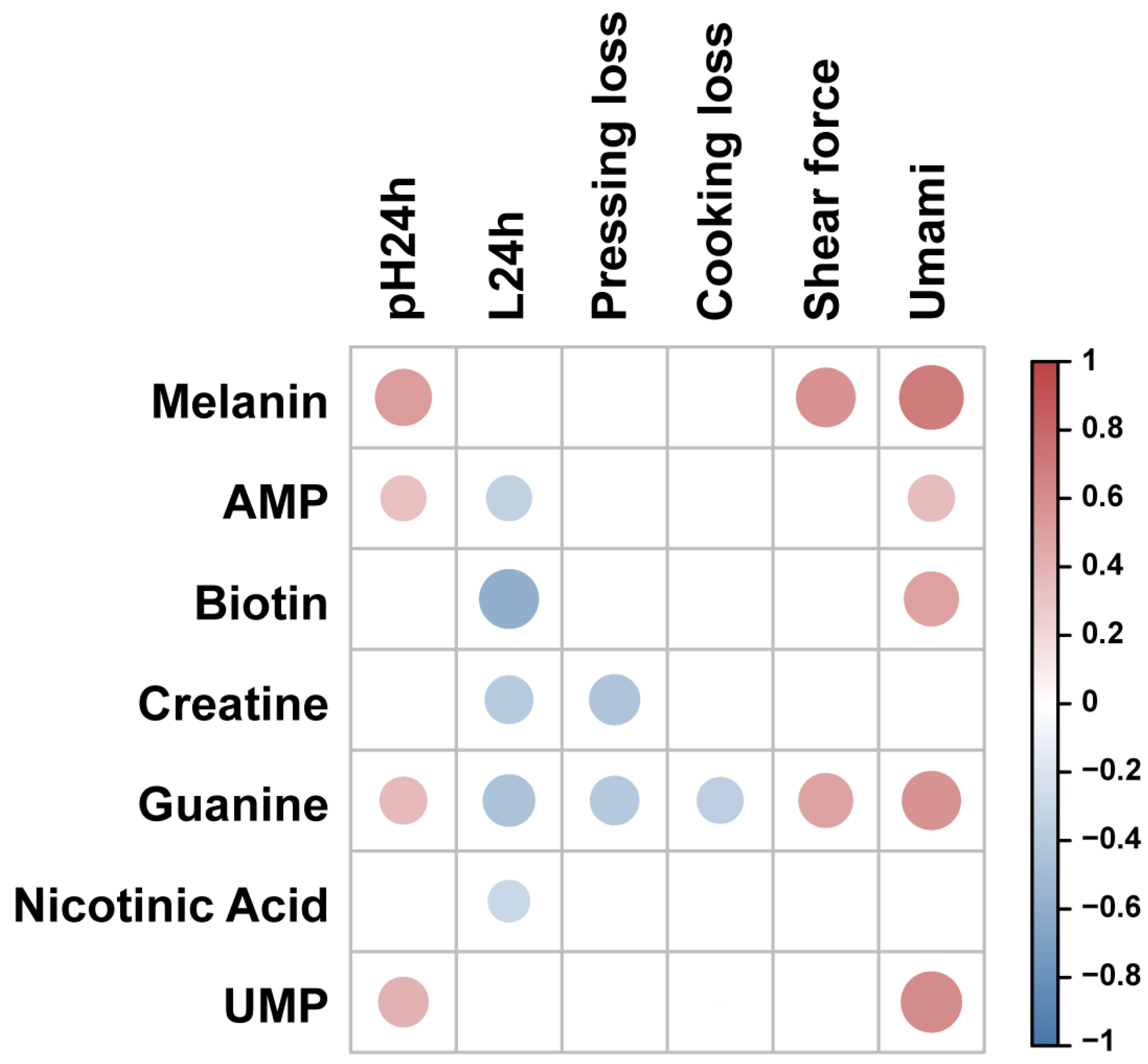
3.3. Effects of Characteristic Bioactive Metabolites on Meat Quality Traits
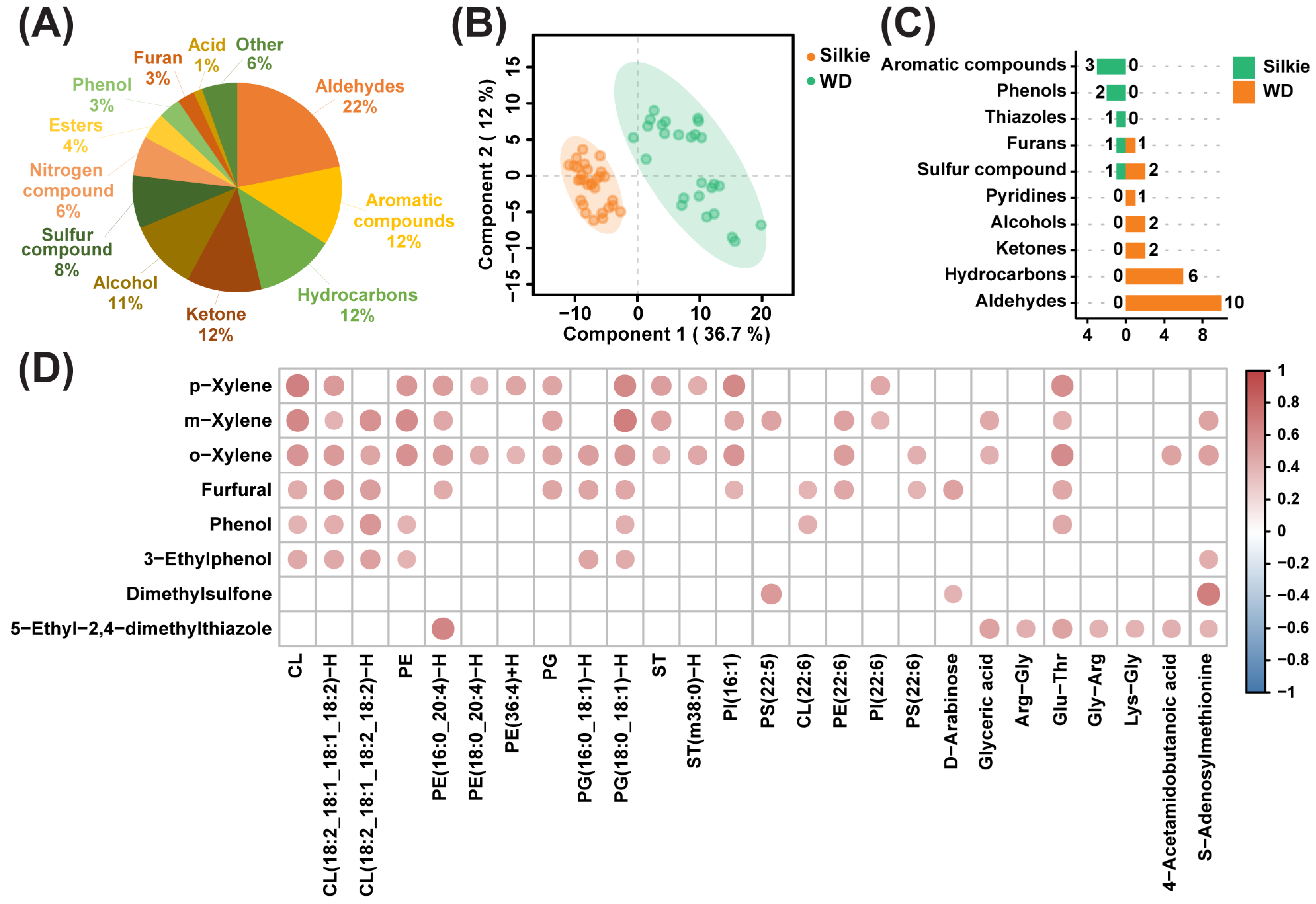
3.4. Effects of Characteristic Bioactive Metabolites, Lipids, and Fatty Acids on Characteristic Volatiles
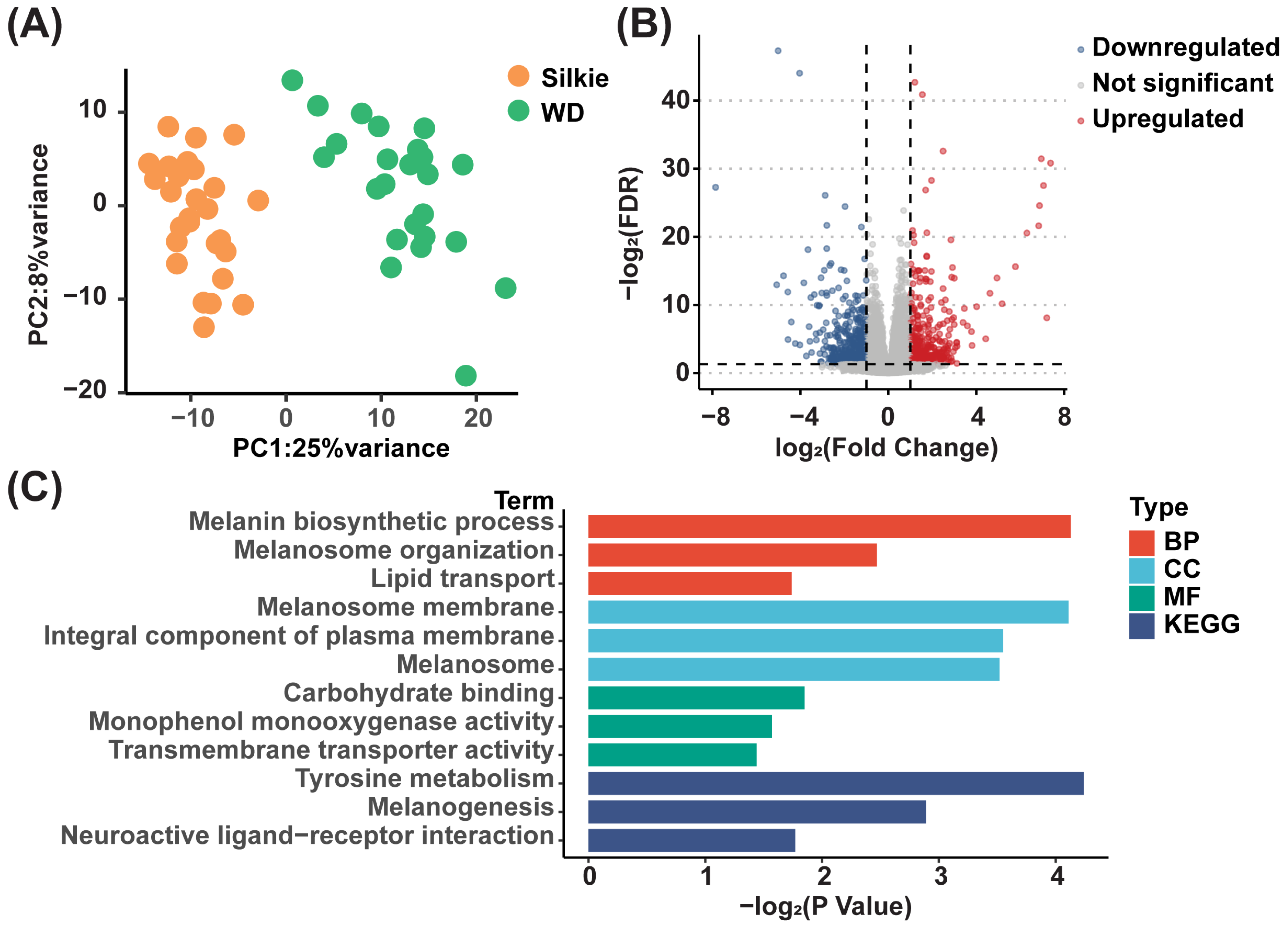
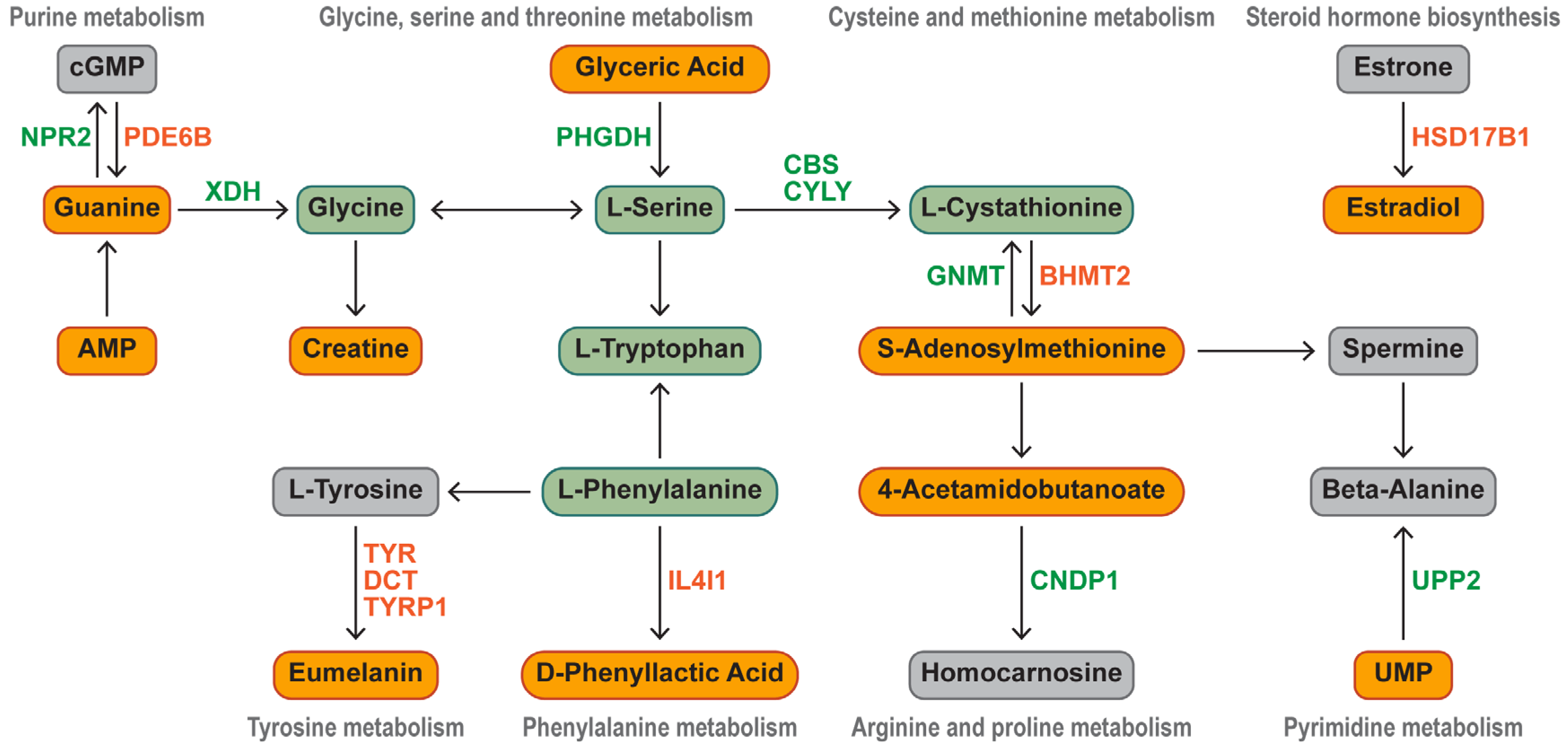
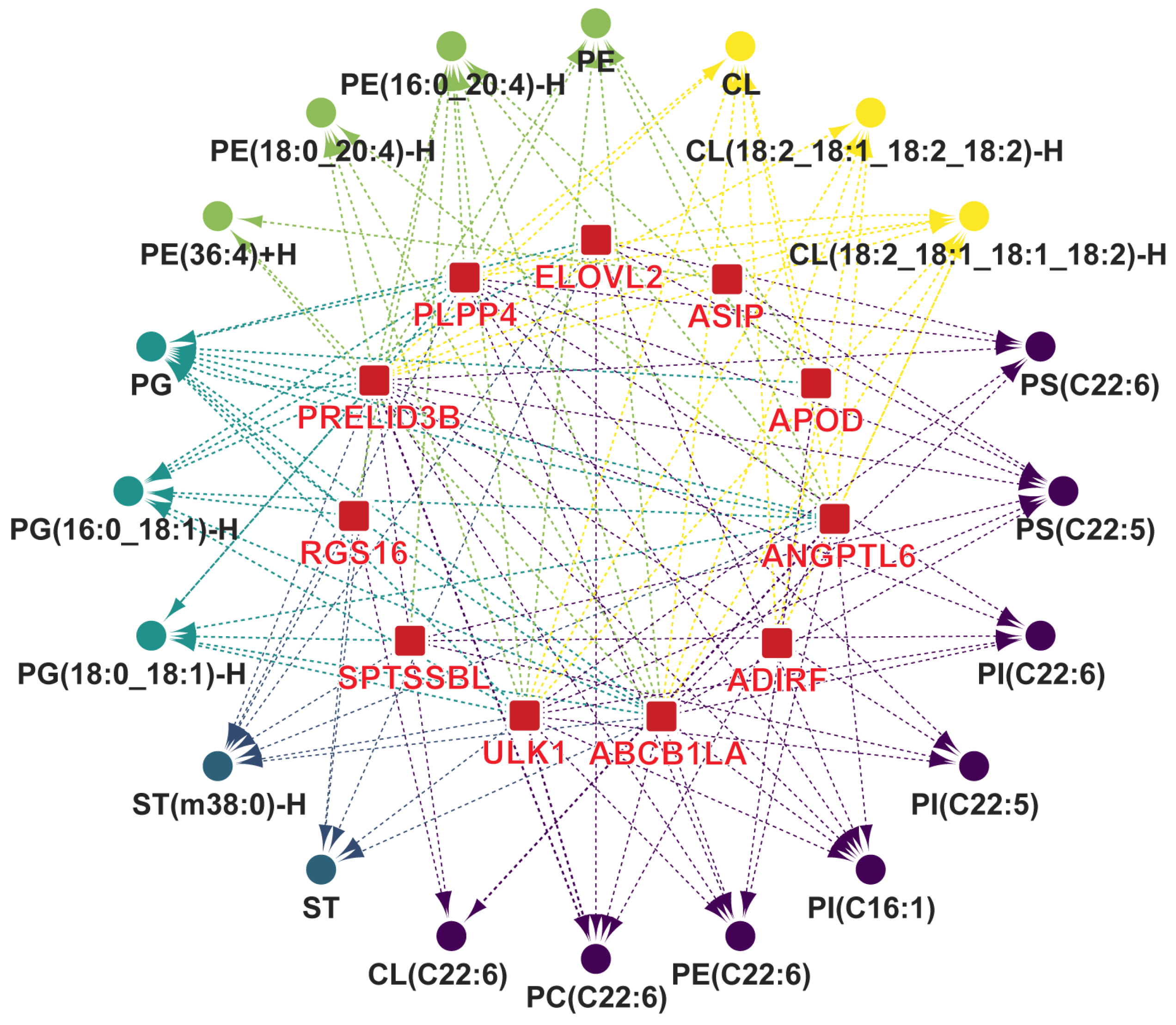
3.5. Identification of Key DEGs Involved in Synthesis of Characteristic Bioactive Metabolites, Lipids, and Fatty Acids
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, J.-H.; Tian, Y.-G.; Wang, Y.; Nie, S.-P.; Xie, M.-Y.; Zhu, S.; Wang, C.-Y.; Zhang, P. Characterization and in vitro antioxidation of papain hydrolysate from black-bone silky fowl (Gallus gallus domesticus Brisson) muscle and its fractions. Food Res. Int. 2011, 44, 133–138. [Google Scholar] [CrossRef]
- Qiu, Y.; Ying, J.; Yan, F.; Yu, H.; Zhao, Y.; Li, H.; Xia, S.; Chen, J.; Zhu, J. Novel antiosteoporotic peptides purified from protein hydrolysates of taihe black-boned silky fowl: By larval zebrafish model and molecular docking. Food Res. Int. 2023, 169, 112850. [Google Scholar] [CrossRef] [PubMed]
- Tu, Y.-G.; Sun, Y.-Z.; Tian, Y.-G.; Xie, M.-Y.; Chen, J. Physicochemical characterisation and antioxidant activity of melanin from the muscles of Taihe Black-bone silky fowl (Gallus gallus domesticus Brisson). Food Chem. 2009, 114, 1345–1350. [Google Scholar] [CrossRef]
- Yang, X.; Tang, C.; Zhao, Q.; Jia, Y.; Qin, Y.; Zhang, J. Melanin: A promising source of functional food ingredient. J. Funct. Foods 2023, 105, 105574. [Google Scholar] [CrossRef]
- Tian, Y.; Xie, M.; Wang, W.; Wu, H.; Fu, Z.; Lin, L. Determination of carnosine in Black-Bone Silky Fowl (Gallus gallus domesticus Brisson) and common chicken by HPLC. Eur. Food Res. Technol. 2007, 226, 311. [Google Scholar] [CrossRef]
- Wei, Q.; Cui, H.; Hu, Y.; Li, J.; Yue, S.; Tang, C.; Zhao, Q.; Yu, Y.; Li, H.; Qin, Y.; et al. Comparative characterization of Taihe silky chicken and Cobb chicken using LC/MS-based lipidomics and GC/MS-based volatilomics. LWT 2022, 163, 113554. [Google Scholar] [CrossRef]
- Mi, S.; Shang, K.; Jia, W.; Zhang, C.-H.; Li, X.; Fan, Y.-Q.; Wang, H. Characterization and discrimination of Taihe black-boned silky fowl (Gallus gallus domesticus Brisson) muscles using LC/MS-based lipidomics. Food Res. Int. 2018, 109, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Zhu, S.; Xie, M.; Wang, W.; Wu, H.; Gong, D. Composition of fatty acids in the muscle of black-bone silky chicken (Gallus gallus demesticus Brissen) and its bioactivity in mice. Food Chem. 2011, 126, 479–483. [Google Scholar] [CrossRef]
- Choo, Y.K.; Kwon, H.J.; Oh, S.T.; Um, J.S.; Kim, B.G.; Kang, C.W.; Lee, S.K.; An, B.K. Comparison of Growth Performance, Carcass Characteristics and Meat Quality of Korean Local Chickens and Silky Fowl. Asian-Australas. J. Anim. Sci. 2014, 27, 398–405. [Google Scholar] [CrossRef]
- Yan, E.; Guo, J.; Yin, J. Nutritional regulation of skeletal muscle energy metabolism, lipid accumulation and meat quality in pigs. Adv. Nutr. 2023, 14, 185–192. [Google Scholar] [CrossRef]
- Sun, A.; Wu, W.; Soladoye, O.P.; Aluko, R.E.; Bak, K.H.; Fu, Y.; Zhang, Y. Maillard reaction of food-derived peptides as a potential route to generate meat flavor compounds: A review. Food Res. Int. 2022, 151, 110823. [Google Scholar] [CrossRef]
- Li, J.; Zhang, D.; Yin, L.; Li, Z.; Yu, C.; Du, H.; Jiang, X.; Yang, C.; Liu, Y. Integration analysis of metabolome and transcriptome profiles revealed the age-dependent dynamic change in chicken meat. Food Res. Int. 2022, 156, 111171. [Google Scholar] [CrossRef]
- Tang, H.; Wang, X.; Xu, L.; Ran, X.; Li, X.; Chen, L.; Zhao, X.; Deng, H.; Liu, X. Establishment of local searching methods for orbitrap-based high throughput metabolomics analysis. Talanta 2016, 156–157, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Yang, M.; Hou, X.; Hou, R.; Wang, L.; Shi, L.; Zhao, F.; Liu, X.; Meng, Q.; Wang, L.; et al. Characterization and difference of lipids and metabolites from Jianhe White Xiang and Large White pork by high-performance liquid chromatography–tandem mass spectrometry. Food Res. Int. 2022, 162, 111946. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Wang, X.; Jiao, Y.; Liu, X. Assessment of potential false positives via orbitrap-based untargeted lipidomics from rat tissues. Talanta 2018, 178, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; Nakanishi, Y.; Valenzuela, R.K.; Brilliant, M.H.; Kolbe, L.; Wakamatsu, K. Usefulness of alkaline hydrogen peroxide oxidation to analyze eumelanin and pheomelanin in various tissue samples: Application to chemical analysis of human hair melanins. Pigm. Cell Melanoma Res. 2011, 24, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Liu, D.; Zhou, G.; Xu, X.; Qi, J.; Shi, P.; Xia, T. Meat quality and cooking attributes of thawed pork with different low field NMR T21. Meat Sci. 2012, 92, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Levy, C.; Robel, P.; Gautray, J.P.; De Brux, J.; Verma, U.; Descomps, B.; Baulieu, E.E. Estradiol and progesterone receptors in human endometrium: Normal and abnormal menstrual cycles and early pregnancy. Am. J. Obstet. Gynecol. 1980, 136, 646–651. [Google Scholar] [CrossRef] [PubMed]
- Gordon, J.L.; Rubinow, D.R.; Eisenlohr-Moul, T.A.; Xia, K.; Schmidt, P.J.; Girdler, S.S. Efficacy of Transdermal Estradiol and Micronized Progesterone in the Prevention of Depressive Symptoms in the Menopause Transition: A Randomized Clinical Trial. JAMA Psychiatry 2018, 75, 149–157. [Google Scholar] [CrossRef]
- Speroff, L. Efficacy and tolerability of a novel estradiol vaginal ring for relief of menopausal symptoms. Obstet. Gynecol. 2003, 102, 823–834. [Google Scholar] [CrossRef] [PubMed]
- Anstee, Q.M.; Day, C.P. S-adenosylmethionine (SAMe) therapy in liver disease: A review of current evidence and clinical utility. J. Hepatol. 2012, 57, 1097–1109. [Google Scholar] [CrossRef]
- Vahora, S.A.; Malek-Ahmadi, P. S-Adenosylmethionine in the treatment of depression. Neurosci. Biobehav. Rev. 1988, 12, 139–141. [Google Scholar] [CrossRef] [PubMed]
- Rutjes, A.W.S.; Nüesch, E.; Reichenbach, S.; Jüni, P. S-Adenosylmethionine for osteoarthritis of the knee or hip. Cochrane Database Syst. Rev. 2009, 2009, CD007321. [Google Scholar] [CrossRef]
- Nitz, K.; Lacy, M.; Bianchini, M.; Wichapong, K.; Kücükgöze, I.A.; Bonfiglio, C.A.; Migheli, R.; Wu, Y.; Burger, C.; Li, Y.; et al. The Amino Acid Homoarginine Inhibits Atherogenesis by Modulating T-Cell Function. Circ. Res. 2022, 131, 701–712. [Google Scholar] [CrossRef]
- Michos, E.D.; Sibley, C.T.; Baer, J.T.; Blaha, M.J.; Blumenthal, R.S. Niacin and Statin Combination Therapy for Atherosclerosis Regression and Prevention of Cardiovascular Disease Events: Reconciling the AIM-HIGH (Atherothrombosis Intervention in Metabolic Syndrome With Low HDL/High Triglycerides: Impact on Global Health Outcomes) Trial With Previous Surrogate Endpoint Trials. J. Am. Coll. Cardiol. 2012, 59, 2058–2064. [Google Scholar] [CrossRef]
- Nagasawa, T.; Nakamichi, H.; Hama, Y.; Higashiyama, S.; Igarashi, Y.; Mitsutake, S. Phytosphingosine is a novel activator of GPR120. J. Biochem. 2018, 164, 27–32. [Google Scholar] [CrossRef]
- Snel, M.; Sleddering, M.A.; Pijl, H.; Nieuwenhuizen, W.F.; Frölich, M.; Havekes, L.M.; Romijn, J.A.; Jazet, I.M. The effect of dietary phytosphingosine on cholesterol levels and insulin sensitivity in subjects with the metabolic syndrome. Eur. J. Clin. Nutr. 2010, 64, 419–423. [Google Scholar] [CrossRef]
- Zhao, Y.; Xu, J.; Zhao, C.; Bao, L.; Wu, K.; Feng, L.; Sun, H.; Shang, S.; Hu, X.; Sun, Q.; et al. Phytosphingosine alleviates Staphylococcus aureus-induced mastitis by inhibiting inflammatory responses and improving the blood-milk barrier in mice. Microb. Pathog. 2023, 182, 106225. [Google Scholar] [CrossRef]
- Hirvonen, O.P.; Kyröläinen, H.; Lehti, M.; Kainulainen, H. Randomized Trial: D-Glyceric Acid Activates Mitochondrial Metabolism in 50–60-Year-Old Healthy Humans. Front. Aging 2021, 2, 752636. [Google Scholar] [CrossRef]
- Hirvonen, O.P.; Lehti, M.; Kyröläinen, H.; Kainulainen, H. Heme Oxygenase-1 and Blood Bilirubin Are Gradually Activated by Oral D-Glyceric Acid. Antioxidants 2022, 11, 2319. [Google Scholar] [CrossRef]
- Zhou, Q.; Gu, R.; Xue, B.; Li, P.; Gu, Q. Phenyl lactic acid alleviates Samonella typhimurium-induced colitis via regulating microbiota composition, SCFA production and inflammatory responses. Food Funct. 2021, 12, 5591–5606. [Google Scholar] [CrossRef]
- Holguin, S.; Martinez, J.; Chow, C.; Wurtman, R. Dietary uridine enhances the improvement in learning and memory produced by administering DHA to gerbils. FASEB J. 2008, 22, 3938–3946. [Google Scholar] [CrossRef] [PubMed]
- Ostojic, S.M. Creatine as a food supplement for the general population. J. Funct. Foods 2021, 83, 104568. [Google Scholar] [CrossRef]
- Tasca, C.I.; Lanznaster, D.; Oliveira, K.A.; Fernández-Dueñas, V.; Ciruela, F. Neuromodulatory Effects of Guanine-Based Purines in Health and Disease. Front. Cell Neurosci. 2018, 12, 376. [Google Scholar] [CrossRef]
- Darwish, A.G.; Das, P.R.; Ismail, A.; Gajjar, P.; Balasubramani, S.P.; Sheikh, M.B.; Tsolova, V.; Sherif, S.M.; El-Sharkawy, I. Untargeted Metabolomics and Antioxidant Capacities of Muscadine Grape Genotypes during Berry Development. Antioxidants 2021, 10, 914. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Rooney, M.R.; Burmeister, L.A.; Basta, N.E.; Lutsey, P.L. Trends in Daily Use of Biotin Supplements Among US Adults, 1999-2016. JAMA 2020, 324, 605–607. [Google Scholar] [CrossRef] [PubMed]
- Lipner, S.R. Rethinking biotin therapy for hair, nail, and skin disorders. J. Am. Acad. Dermatol. 2018, 78, 1236–1238. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kim, B.-K.; Park, S.-K. Supplementation with phosphatidylethanolamine confers anti-oxidant and anti-aging effects via hormesis and reduced insulin/IGF-1-like signaling in C. elegans. Mech. Ageing Dev. 2021, 197, 111498. [Google Scholar] [CrossRef] [PubMed]
- Falabella, M.; Vernon, H.J.; Hanna, M.G.; Claypool, S.M.; Pitceathly, R.D.S. Cardiolipin, Mitochondria, and Neurological Disease. Trends Endocrinol. Metab. 2021, 32, 224–237. [Google Scholar] [CrossRef]
- Qiu, S.; Palavicini, J.P.; Wang, J.; Gonzalez, N.S.; He, S.; Dustin, E.; Zou, C.; Ding, L.; Bhattacharjee, A.; Van Skike, C.E.; et al. Adult-onset CNS myelin sulfatide deficiency is sufficient to cause Alzheimer’s disease-like neuroinflammation and cognitive impairment. Mol. Neurodegener. 2021, 16, 64. [Google Scholar] [CrossRef]
- Frigolet, M.E.; Gutiérrez-Aguilar, R. The Role of the Novel Lipokine Palmitoleic Acid in Health and Disease. Adv. Nutr. 2017, 8, 173S–181S. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Ambigaipalan, P. Omega-3 Polyunsaturated Fatty Acids and Their Health Benefits. Annu. Rev. Food. Sci. Technol. 2018, 9, 345–381. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, G.; Zhang, Y.; Wang, X.; Jin, Q.; Zhang, H. Advances in exogenous docosahexaenoic acid-containing phospholipids: Sources, positional isomerism, biological activities, and advantages. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1420–1448. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.-Y.; Hsu, F.-W.; Chang, H.-S.; Lin, L.-C.; Sakata, R. Effect of different acids on the extraction of pepsin-solubilised collagen containing melanin from silky fowl feet. Food Chem. 2009, 113, 563–567. [Google Scholar] [CrossRef]
- Starkey, C.P.; Geesink, G.H.; Oddy, V.H.; Hopkins, D.L. Explaining the variation in lamb longissimus shear force across and within ageing periods using protein degradation, sarcomere length and collagen characteristics. Meat Sci. 2015, 105, 32–37. [Google Scholar] [CrossRef]
- Wang, D.; Deng, S.; Zhang, M.; Geng, Z.; Sun, C.; Bian, H.; Xu, W.; Zhu, Y.; Liu, F.; Wu, H. The effect of adenosine 5′-monophosphate (AMP) on tenderness, microstructure and chemical-physical index of duck breast meat. J. Sci. Food Agric. 2016, 96, 1467–1473. [Google Scholar] [CrossRef]
- Martelli, G.; Sardi, L.; Parisini, P.; Badiani, A.; Parazza, P.; Mordenti, A. The effects of a dietary supplement of biotin on Italian heavy pigs’ (160 kg) growth, slaughtering parameters, meat quality and the sensory properties of cured hams. Livest. Prod. Sci. 2005, 93, 117–124. [Google Scholar] [CrossRef]
- Young, J.F.; Bertram, H.C.; Rosenvold, K.; Lindahl, G.; Oksbjerg, N. Dietary creatine monohydrate affects quality attributes of Duroc but not Landrace pork. Meat Sci. 2005, 70, 717–725. [Google Scholar] [CrossRef]
- Li, J.; Zhang, L.; Fu, Y.; Li, Y.; Jiang, Y.; Zhou, G.; Gao, F. Creatine Monohydrate and Guanidinoacetic Acid Supplementation Affects the Growth Performance, Meat Quality, and Creatine Metabolism of Finishing Pigs. J. Agric. Food Chem. 2018, 66, 9952–9959. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Hao, J.; Yin, H.; Duan, C.; Wang, B.; Li, W. Effects of dietary nicotinic acid supplementation on meat quality, carcass characteristics, lipid metabolism, and tibia parameters of Wulong geese. Poult. Sci. 2021, 100, 101430. [Google Scholar] [CrossRef] [PubMed]
- Fuke, S.; Shimizu, T. Sensory and preference aspects of umami. Trends Food Sci. Technol. 1993, 4, 246–251. [Google Scholar] [CrossRef]
- Huang, C.; Zheng, M.; Huang, Y.; Liu, X.; Zhong, L.; Ji, J.; Zhou, L.; Zeng, Q.; Ma, J.; Huang, L. The effect of purine content on sensory quality of pork. Meat Sci. 2021, 172, 108346. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Y.; Li, H.; Bai, T.; Qian, Q.; Peng, H.; Mu, Y.; Wang, L.; Liu, B.; Chen, J.; et al. Effect of 4 °C and ice temperature on umami-enhancing nucleotides of conditioned pork. Food Chem. 2023, 401, 134146. [Google Scholar] [CrossRef]
- Gong, M.; Zhou, Z.; Liu, S.; Zhu, S.; Li, G.; Zhong, F.; Mao, J. Formation pathways and precursors of furfural during Zhenjiang aromatic vinegar production. Food Chem. 2021, 354, 129503. [Google Scholar] [CrossRef]
- Han, Y.; You, M.; Wang, S.; Yuan, Q.; Gao, P.; Zhong, W.; Yin, J.; Hu, C.; He, D.; Wang, X. Enzymatic methods for the preparation of fragrant rapeseed oil: Effect of reducing sugars on flavor production using the Maillard reaction. LWT 2023, 189, 115497. [Google Scholar] [CrossRef]
- Huang, X.; You, Y.; Liu, Q.; Dong, H.; Bai, W.; Lan, B.; Wu, J. Effect of gamma irradiation treatment on microstructure, water mobility, flavor, sensory and quality properties of smoked chicken breast. Food Chem. 2023, 421, 136174. [Google Scholar] [CrossRef]
- Sohail, A.; Al-Dalali, S.; Wang, J.; Xie, J.; Shakoor, A.; Asimi, S.; Shah, H.; Patil, P. Aroma compounds identified in cooked meat: A review. Food Res. Int. 2022, 157, 111385. [Google Scholar] [CrossRef]
- van Helvoort, A.; Smith, A.J.; Sprong, H.; Fritzsche, I.; Schinkel, A.H.; Borst, P.; van Meer, G. MDR1 P-glycoprotein is a lipid translocase of broad specificity, while MDR3 P-glycoprotein specifically translocates phosphatidylcholine. Cell 1996, 87, 507–517. [Google Scholar] [CrossRef] [PubMed]
- Qu, J.; Fourman, S.; Fitzgerald, M.; Liu, M.; Nair, S.; Oses-Prieto, J.; Burlingame, A.; Morris, J.H.; Davidson, W.S.; Tso, P.; et al. Low-density lipoprotein receptor-related protein 1 (LRP1) is a novel receptor for apolipoprotein A4 (APOA4) in adipose tissue. Sci. Rep. 2021, 11, 13289. [Google Scholar] [CrossRef] [PubMed]
- Rassart, E.; Desmarais, F.; Najyb, O.; Bergeron, K.-F.; Mounier, C. Apolipoprotein D. Gene 2020, 756, 144874. [Google Scholar] [CrossRef] [PubMed]
- Miliara, X.; Tatsuta, T.; Berry, J.-L.; Rouse, S.L.; Solak, K.; Chorev, D.S.; Wu, D.; Robinson, C.V.; Matthews, S.; Langer, T. Structural determinants of lipid specificity within Ups/PRELI lipid transfer proteins. Nat. Commun. 2019, 10, 1130. [Google Scholar] [CrossRef] [PubMed]
- Xie, T.; Liu, Y.; Lu, H.; Iqbal, A.; Ruan, M.; Jiang, P.; Yu, H.; Meng, J.; Zhao, Z. The Knockout of the ASIP Gene Altered the Lipid Composition in Bovine Mammary Epithelial Cells via the Expression of Genes in the Lipid Metabolism Pathway. Animals 2022, 12, 1389. [Google Scholar] [CrossRef] [PubMed]
- Pauter, A.M.; Trattner, S.; Gonzalez-Bengtsson, A.; Talamonti, E.; Asadi, A.; Dethlefsen, O.; Jacobsson, A. Both maternal and offspring Elovl2 genotypes determine systemic DHA levels in perinatal mice. J. Lipid Res. 2017, 58, 111–123. [Google Scholar] [CrossRef]
- Hoch, D.G.; Abegg, D.; Hannich, J.T.; Pechalrieu, D.; Shuster, A.; Dwyer, B.G.; Wang, C.; Zhang, X.; You, Q.; Riezman, H.; et al. Combined Omics Approach Identifies Gambogic Acid and Related Xanthones as Covalent Inhibitors of the Serine Palmitoyltransferase Complex. Cell Chem. Biol. 2020, 27, 586–597. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.A.; Kim, H.J.; Ahn, H.Y.; Park, K.U.; Yi, K.H.; Park, D.J.; Jang, H.C.; Park, Y.J. Influence of thyroid dysfunction on serum levels of angiopoietin-like protein 6. Metabolism 2015, 64, 1279–1283. [Google Scholar] [CrossRef]
- Zheng, B.; Chen, D.; Farquhar, M.G. MIR16, a putative membrane glycerophosphodiester phosphodiesterase, interacts with RGS16. Proc. Natl. Acad. Sci. USA 2000, 97, 3999–4004. [Google Scholar] [CrossRef]
- Ro, S.-H.; Jung, C.H.; Hahn, W.S.; Xu, X.; Kim, Y.-M.; Yun, Y.S.; Park, J.-M.; Kim, K.H.; Seo, M.; Ha, T.-Y.; et al. Distinct functions of Ulk1 and Ulk2 in the regulation of lipid metabolism in adipocytes. Autophagy 2013, 9, 2103–2114. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Silkie Chickens | Wuding Chickens | p Value | |
|---|---|---|---|
| pH24h | 5.75 ± 0.28 a | 5.45 ± 0.29 b | 0.01 |
| L*24h | 48.83 ± 2.16 b | 59.40 ± 3.30 a | <0.001 |
| a*24h | 3.00 ± 1.21 | 2.57 ± 1.53 | |
| b*24h | 5.24 ± 1.96 b | 8.19 ± 2.92 a | <0.001 |
| Pressing loss (%) | 7.30 ± 1.51 b | 8.31 ± 1.88 a | 0.02 |
| Cooking loss (%) | 13.95 ± 1.61 b | 15.19 ± 2.22 a | 0.02 |
| Shear force (kg) | 3.62 ± 0.93 a | 3.15 ± 0.58 b | <0.001 |
| Melanin content (ng/g) | 439.72 ± 337.85 a | 0.00 ± 0.00 b | <0.001 |
| Silkie Chickens | Wuding Chickens | p Value | |
|---|---|---|---|
| Umami | 8.52 ± 0.51 a | 8.02 ± 0.49 b | <0.01 |
| Richness | 5.39 ± 0.43 | 5.32 ± 0.48 | |
| Bitterness | 1.90 ± 0.58 | 1.79 ± 0.58 | |
| Aftertaste-B | −0.26 ± 0.16 | −0.33 ± 0.23 | |
| Astringency | −3.97 ± 0.36 b | −3.68 ± 0.32 a | 0.01 |
| Aftertaste-A | 0.004 ± 0.03 | −0.01 ± −0.01 | |
| Saltiness | −12.10 ± 0.50 | −12.22 ± 0.52 | |
| Sourness | −21.73 ± 1.78 b | −19.90 ± 1.66 a | <0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Tang, C.; Ma, B.; Zhao, Q.; Jia, Y.; Meng, Q.; Qin, Y.; Zhang, J. Identification of Characteristic Bioactive Compounds in Silkie Chickens, Their Effects on Meat Quality, and Their Gene Regulatory Network. Foods 2024, 13, 969. https://doi.org/10.3390/foods13060969
Yang X, Tang C, Ma B, Zhao Q, Jia Y, Meng Q, Qin Y, Zhang J. Identification of Characteristic Bioactive Compounds in Silkie Chickens, Their Effects on Meat Quality, and Their Gene Regulatory Network. Foods. 2024; 13(6):969. https://doi.org/10.3390/foods13060969
Chicago/Turabian StyleYang, Xinting, Chaohua Tang, Bowen Ma, Qingyu Zhao, Yaxiong Jia, Qingshi Meng, Yuchang Qin, and Junmin Zhang. 2024. "Identification of Characteristic Bioactive Compounds in Silkie Chickens, Their Effects on Meat Quality, and Their Gene Regulatory Network" Foods 13, no. 6: 969. https://doi.org/10.3390/foods13060969
APA StyleYang, X., Tang, C., Ma, B., Zhao, Q., Jia, Y., Meng, Q., Qin, Y., & Zhang, J. (2024). Identification of Characteristic Bioactive Compounds in Silkie Chickens, Their Effects on Meat Quality, and Their Gene Regulatory Network. Foods, 13(6), 969. https://doi.org/10.3390/foods13060969