
Clovamide and Its Derivatives—Bioactive Components of Theobroma cacao and Other Plants in the Context of Human Health

Abstract
:1. Introduction
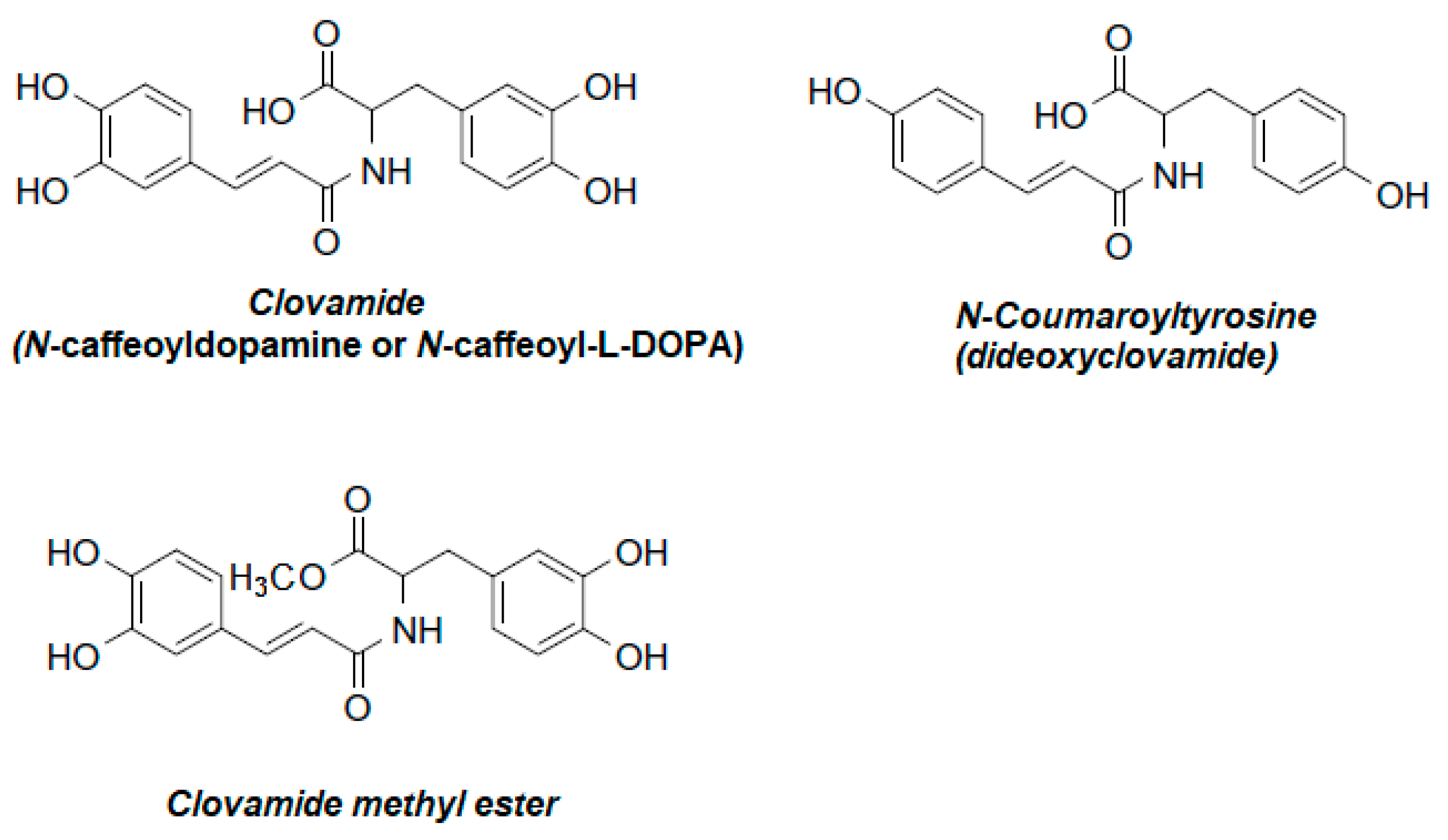
2. Plant Sources of Clovamides
3. Bioavailability and Metabolism of Clovamide-Type Compounds
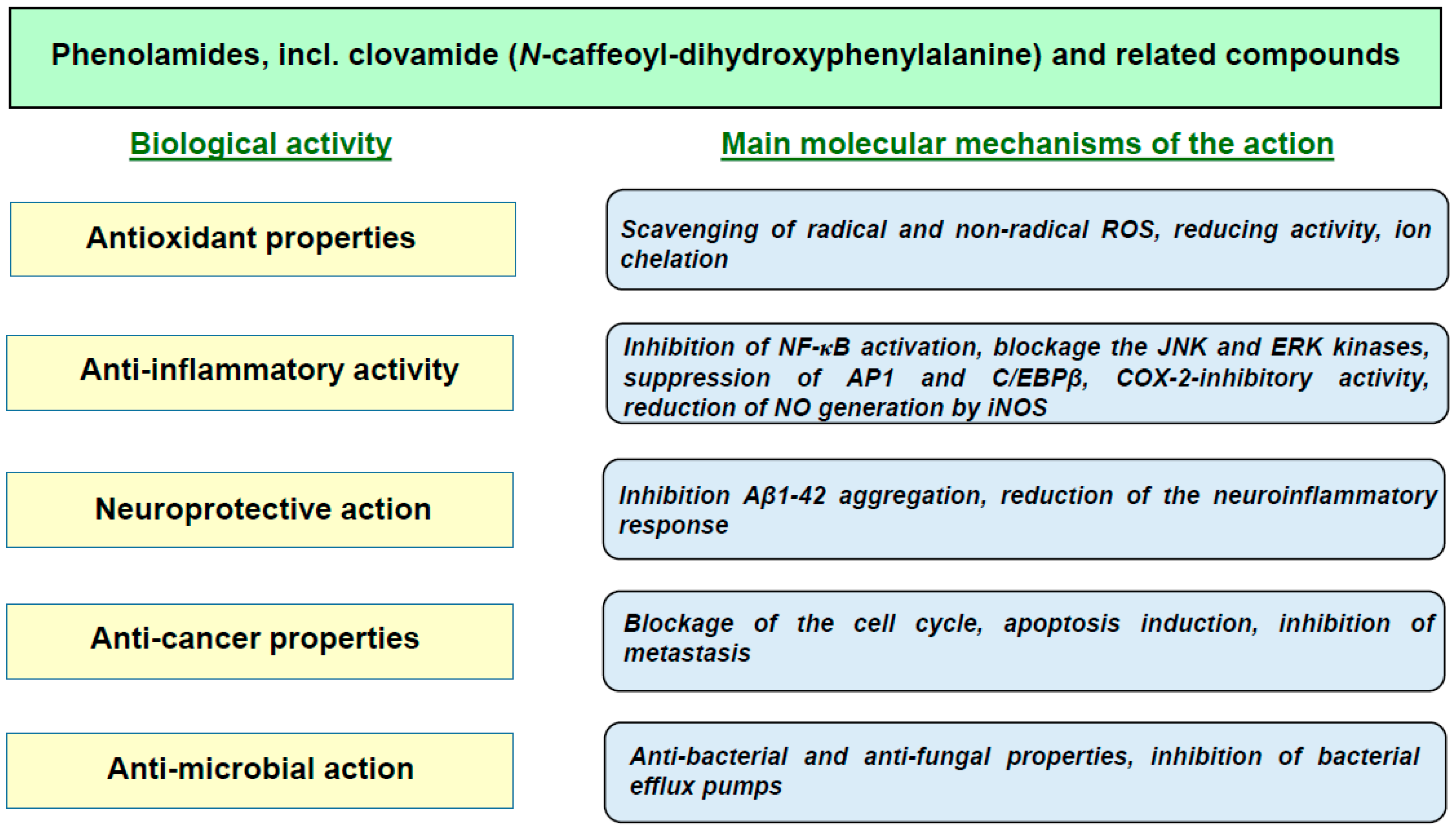
4. Biological Activity of Clovamides
4.1. Antioxidant Action

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Model | EC50 or IC50 Values Established for Clovamide | EC50 or IC50 Values Established for Reference Compounds | References |
|---|---|---|---|
| DPPH• scavenging | Clovamide: 2.65 μg/mL | Caffeic acid: 2.93 μg/mL Epicatechin: 3.11 μg/mL Gallic acid: 1.03 μg/mL Rosmarinic acid: 2.49 μg/mL Myricetin: 1.95 μg/mL Quercetin: 1.99 μg/mL Kaempferol: 4.26 μg/mL BHA: 8.18 μg/mL Trolox: 3.32 μg/mL Octyl gallate: 1.65 μg/mL | [56] |
| Clovamide: 4.9 μg/mL | Caffeic acid: 4.2 μg/mL Chlorogenic acid: 8.3 μg/mL Trolox: 5.7 μg/mL | [30] | |
| Clovamide: 0.05 mol/mol | Rosmarinic acid: 0.58 mol/mol α-Tocopherol: 0.025 mol/mol Ascorbic acid: 0.025 mol/mol L-dopamine: 0.095 mol/mol | [59] | |
| ONOO− scavenging | Clovamide: 19.3 μg/mL | Caffeic acid: 15.0 μg/mL Chlorogenic acid: 27.4 μg/mL Trolox: <5 μg/mL | [30] |
| Superoxide anion scavenging | Clovamide: 60 nmol/L | Rosmarinic acid: 95 nmol/L α-Tocopherol: >10 000 nmol/L Ascorbic acid: 700 nmol/L L-dopamine: 200 nmol/L | [59] |
| β-carotene bleaching | Clovamide: 0.02 mmol/L | α-Tocopherol: 0.08 mmol/L Ascorbic acid: >0.09 L-dopamine: >1.1 mmol/L | [59] |
4.2. Anti-Inflammatory Effects
4.3. Neuroprotective Effects
4.4. Anti-Platelet Action
4.5. Anticancer Properties
4.6. Antiviral, Antibacterial and Anti-Trypanosomal Activities
4.7. Estrogenic Activity
5. Concluding Remarks
Funding
Data Availability Statement
Conflicts of Interest
References
- Lippi, D. Sin and Pleasure: The History of Chocolate in Medicine. J. Agric. Food Chem. 2015, 63, 9936–9941. [Google Scholar] [CrossRef] [PubMed]
- Montagna, M.T.; Diella, G.; Triggiano, F.; Caponio, G.R.; De Giglio, O.; Caggiano, G.R.; Di Ciaula, A.; Portincasa, P. Chocolate, “Food of the Gods”: History, Science, and Human Health. Int. J. Environ. Res. Public Health 2019, 16, 4960. [Google Scholar] [CrossRef] [PubMed]
- Magrone, T.; Russo, M.A.; Jirillo, E. Cocoa and Dark Chocolate Polyphenols: From Biology to Clinical Applications. Front. Immunol. 2017, 8, 677. [Google Scholar] [CrossRef] [PubMed]
- Tan, T.Y.C.; Lim, X.Y.; Yeo, J.H.H.; Lee, S.W.H.; Lai, N.M. The Health Effects of Chocolate and Cocoa: A Systematic Review. Nutrients 2021, 13, 2909. [Google Scholar] [CrossRef] [PubMed]
- Roumani, M.; Duval, R.E.; Ropars, A.; Risler, A.; Robin, C.; Larbat, R. Phenolamides: Plant Specialized Metabolites with a Wide Range of Promising Pharmacological and Health-Promoting Interests. Biomed. Pharmacother. 2020, 131, 110762. [Google Scholar] [CrossRef] [PubMed]
- Knollenberg, B.J.; Li, G.-X.; Lambert, J.D.; Maximova, S.N.; Guiltinan, M.J. Clovamide, a Hydroxycinnamic Acid Amide, Is a Resistance Factor Against Phytophthora spp. in Theobroma cacao. Front. Plant Sci. 2020, 11, 617520. [Google Scholar] [CrossRef] [PubMed]
- Park, J.B. Quantitation of Clovamide-Type Phenylpropenoic Acid Amides in Cells and Plasma Using High-Performance Liquid Chromatography with a Coulometric Clectrochemical Detector. J. Agric. Food Chem. 2005, 53, 8135–8140. [Google Scholar] [CrossRef]
- Xie, Y.; Huang, B.; Yu, K.; Shi, F.; Liu, T.; Xu, W. Caffeic Acid Derivatives: A New Type of Influenza Neuraminidase Inhibitors. Bioorg. Med. Chem. Lett. 2013, 23, 3556–3560. [Google Scholar] [CrossRef]
- Bouchez, P.; Teixeira Benites, V.; Baidoo, E.E.K.; Mortimer, J.C.; Sullivan, M.L.; Scheller, H.V.; Eudes, A. Production of Clovamide and Its Analogues in Saccharomyces cerevisiae and Lactococcus lactis. Lett. Appl. Microbiol. 2019, 69, 181–189. [Google Scholar]
- Yoshihara, T.; Yoshikawa, H.; Sakamura, S.; Sakuma, T. Clovamides: L-DOPA Conjugated with Trans- and Cis-Caffeic acids in Red Clover. Agric. Biol. Chem. 1974, 38, 1107–1109. [Google Scholar] [CrossRef]
- Sanbongi, C.; Osakabe, N.; Natsume, M.; Takizawa, T.; Gomi, S.; Osawa, T. Antioxidative Polyphenols Isolated from Theobroma cacao. J. Agric. Food Chem. 1998, 46, 454–457. [Google Scholar] [CrossRef] [PubMed]
- de Moraes Barros, H.R.; García-Villalba, R.; Tomás-Barberán, F.A.; Genovese, M.I. Evaluation of the Distribution and Metabolism of Polyphenols Derived from Cupuassu (Theobroma grandiflorum) in Mice Gastrointestinal Tract by UPLC-ESI-QTOF. J. Funct. Foods 2016, 22, 477–489. [Google Scholar] [CrossRef]
- Yoshihara, T.; Yoshikawa, H.; Kunimatsu, S.; Sakamura, S.; Sakuma, T. New Amino Acid Derivatives Conjugated with Caffeic Acid and DOPA from Red Clover (Trifolium pratense). Agric. Biol. Chem. 1977, 41, 1679–1684. [Google Scholar]
- Szajwaj, B.; Moldoch, J.; Masullo, M.; Piacente, S.; Oleszek, W.; Stochmal, A. Amides and Esters of Phenylpropenoic Acids from the Aerial Parts of Trifolium pallidum. Nat. Prod. Commun. 2011, 6, 1293–1296. [Google Scholar] [CrossRef] [PubMed]
- Masike, K.; Khoza, B.S.; Steenkamp, P.A.; Smit, E.; Dubery, I.A.; Madala, N.E. A Metabolomics-Guided Exploration of the Phytochemical Constituents of Vernonia fastigiata with the Aid of Pressurized Hot Water Extraction and Liquid Chromatography-Mass Spectrometry. Molecules 2017, 22, 1200. [Google Scholar] [CrossRef] [PubMed]
- Van Heerden, F.R.; Brandt, E.V.; Roux, D.G. Isolation and synthesis of trans- and cis-(−)-clovamides and their deoxy analogues from the bark of Dalbergia melanoxylon. Phytochemistry 1980, 19, 2125–2129. [Google Scholar] [CrossRef]
- Nascimento, L.E.S.; Arriola, N.D.A.; da Silva, L.A.L.; Faqueti, L.G.; Sandjo, L.P.; de Araújo, C.E.S.; Biavatti, M.W.; Barcelos-Oliveira, J.L.; de Mello Castanho Amboni, R.D. Phytochemical profile of different anatomical parts of jambu (Acmella oleracea (L.) R.K. Jansen): A comparison between hydroponic and conventional cultivation using PCA and cluster analysis. Food Chem. 2020, 332, 127393. [Google Scholar] [CrossRef] [PubMed]
- Kasper, J.; Melzig, M.F.; Jenett-Siems, K. New Phenolic Compounds of Acmella ciliata. Planta Medica 2010, 76, 633–635. [Google Scholar] [CrossRef] [PubMed]
- El-Sharawy, R.T.; Elkhateeb, A.; Marzouk, M.M.; Abd El-Latif, R.R.; Abdelrazig, S.E.; El-Ansari, M.A. Antiviral and An-tiparasitic Activities of Clovamide: The Major Constituent of Dichrostachys cinerea (L.) Wight et Arn. J. Appl. Pharm. Sci. 2017, 7, 219–223. [Google Scholar]
- Abouelela, M.E.; Orabi, M.A.A.; Abdelhamid, R.A.; Abdelkader, M.S.; Madkor, H.R.; Darwish, F.M.M.; Hatano, T.; Elsadek, B.E.M. Ethyl Acetate Extract of Ceiba pentandra (L.) Gaertn. Reduces Methotrexate-Induced Renal Damage in Rats via Antioxidant, Anti-inflammatory, and Antiapoptotic Actions. J. Tradit. Complement. Med. 2020, 10, 478–486. [Google Scholar] [CrossRef]
- Burlec, A.F.; Pecio, Ł.; Mircea, C.; Cioancă, O.; Corciovă, A.; Nicolescu, A.; Oleszek, W.; Hăncianu, M. Chemical Profile and Antioxidant Activity of Zinnia elegans Jacq. Fractions. Molecules 2019, 24, 2934. [Google Scholar] [CrossRef]
- Francišković, M.; Gonzalez-Pérez, R.; Orčić, D.; Sánchez de Medina, F.; Martínez-Augustin, O.; Svirčev, E.; Simin, N.; Mimica-Dukić, N. Chemical Composition and Immuno-Modulatory Effects of Urtica dioica L. (Stinging Nettle) Extracts. Phytother. Res. 2017, 31, 1183–1191. [Google Scholar] [CrossRef] [PubMed]
- Ye, N.; Belli, S.; Caruso, F.; Roy, G.; Rossi, M. Antioxidant Studies by Hydrodynamic Voltammetry and DFT, Quantitative Analyses by HPLC-DAD of Clovamide, a Natural Phenolic Compound Found in Theobroma cacao L. beans. Food Chem. 2021, 341, 128260. [Google Scholar] [CrossRef]
- Arlorio, M.; Locatelli, M.; Travaglia, F.; Coïsson, J.-D.; Del Grosso, E.; Minassi, A.; Appendino, G.; Martelli, A. Roasting Impact on the Contents of Clovamide (N-caffeoyl-L-DOPA) and the Antioxidant Activity of Cocoa Beans (Theobroma cacao L.). Food Chem. 2008, 106, 967–975. [Google Scholar] [CrossRef]
- Lechtenberg, M.; Henschel, K.; Liefländer-Wulf, U.; Quandt, B.; Hensel, A. Fast Determination of N-Phenylpropenoyl-l-Amino acids (NPA) in Cocoa Samples from Different Origins by Ultra-Performance Liquid Chro-matography and Capillary Electrophoresis. Food Chem. 2012, 135, 1676–1684. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.N.; Kellard, B.; Ah-Sing, E. Caffeoyltyrosine from Green Robusta Coffee Beans. Phytochemistry 1989, 28, 1989–1990. [Google Scholar] [CrossRef]
- Kolodziejczyk-Czepas, J. Trifolium species—The Latest Findings on Chemical Profile, Ethnomedicinal Use and Pharmaco-logical Properties. J. Pharm. Pharmacol. 2016, 68, 845–861. [Google Scholar] [CrossRef] [PubMed]
- Kanadys, W.; Barańska, A.; Błaszczuk, A.; Polz-Dacewicz, M.; Drop, B.; Kanecki, K.; Malm, M. Evaluation of Clinical Meaningfulness of Red Clover (Trifolium pratense L.) Extract to Relieve Hot Flushes and Menopausal Symptoms in Peri- and Post-Menopausal Women: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Nutrients 2021, 13, 1258. [Google Scholar] [CrossRef]
- Oleszek, W.; Stochmal, A.; Janda, B. Concentration of Isoflavones and Other Phenolics in the Aerial Parts of Trifolium Species. J. Agric. Food Chem. 2007, 55, 8095–8100. [Google Scholar] [CrossRef]
- Kolodziejczyk-Czepas, J.; Krzyżanowska-Kowalczyk, J.; Sieradzka, M.; Nowak, P.; Stochmal, A. Clovamide and Clovamide-Rich Extracts of Three Trifolium Species as Antioxidants and Moderate Antiplatelet Agents In Vitro. Phytochemistry 2017, 143, 54–63. [Google Scholar] [CrossRef]
- Sullivan, M.L.; Zeller, W.E. Efficacy of Various Naturally Occurring Caffeic Acid Derivatives in Preventing Post-Harvest Protein Losses in Forages. J. Sci. Food Agric. 2013, 93, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Tava, A.; Pecio, Ł.; Stochmal, A.; Pecetti, L. Clovamide and Flavonoids from Leaves of Trifolium pratense and T. pratense subsp. nivale Grown in Italy. Nat. Prod. Commun. 2015, 10, 933–936. [Google Scholar] [CrossRef] [PubMed]
- Stark, T.; Lang, R.; Keller, D.; Hense, A.; Hofmann, T. Absorption of N-Phenylpropenoyl-L-Amino Acids in Healthy Humans by Oral Administration of Cocoa (Theobroma cacao). Mol. Nutr. Food Res. 2008, 52, 1201–1214. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and Bioefficacy of Polyphenols in Humans. I. Review of 97 Bioavailability Studies. Am. J. Clin. Nutr. 2005, 81 (Suppl. 1), 230S–242S. [Google Scholar] [CrossRef]
- Vitaglione, P.; Lumaga, R.B.; Ferracane, R.; Sellitto, S.; Morelló, J.R.; Miranda, J.R.; Shimoni, E.; Fogliano, V. Human Bioavailability of Flavanols and Phenolic acids from Cocoa-Nut Creams Enriched with Free or Microencapsulated Cocoa Polyphenols. Br. J. Nutr. 2013, 109, 1832–1843. [Google Scholar] [CrossRef] [PubMed]
- Di Lorenzo, C.; Colombo, F.; Biella, S.; Stockley, C.; Restani, P. Polyphenols and Human Health: The Role of Bioavailability. Nutrients 2021, 13, 273. [Google Scholar] [CrossRef]
- Konishi, Y.; Kobayashi, S. Transepithelial Transport of Rosmarinic Acid in Intestinal Caco-2 Cell Monolayers. Biosci. Biotechnol. Biochem. 2005, 69, 583–591. [Google Scholar] [CrossRef] [PubMed]
- Baba, S.; Osakabe, N.; Natsume, M.; Yasuda, A.; Muto, Y.; Hiyoshi, T.; Takano, H.; Yoshikawa, T.; Terao, J. Absorption, Metabolism, Degradation and Urinary Excretion of Rosmarinic Acid After Intake of Perilla frutescens extract in Humans. Eur. J. Nutr. 2005, 44, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Rios, L.Y.; Gonthier, M.-P.; Rémésy, C.; Mila, I.; Lapierre, C.; Lazarus, S.A.; Williamson, G.; Scalbert, A. Chocolate Intake Increases Urinary Excretion of Polyphenol-Derived Phenolic Acids in Healthy Human Subjects. Am. J. Clin. Nutr. 2003, 77, 912–918. [Google Scholar] [CrossRef]
- Urpi-Sarda, M.; Llorach, R.; Khan, N.; Monagas, M.; Rotches-Ribalta, M.; Lamuela-Raventos, R.; Estruch, R.; Tinahones, F.J.; Andres-Lacueva, C. Effect of Milk on the Urinary Excretion of Microbial Phenolic Acids After Cocoa Powder Consumption in Humans. J. Agric. Food Chem. 2010, 58, 4706–4711. [Google Scholar] [CrossRef]
- Lugrin, J.; Rosenblatt-Velin, N.; Parapanov, R.; Liaudet, L. The Role of Oxidative Stress during Inflammatory Processes. Biol. Chem. 2014, 395, 203–230. [Google Scholar] [CrossRef] [PubMed]
- Higashi, Y. Roles of Oxidative Stress and Inflammation in Vascular Endothelial Dysfunction-Related Disease. Antioxidants 2022, 11, 1958. [Google Scholar] [CrossRef] [PubMed]
- Cecerska-Heryć, E.; Polikowska, A.; Serwin, N.; Roszak, M.; Grygorcewicz, B.; Heryć, R.; Michalczyk, A.; Dołęgowska, B. Importance of Oxidative Stress in the Pathogenesis, Diagnosis, and Monitoring of Patients with Neuropsychiatric Disorders, a Review. Neurochem. Int. 2022, 153, 105269. [Google Scholar] [CrossRef] [PubMed]
- Forman, H.J.; Zhang, H. Targeting Oxidative Stress in Disease: Promise and Limitations of Antioxidant Therapy. Nat. Rev. Drug Discov. 2021, 20, 689–709. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, K.; Aggarwal, B.B.; Singh, R.B.; Buttar, H.S.; Wilson, D.; De Meester, F. Food Antioxidants and Their Anti-Inflammatory Properties: A Potential Role in Cardiovascular Diseases and Cancer Prevention. Diseases 2016, 4, 28. [Google Scholar] [CrossRef] [PubMed]
- Sharifi-Rad, M.; Anil Kumar, N.V.; Zucca, P.; Varoni, E.M.; Dini, L.; Panzarini, E.; Rajkovic, J.; Tsouh Fokou, P.V.; Azzini, E.; Peluso, I.; et al. Lifestyle, Oxidative Stress, and Antioxidants: Back and Forth in the Pathophysiology of Chronic Diseases. Front. Physiol. 2020, 11, 694. [Google Scholar] [CrossRef] [PubMed]
- Rudrapal, M.; Khairnar, S.J.; Khan, J.; Bin Dukhyil, A.; Ansari, M.A.; Alomary, M.N.; Alshabrmi, F.M.; Palai, S.; Deb, P.K.; Devi, R. Dietary Polyphenols and Their Role in Oxidative Stress-Induced Human Diseases: Insights into Protective Effects, Antioxidant Potentials and Mechanism(s) of Action. Front. Pharmacol. 2022, 13, 806470. [Google Scholar] [CrossRef] [PubMed]
- Zeb, A. Concept, Mechanism, and Applications of Phenolic Antioxidants in Foods. J. Food Biochem. 2022, 44, e13394. [Google Scholar] [CrossRef] [PubMed]
- Madala, N.E.; Kabanda, M.M. LC-MS Based Validation and DFT Investigation on the Antioxidant Properties of Clovamide: •OH and •OOH Scavenging and Cu(II) Chelation Mechanisms. J. Mol. Struct. 2021, 15, 1236. [Google Scholar] [CrossRef]
- Locatelli, M.; Travaglia, F.; Giovannelli, L.; Coïsson, J.D.; Bordiga, M.; Pattarino, F.; Arlorio, M. Clovamide and Phenolics from Cocoa Beans (Theobroma cacao L.) Inhibit Lipid Peroxidation in Liposomal Systems. Food Res. Int. 2013, 50, 129–134. [Google Scholar] [CrossRef]
- Lotito, S.B.; Frei, B. Consumption of Flavonoid-Rich Foods and Increased Plasma Antioxidant Capacity in Humans: Cause, Consequence, or Epiphenomenon? Free. Radic. Biol. Med. 2006, 41, 1727–1746. [Google Scholar] [CrossRef] [PubMed]
- Frankel, E.N.; German, J.B. Antioxidants in Foods and Health: Problems and Fallacies in the Field. J. Sci. Food Agric. 2006, 86, 1999–2001. [Google Scholar] [CrossRef]
- Holst, B.; Williamson, G. Nutrients and Phytochemicals: From Bioavailability to Bioefficacy Beyond Antioxidants. Curr. Opin. Biotechnol. 2008, 19, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Hollman, P.C.; Cassidy, A.; Comte, B.; Heinonen, M.; Richelle, M.; Richling, E.; Serafini, M.; Scalbert, A.; Sies, H.; Vidry, S. The Biological Relevance of Direct Antioxidant Effects of Polyphenols for Cardiovascular Health in Humans Is Not Established. J. Nutr. 2011, 141, 989–1009. [Google Scholar] [CrossRef] [PubMed]
- Kotha, R.R.; Tareq, F.S.; Yildiz, E.; Luthria, D.L. Oxidative Stress and Antioxidants—A Critical Review on In Vitro Antioxidant Assays. Antioxidants 2022, 11, 2388. [Google Scholar] [CrossRef] [PubMed]
- Locatelli, M.; Gindro, R.; Travaglia, F.; Coïsson, J.-D.; Rinaldi, M.; Arlorio, M. Study of the DPPH-Scavenging Activity: Development of a Free Software for the Correct Interpretation of Data. Food Chem. 2009, 114, 889–897. [Google Scholar] [CrossRef]
- Marinova, E.; Georgiev, L.; Totseva, I.; Seizova, K.; Milkova, T. Antioxidant Activity and Mechanism of Action of Some Synthesised Phenolic acid Amides of Aromatic Amines. Czech J. Food Sci. 2013, 31, 5–13. [Google Scholar] [CrossRef]
- Sarr, S.O.; Gassama, A.; Manga, F.; Grellepois, F.; Lavaud, C. Synthesis and Study of Antioxidant Activities of Trans-(-)-Clovamide Derivatives. Am. J. Chem. Appl. 2018, 5, 58–63. [Google Scholar]
- Ley, J.P.; Bertram, H.-J. Synthesis of Lipophilic Clovamide Derivatives and Their Antioxidative Potential against Lipid Peroxidation. J. Agric. Food Chem. 2003, 51, 4596–4602. [Google Scholar] [CrossRef]
- Strobel, N.A.; Fassett, R.G.; Marsh, S.A.; Coombes, J.S. Oxidative Stress Biomarkers as Predictors of Cardiovascular Disease. Int. J. Cardiol. 2011, 147, 191–201. [Google Scholar] [CrossRef]
- Dubois-Deruy, E.; Peugnet, V.; Turkieh, A.; Pinet, F. Oxidative Stress in Cardiovascular Diseases. Antioxidants 2020, 9, 864. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.-Y.; Xu, X.; Li, X.-C. Cardiovascular Diseases: Oxidative Damage and Antioxidant Protection. Eur. Rev. Med. Pharmacol. Sci. 2014, 18, 3091–3096. [Google Scholar]
- Arlorio, M.; Coisson, J.D.; Travaglia, F.; Locatelli, M.; Bordiga, M.; Zamperone, A.; Pietronave, S.; Brunelleschi, S.; Prat, M. Radical Scavenging Capacity of Cocoa Polyphenols Triggers Anti-Inflammatory Properties in Human Monocytes and Al-lows Protective Effects on H9c2 Cardiomyoblast Exposed to Oxidative Stress. Pol. J. Food Nutr. Sci. 2011, 61, 22–23. [Google Scholar]
- Antonini, S.; Colangelo, D.; Oltolina, F.; Diena, M.; Arlorio, M.; Prat, M. Clovamide Protects Cardiac Progenitor Cells from H2O2-Induced Oxidative Stress. J. Appl. Biotechnol. Bioeng. 2018, 5, 1. [Google Scholar] [CrossRef]
- Kolodziejczyk, J.; Olas, B.; Wachowicz, B.; Szajwaj, B.; Stochmal, A.; Oleszek, W. Clovamide-Rich Extract from Trifolium pallidum Reduces Oxidative Stress-Induced Damage to Blood Platelets and Plasma. J. Physiol. Biochem. 2011, 67, 391–399. [Google Scholar] [CrossRef]
- Luo, C.; Zou, L.; Sun, H.; Peng, J.; Gao, C.; Bao, L.; Ji, R.; Jin, Y.; Sun, S. A Review of the Anti-Inflammatory Effects of Rosmarinic Acid on Inflammatory Diseases. Front. Pharmacol. 2020, 11, 153. [Google Scholar] [CrossRef]
- Rocha, J.; Eduardo-Figueira, M.; Barateiro, A.; Fernandes, A.; Brites, D.; Bronze, R.; Duarte, C.M.; Serra, A.T.; Pinto, R.; Freitas, M.; et al. Anti-Inflammatory Effect of Rosmarinic Acid and an Extract of Rosmarinus officinalis in Rat Models of Local and Systemic Inflammation. Basic Clin. Pharmacol. Toxicol. 2015, 116, 398–413. [Google Scholar] [CrossRef] [PubMed]
- Noor, S.; Mohammad, T.; Rub, M.A.; Raza, A.; Azum, N.; Yadav, D.K.; Hassan, M.I.; Asiri, A.M. Biomedical Features and Therapeutic Potential of Rosmarinic Acid. Arch. Pharmacal Res. 2022, 45, 205–228. [Google Scholar] [CrossRef]
- Zeng, H.; Locatelli, M.; Bardelli, C.; Amoruso, A.; Coisson, J.D.; Travaglia, F.; Arlorio, M.; Brunelleschi, S. Anti-Inflammatory Properties of Clovamide and Theobroma cacao Phenolic Extracts in Human Monocytes: Evaluation of Respiratory Burst, Cytokine Release, NF-κB Activation, and PPARγ Modulation. J. Agric. Food Chem. 2011, 59, 5342–5350. [Google Scholar] [CrossRef]
- Liu, R.; Guo, Y.; Yu, J.; Wei, X.; Zhou, F.; Yuan, X.; Cai, L.; Yu, C. Protective effect of N-(E)-p-Coumaroyltyrosine on LPS-Induced acute Inflammatory Injury and Signaling Pathway Analysis. Fish Shellfish Immunol. 2024, 144, 109242. [Google Scholar] [CrossRef]
- Nichols, E.; Vos, T. Estimating the Global Mortality from Alzheimer’s Disease and other Dementias: A New Method and Results from the Global Burden of Disease Study 2019. Alzheimer’s Dement. 2020, 16, e042236. [Google Scholar] [CrossRef]
- Tyler, S.E.B.; Tyler, L.D.K. Pathways to Healing: Plants with Therapeutic Potential for Neurodegenerative Diseases. IBRO Neurosci. Rep. 2023, 14, 210–234. [Google Scholar] [CrossRef] [PubMed]
- Cimini, A.; Gentile, R.; D’Angelo, B.; Benedetti, E.; Cristiano, L.; Avantaggiati, M.L.; Giordano, A.; Ferri, C.; Desideri, G. Cocoa Powder Triggers Neuroprotective and Preventive Effects in a Human Alzheimer’s Disease Model by Modulating BDNF Signaling Pathway. J. Cell Biochem. 2013, 114, 2209–2220. [Google Scholar] [CrossRef] [PubMed]
- Barrera-Reyes, P.K.; de Lara, J.C.-F.; González-Soto, M.; Tejero, M.E. Effects of Cocoa-Derived Polyphenols on Cognitive Function in Humans. Systematic Review and Analysis of Methodological Aspects. Plant Foods Hum. Nutr. 2020, 75, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Fallarini, S.; Miglio, G.; Paoletti, T.; Minassi, A.; Amoruso, A.; Bardelli, C.; Brunelleschi, S.; Lombardi, G. Clovamide and Rosmarinic Acid Induce Neuroprotective Effects in In Vitro Models of Neuronal Death. Br. J. Pharmacol. 2009, 157, 1072–1084. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.-H.; Hu, X.-L.; Lv, X.-Y.; Wang, B.-L.; Lin, J.; Zhang, X.-Q.; Ye, W.-C.; Xiong, F.; Wang, H. Synthesis and Biological Evaluation of Clovamide Analogues with Catechol Functionality as Potent Parkinson’s Disease Agents In Vitro and In Vivo. Bioorganic Med. Chem. Lett. 2019, 29, 302–312. [Google Scholar] [CrossRef] [PubMed]
- Tsunoda, T.; Takase, M.; Shigemori, H. Structure-Activity Relationship of Clovamide and Its Related Compounds for the Inhibition of Amyloid β Aggregation. Bioorganic Med. Chem. 2018, 26, 3202–3209. [Google Scholar] [CrossRef] [PubMed]
- Nomoto, D.; Tsunoda, T.; Shigemori, H. Effects of Clovamide and Its Related Compounds on the Aggregations of Amyloid Polypeptides. J. Nat. Med. 2021, 75, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Betkekar, V.V.; Ohmori, K.; Suzuki, K.; Shigemori, H. Evaluation of Amyloid Polypeptide Aggregation Inhibition and Disaggregation Activity of A-Type Procyanidins. Pharmaceuticals 2021, 14, 1118. [Google Scholar] [CrossRef]
- Li, J.; Zhu, M.; Manning-Bog, A.B.; Di Monte, D.A.; Fink, A.L. Dopamine and L-DOPA Disaggregate Amyloid Fibrils: Implications for Parkinson’s and Alzheimer’s Disease. FASEB J. 2004, 18, 962–964. [Google Scholar] [CrossRef]
- Hase, T.; Shishido, S.; Yamamoto, S.; Yamashita, R.; Nukima, H.; Taira, S.; Toyoda, T.; Abe, K.; Hamaguchi, T.; Ono, K.; et al. Rosmarinic acid Suppresses Alzheimer’s Disease Development by Reducing Amyloid β Aggregation by Increasing Monoamine secretion. Sci. Rep. 2019, 9, 8711. [Google Scholar] [CrossRef]
- Park, J.-Y.; Kim, B.-W.; Lee, H.U.; Choi, D.-K.; Yoon, S.-H. Synthesis of Clovamide Analogues That Inhibit NO Production in Activated BV-2 Microglial Cells. Biol. Pharm. Bull. 2017, 40, 1475–1482. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.-L.; Lin, J.; Lv, X.-Y.; Feng, J.-H.; Zhang, X.-Q.; Wang, H.; Ye, W.-C. Synthesis and Biological Evaluation of Clovamide Analogues as Potent Anti-Neuroinflammatory Agents in Vitro and In Vivo. Eur. J. Med. Chem. 2018, 151, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Min, S.; More, S.V.; Park, J.Y.; Jeon, S.B.; Park, S.Y.; Park, E.J.; Yoon, S.H.; Choi, D.K. EOP, A Newly Synthesized Ethyl Pyruvate Derivative, Attenuates the Production of Inflammatory Mediators via p38, ERK and NF-κB Pathways in Lipo-polysaccharide-Activated BV-2 Microglial Cells. Molecules 2014, 19, 19361–19375. [Google Scholar] [CrossRef] [PubMed]
- Yarza, R.; Vela, S.; Solas, M.; Ramirez, M.J. c-Jun N-terminal Kinase (JNK) Signaling as a Therapeutic Target for Alzheimer’s Disease. Front. Pharmacol. 2016, 6, 321. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.W.; Park, J.I.; More, S.V.; Park, J.Y.; Kim, B.W.; Jeon, S.B.; Yun, Y.S.; Park, E.J.; Yoon, S.H.; Choi, D.K. Anti-Neuroinflammatory Effects of DPTP, A Novel Synthetic Clovamide Derivative in In Vitro and In Vivo Model of Neuroinflammation. Brain Res. Bull. 2015, 112, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Nignpense, B.E.; Chinkwo, K.A.; Blanchard, C.L.; Santhakumar, A.B. Polyphenols: Modulators of Platelet Function and Platelet Microparticle Generation? Int. J. Mol. Sci. 2019, 21, 146. [Google Scholar] [CrossRef] [PubMed]
- Ludovici, V.; Barthelmes, J.; Nagele, M.P.; Flammer, A.J.; Sudano, I. Polyphenols: Anti-Platelet Nutraceutical? Curr. Pharm. Des. 2018, 24, 146–157. [Google Scholar] [CrossRef]
- Nam, G.S.; Park, H.J.; Nam, K.S. The Antithrombotic Effect of Caffeic Acid Is Associated with A cAMP-Dependent Pathway and Clot Retraction in Human Platelets. Thromb. Res. 2020, 195, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Park, J.B. Caffedymine from Cocoa Has COX Inhibitory Activity Suppressing the Expression of a Platelet Activation Marker, P-Selectin. J. Agric. Food Chem. 2007, 55, 2171–2175. [Google Scholar] [CrossRef]
- Park, J.B.; Schoene, N. Clovamide-Type Phenylpropenoic Acid Amides, N-Coumaroyldopamine and N-Caffeoyldopamine, Inhibit Platelet-Leukocyte Interactions via Suppressing P-Selectin Expression. J. Pharmacol. Exp. Ther. 2006, 317, 813–819. [Google Scholar] [CrossRef]
- Park, J.B.; Schoene, N. Synthesis and Characterization of N-Coumaroyltyramine as a Potent Phytochemical Which Arrests Human Transformed Cells via Inhibiting Protein Tyrosine Kinases. Biochem. Biophys. Res. Commun. 2002, 292, 1104–1110. [Google Scholar] [CrossRef] [PubMed]
- Wongsakul, A.; Lertnitikul, N.; Suttisri, R.; Jianmongkol, S. N-Trans-p-Coumaroyltyramine Enhances Indomethacin- and Diclofenac-induced Apoptosis Through Endoplasmic Reticulum Stress-dependent Mechanism in MCF-7 Cells. Anticancer Res. 2022, 42, 1833–1844. [Google Scholar] [CrossRef] [PubMed]
- Peraza-Labrador, A.; Buitrago, D.M.; Coy-Barrera, E.; Perdomo-Lara, S.J. Antiproliferative and Pro-Apoptotic Effects of a Phenolic-Rich Extract from Lycium barbarum Fruits on Human Papillomavirus (HPV) 16-Positive Head Cancer Cell Lines. Molecules 2022, 27, 3568. [Google Scholar] [CrossRef] [PubMed]
- Ye, N.; Caruso, F.; Rossi, M. Mechanistic Insights into the Inhibition of SARS-CoV-2 Main Protease by Clovamide and Its Derivatives: In Silico Studies. Biophysica 2021, 1, 28. [Google Scholar] [CrossRef]
- Niehues, M.; Stark, T.; Keller, D.; Hofmann, T.; Hensel, A. Antiadhesion as a Functional Concept for Prevention of Pathogens: N-phenylpropenoyl-L-amino Acid Amides as Inhibitors of the Helicobacter pylori BabA outer membrane protein. Mol. Nutr. Food Res. 2011, 55, 1104–1117. [Google Scholar] [CrossRef]
- Canivenc-Lavier, M.-C.; Bennetau-Pelissero, C. Phytoestrogens and Health Effects. Nutrients 2023, 15, 317. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.-N.; Lin, C.-C.; Liu, C.-F. Efficacy of Phytoestrogens for Menopausal Symptoms: A Meta-Analysis and Systematic Review. Climacteric 2015, 18, 260–269. [Google Scholar] [CrossRef]
- Patra, S.; Gorai, S.; Pal, S.; Ghosh, K.; Pradhan, S.; Chakrabarti, S. A Review on Phytoestrogens: Current Status and Future Direction. Phytother. Res. 2023, 37, 3097–3120. [Google Scholar] [CrossRef]
- Powers, C.N.; Setzer, W.N. A Molecular Docking Study of Phytochemical Estrogen Mimics from Dietary Herbal Supplements. Silico Pharmacol. 2015, 3, 4. [Google Scholar] [CrossRef]
- Marinho, S.; Illanes, M.; Ávila-Román, J.; Motilva, V.; Talero, E. Anti-Inflammatory Effects of Rosmarinic Acid-Loaded Nanovesicles in Acute Colitis through Modulation of NLRP3 Inflammasome. Biomolecules 2021, 11, 162. [Google Scholar] [CrossRef]




| Type of Sample | Clovamide Content | References |
|---|---|---|
| mg/g of the product | ||
| Raw beans | 0.052 | [23] |
| Roasted beans | 0.044 | |
| Side products (winnowed) | 0.024 | |
| End products (winnowed) | 0.065 | |
| mg/g of cocoa powder | ||
| Unroasted beans, of Ghana origin | 0.0026 | [24] |
| Roasted beans, of Ghana origin | 0.0012 | |
| Unroasted beans, of Arriba origin | 0.0013 | |
| Roasted beans, of Arriba origin | 0.0005 | |
| Unroasted beans, of Ivory Coast origin | 0.0021 | |
| Roasted beans, of Ivory Coast origin | 0.0011 | |
| mg/g in defatted raw beans | ||
| 18 samples of cocoa beans, originating from 12 countries, 4 continents | 0.12–0.37 | [25] |
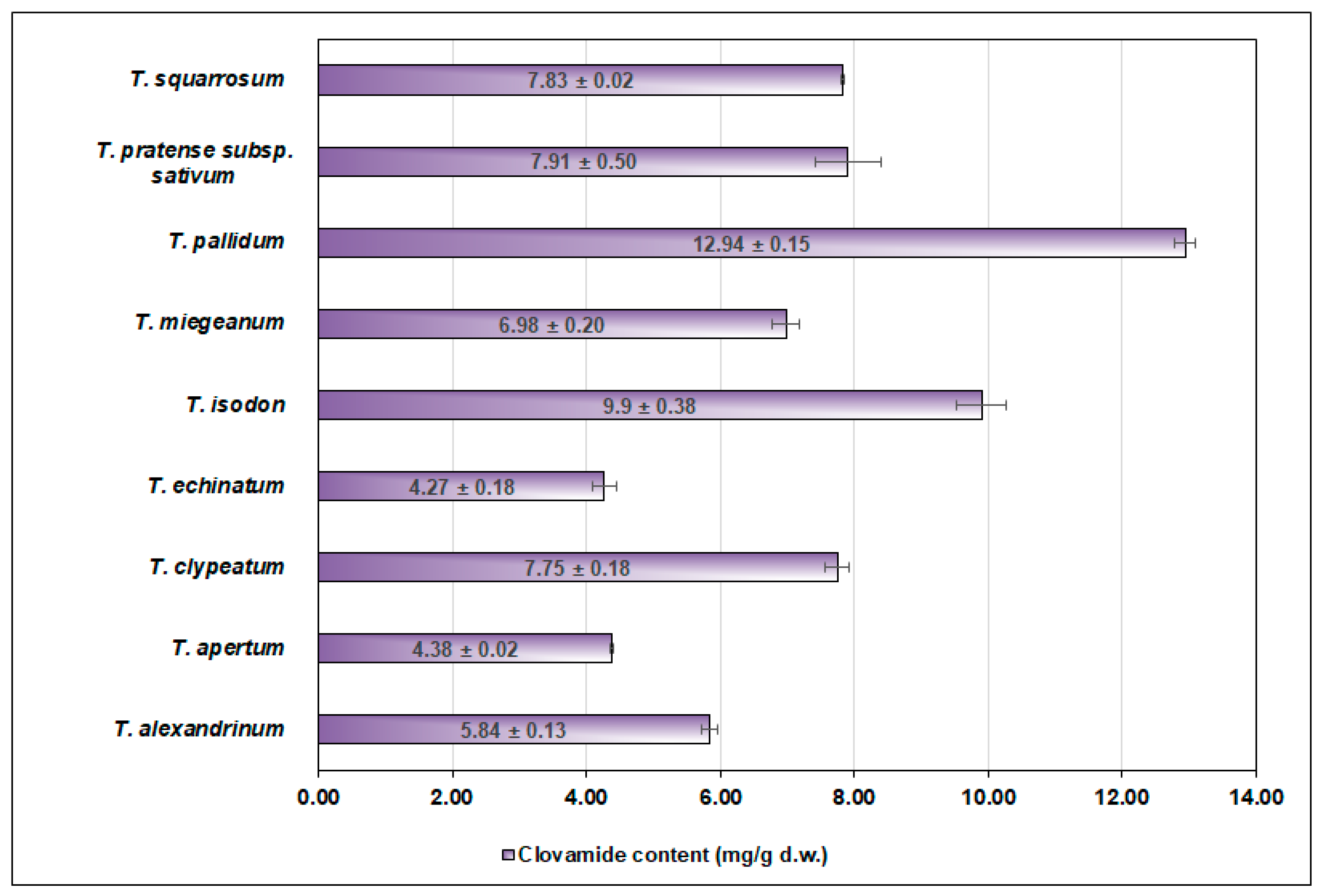
| Tifolium Species | Clovamide % in the Total Phenolics Content |
|---|---|
| T. alexandrinum L. | 21.54 |
| T. apertum Bobrov | 16.06 |
| T. clypeatum L. | 28.32 |
| T. echinatum M.Bieb. | 13.52 |
| T. isodon Murb. | 18.07 |
| T. miegeanum Maire | 14.20 |
| T. pallidum Waldst. & Kit | 36.90 |
| T. pratense subsp. sativum (Schreb.) Ser. | 19.61 |
| T. squarrosum L. | 15.12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kolodziejczyk-Czepas, J. Clovamide and Its Derivatives—Bioactive Components of Theobroma cacao and Other Plants in the Context of Human Health. Foods 2024, 13, 1118. https://doi.org/10.3390/foods13071118
Kolodziejczyk-Czepas J. Clovamide and Its Derivatives—Bioactive Components of Theobroma cacao and Other Plants in the Context of Human Health. Foods. 2024; 13(7):1118. https://doi.org/10.3390/foods13071118
Chicago/Turabian StyleKolodziejczyk-Czepas, Joanna. 2024. "Clovamide and Its Derivatives—Bioactive Components of Theobroma cacao and Other Plants in the Context of Human Health" Foods 13, no. 7: 1118. https://doi.org/10.3390/foods13071118
APA StyleKolodziejczyk-Czepas, J. (2024). Clovamide and Its Derivatives—Bioactive Components of Theobroma cacao and Other Plants in the Context of Human Health. Foods, 13(7), 1118. https://doi.org/10.3390/foods13071118