
New Approach Methods to Assess the Enteropathogenic Potential of Strains of the Bacillus cereus Group, including Bacillus thuringiensis

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Characterization of the Bacterial Dataset
2.2.1. Bt Species Attribution
2.2.2. PanC-Typing and Detection of Toxin Genes
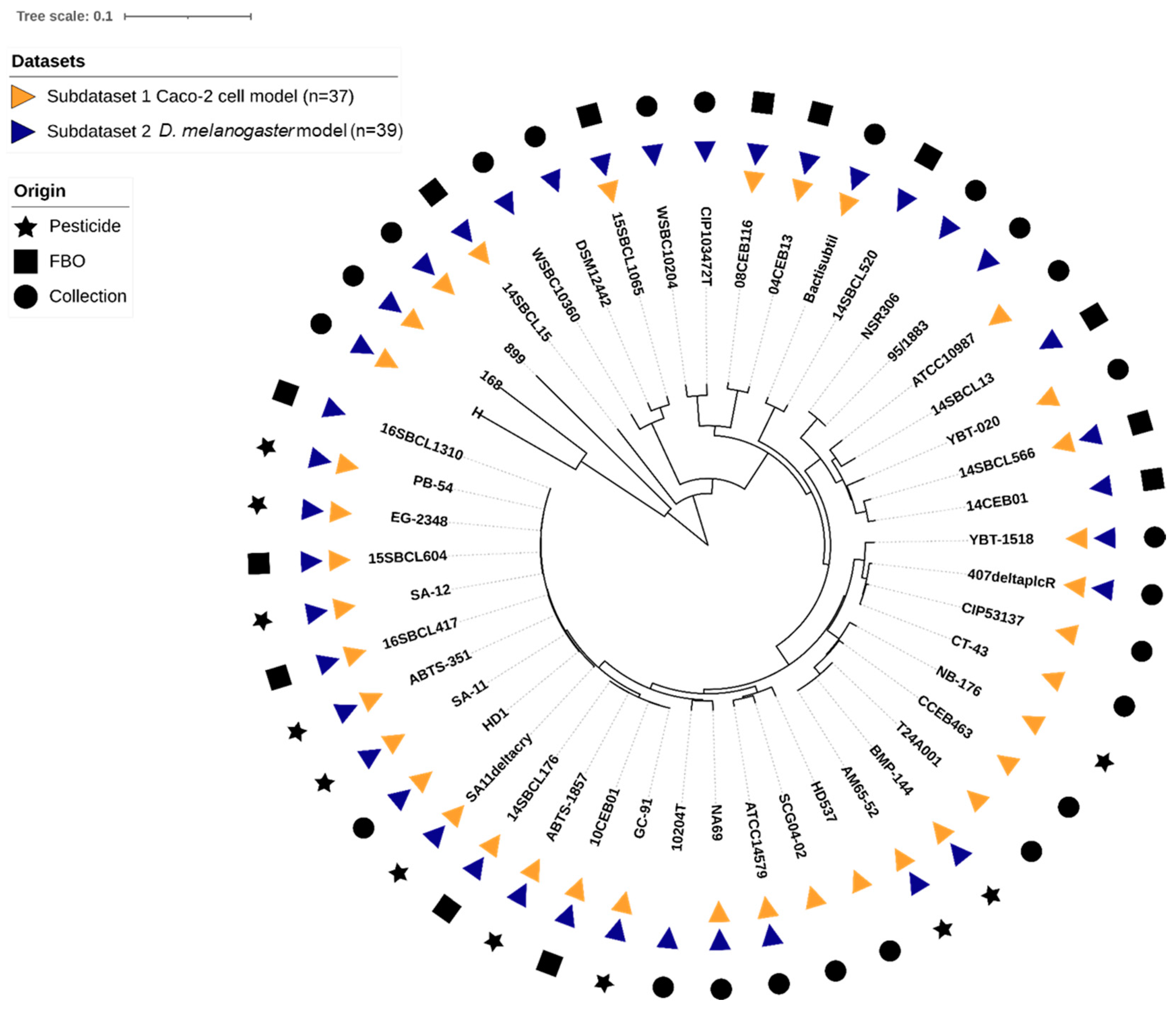
2.2.3. Bacterial Dataset Phylogeny
2.3. Bacterial Culture for Supernatant Production
2.4. Spore Production
2.5. Toxicity Assays on Caco-2 Cells
2.5.1. Cell Line
2.5.2. Cell Treatment
2.5.3. Cytotoxicity
2.5.4. IL-8 Release
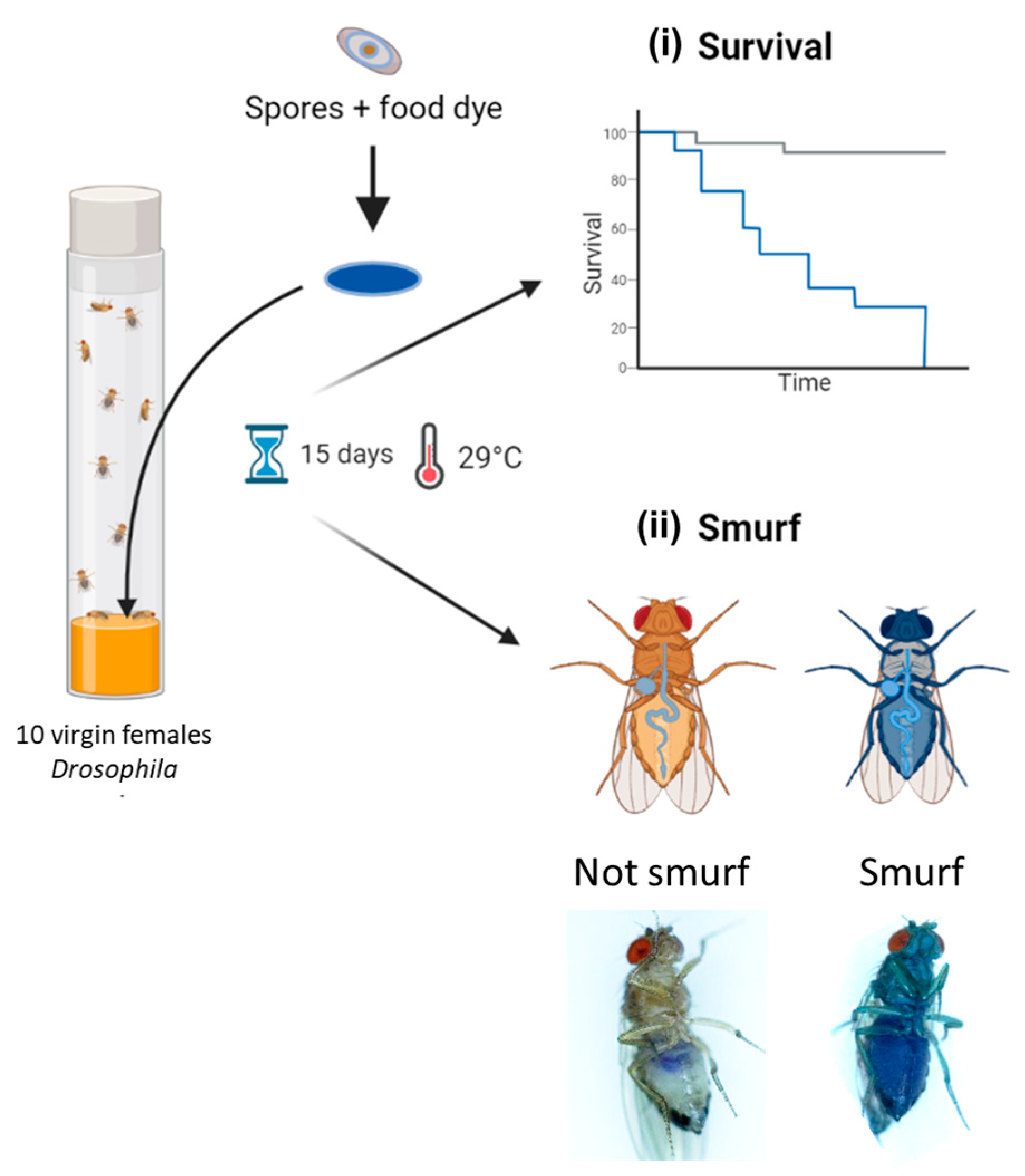
2.6. Drosophila melanogaster Rearing and Enterotoxicity Assays
3. Results
3.1. Description of the Bacterial Strain Panel
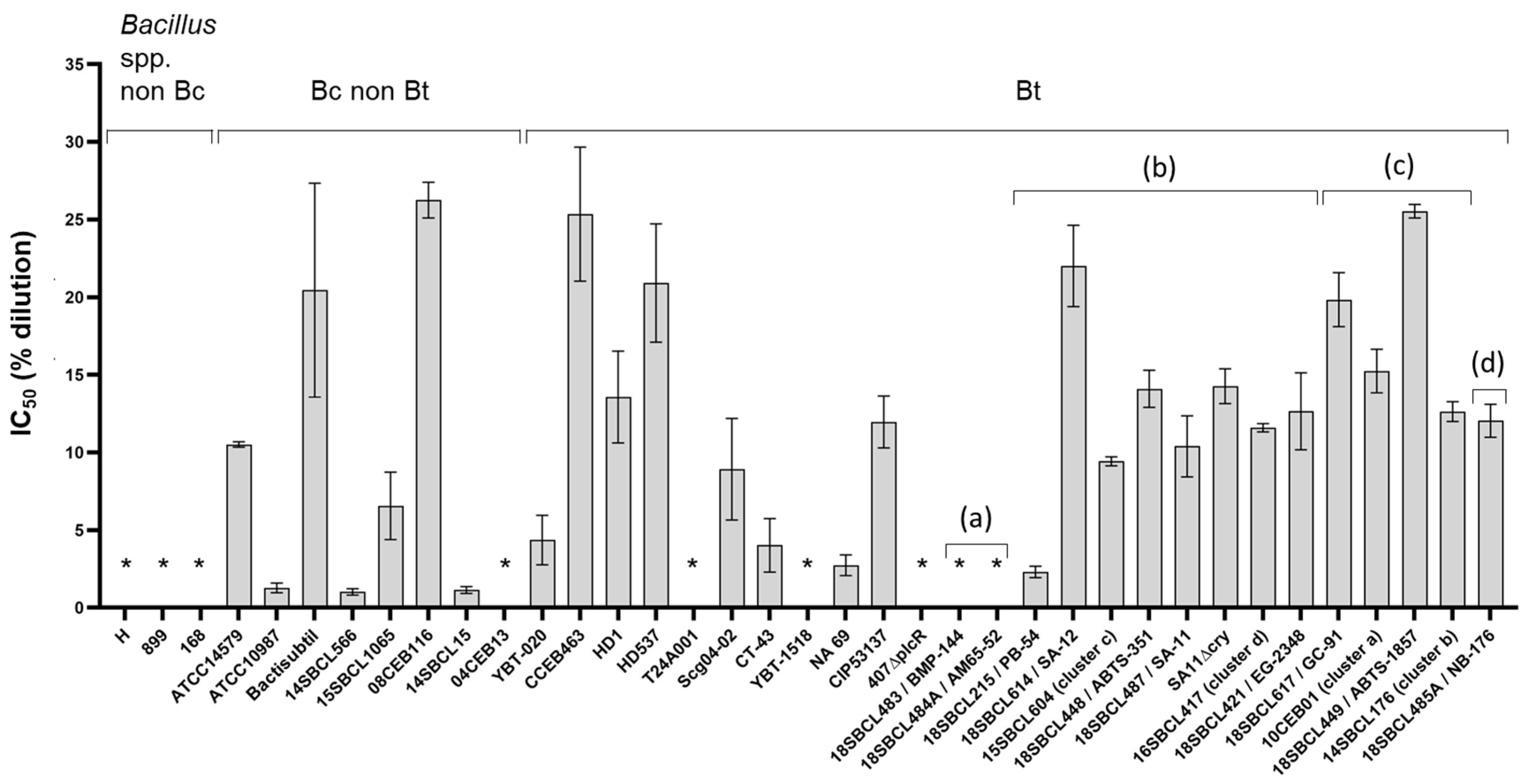
3.2. Caco-2 Cytotoxicity of Bacillus spp. Supernatants
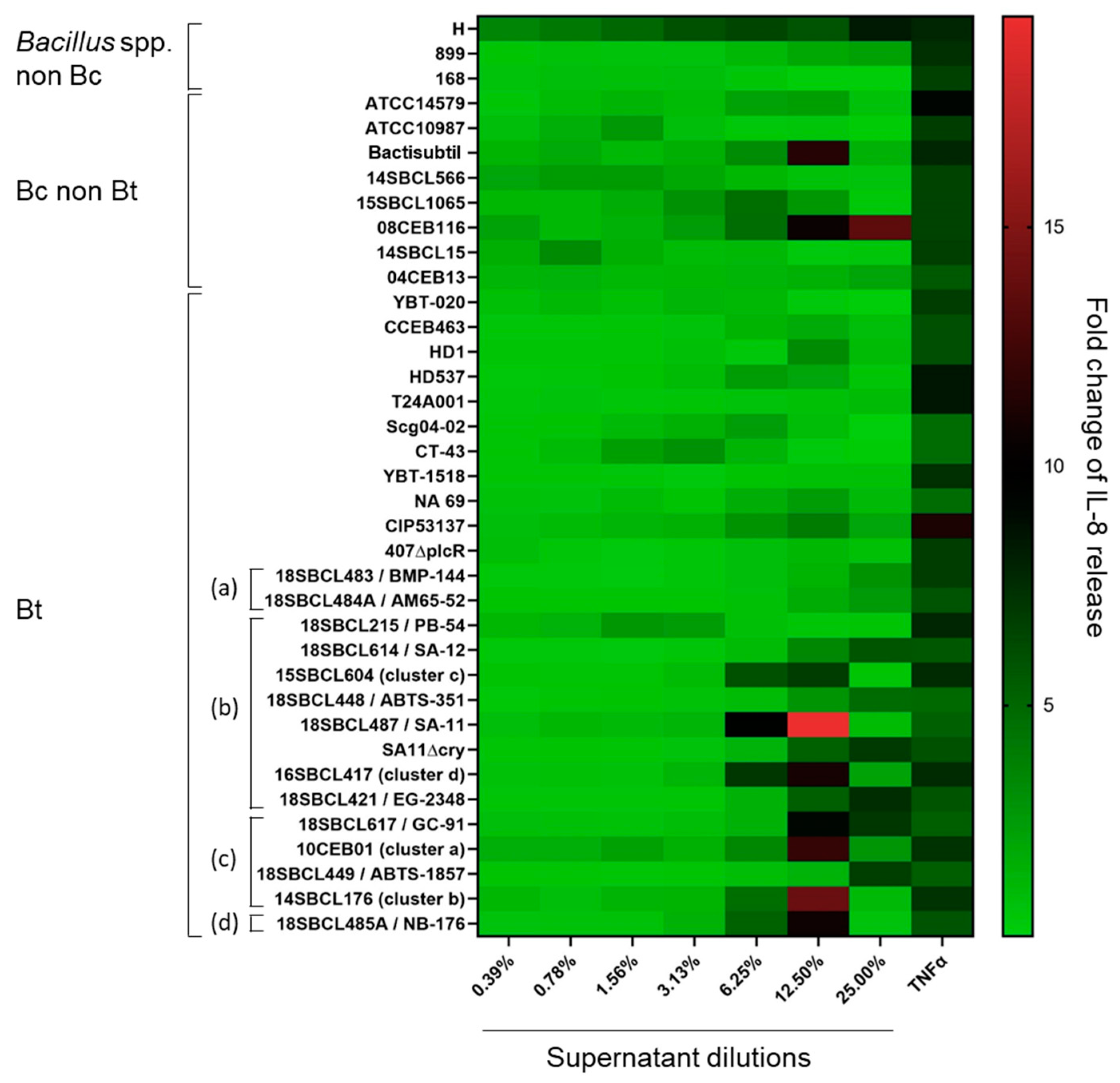
3.3. Pro-Inflammatory Effect of Bacillus spp. Supernatants
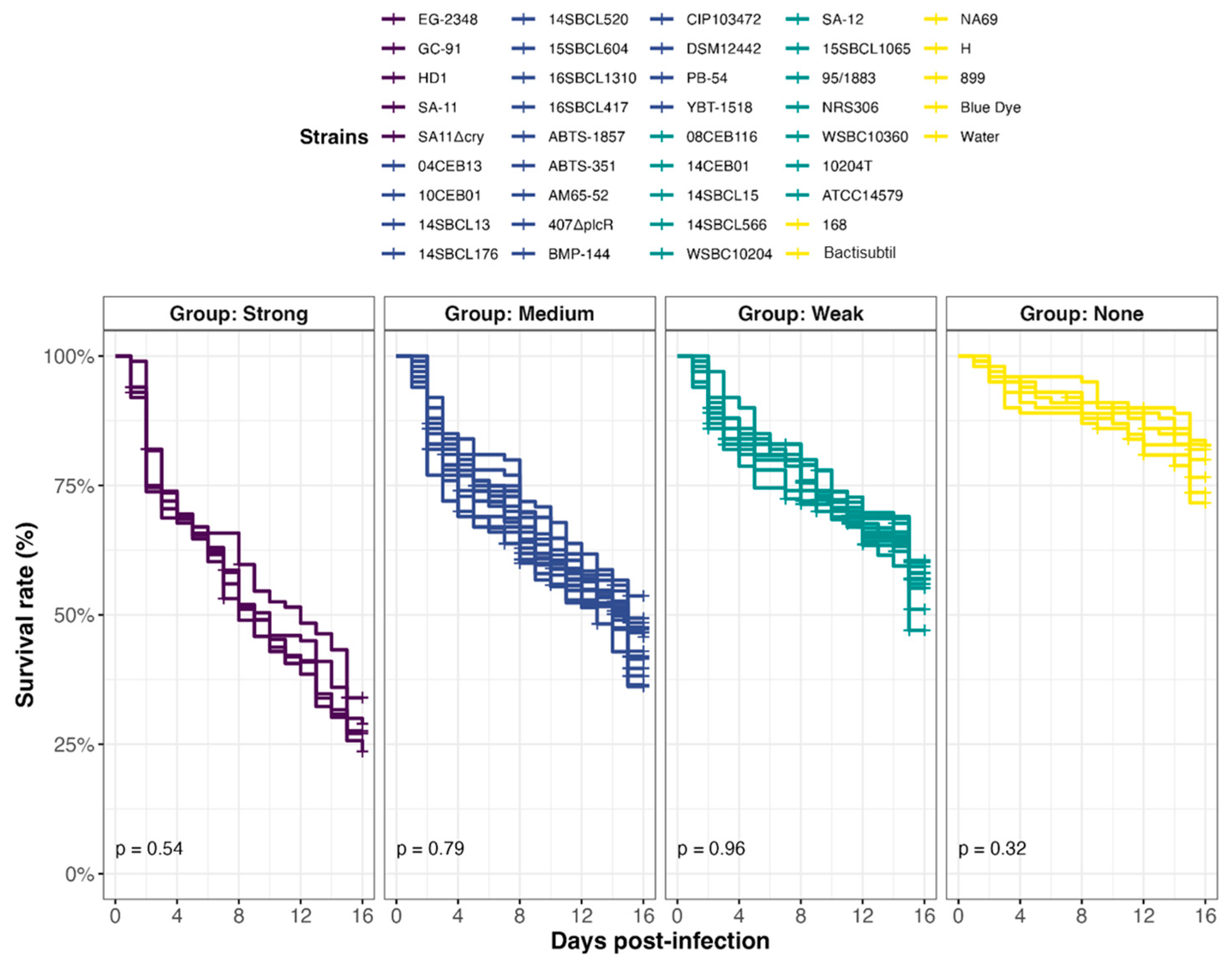
3.4. Effect of the Ingestion of Bacillus spp. Spores on D. melanogaster Lethality
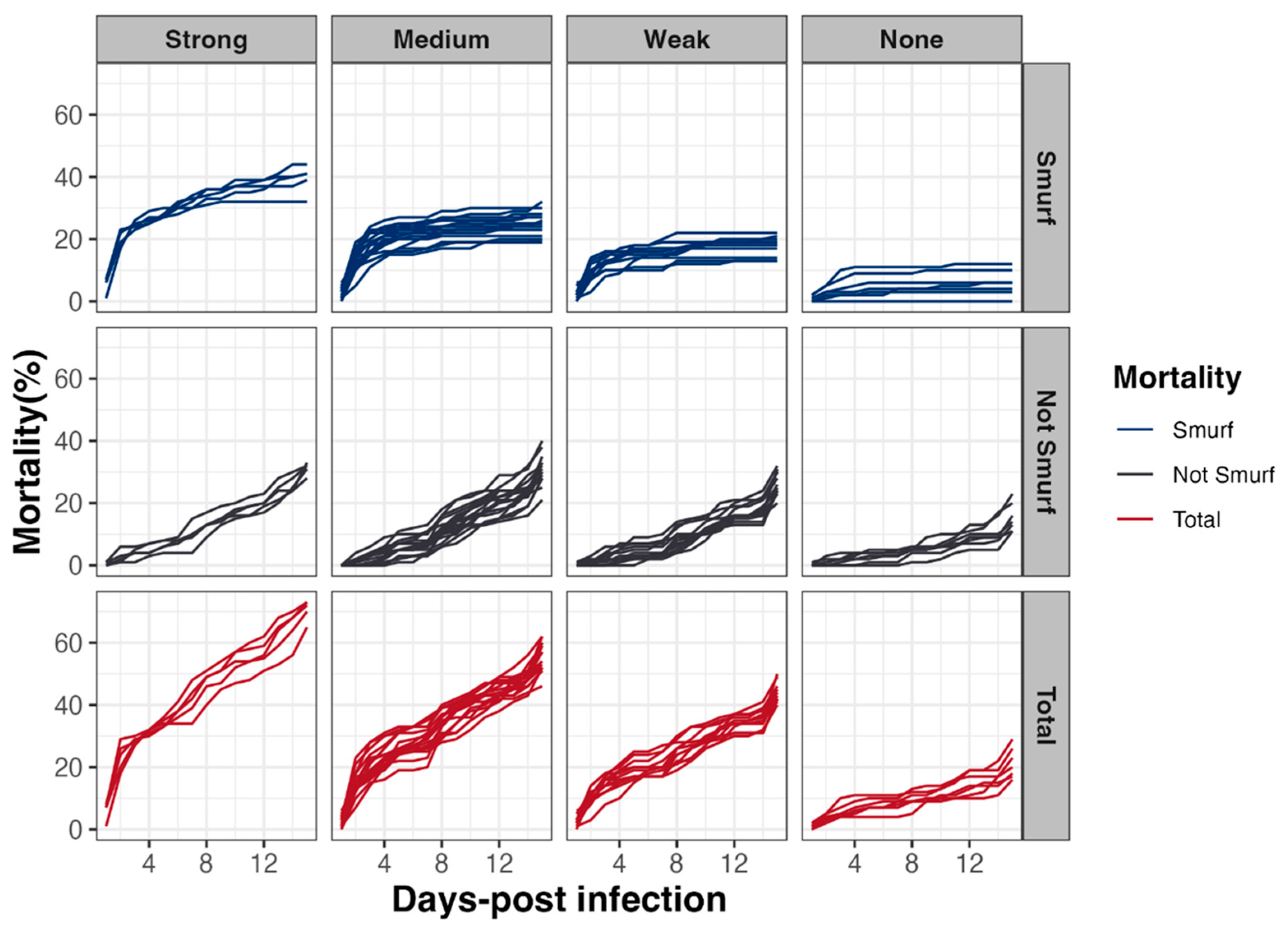
3.5. Early Mortality of D. melanogaster Correlates with Intestinal Leakiness
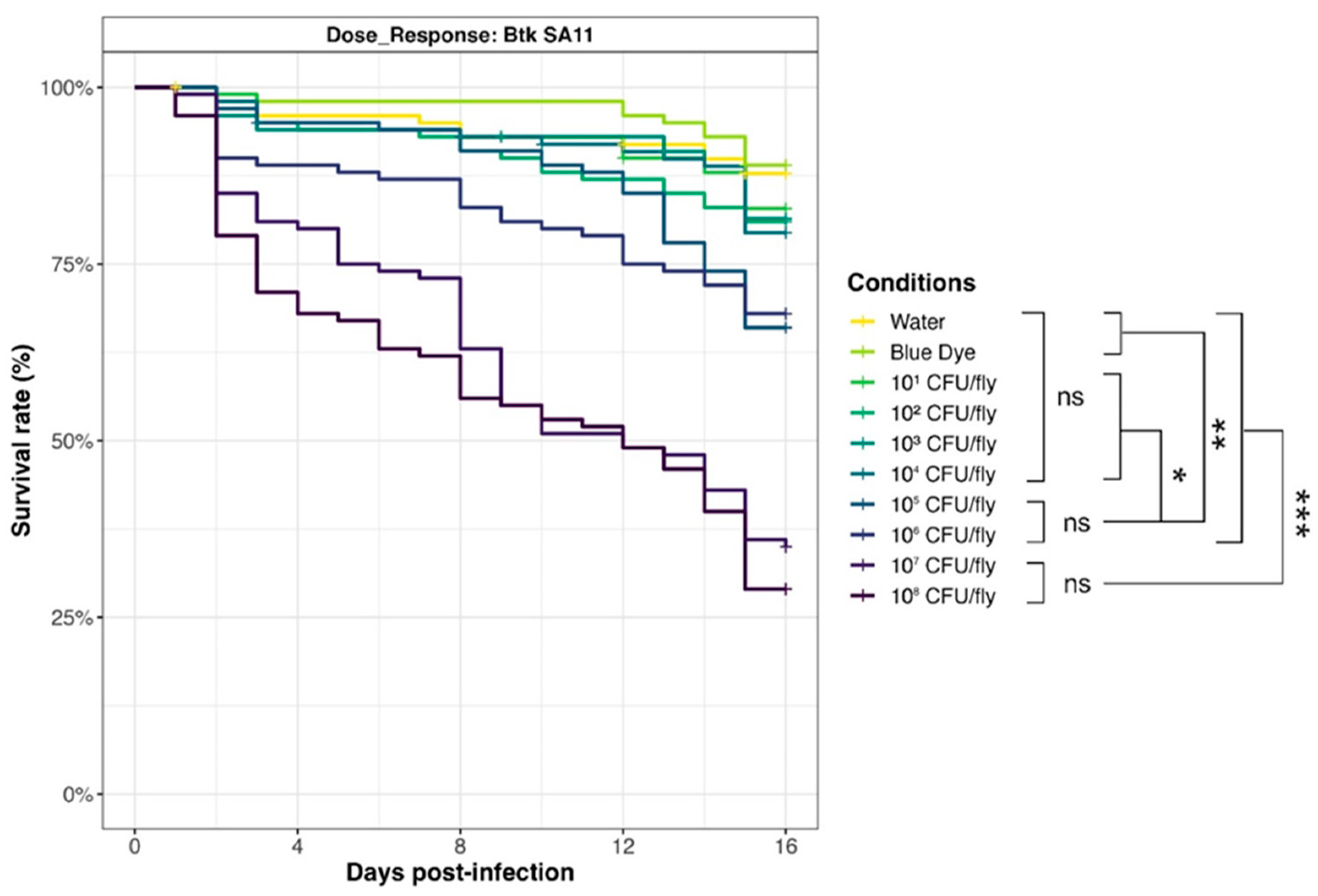
3.6. Drosophila melanogaster Exposure–Response Relationship
3.7. Comparison of Toxicity Results from the Two Experimental Models
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Enosi Tuipulotu, D.; Mathur, A.; Ngo, C.; Man, S.M. Bacillus cereus: Epidemiology, Virulence Factors, and Host-Pathogen Interactions. Trends Microbiol. 2021, 29, 458–471. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.A.; Beno, S.M.; Kent, D.J.; Carroll, L.M.; Martin, N.H.; Boor, K.J.; Kovac, J. Bacillus wiedmannii sp. nov., a psychrotolerant and cytotoxic Bacillus cereus group species isolated from dairy foods and dairy environments. Int. J. Syst. Evol. Microbiol. 2016, 66, 4744–4753. [Google Scholar] [CrossRef] [PubMed]
- Guinebretiere, M.H.; Thompson, F.L.; Sorokin, A.; Normand, P.; Dawyndt, P.; Ehling-Schulz, M.; Svensson, B.; Sanchis, V.; Nguyen-The, C.; Heyndrickx, M.; et al. Ecological diversification in the Bacillus cereus Group. Environ. Microbiol. 2008, 10, 851–865. [Google Scholar] [CrossRef] [PubMed]
- Didelot, X.; Barker, M.; Falush, D.; Priest, F.G. Evolution of pathogenicity in the Bacillus cereus group. Syst. Appl. Microbiol. 2009, 32, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Carroll, L.M.; Wiedmann, M.; Kovac, J. Proposal of a Taxonomic Nomenclature for the Bacillus cereus Group Which Reconciles Genomic Definitions of Bacterial Species with Clinical and Industrial Phenotypes. mBio 2020, 11, e00034-20. [Google Scholar] [CrossRef] [PubMed]
- Shimoyama, Y.; Umegaki, O.; Ooi, Y.; Agui, T.; Kadono, N.; Minami, T. Bacillus cereus pneumonia in an immunocompetent patient: A case report. JA Clin. Rep. 2017, 3, 25. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Li, Y.Y.; Xu, Y.; Sun, B.J.; Shao, J.; Zhang, D.; Li, K.; Fan, D.D.; Xue, Z.B.; Chen, W.H.; et al. Molecular Signatures Related to the Virulence of Bacillus cereus Sensu Lato, a Leading Cause of Devastating Endophthalmitis. mSystems 2019, 4, e00745-19. [Google Scholar] [CrossRef] [PubMed]
- Lotte, R.; Herisse, A.L.; Berrouane, Y.; Lotte, L.; Casagrande, F.; Landraud, L.; Herbin, S.; Ramarao, N.; Boyer, L.; Ruimy, R. Virulence Analysis of Bacillus cereus Isolated after Death of Preterm Neonates, Nice, France, 2013. Emerg. Infect. Dis. 2017, 23, 845–848. [Google Scholar] [CrossRef]
- SPF. Surveillance des Toxi-Infections Alimentaires Collectives—Données de la Déclaration Obligatoire, 2021; Santé Publique France: Saint-Maurice, France, 2023.
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union One Health 2022 Zoonoses Report. EFSA J. 2023, 21, e8442. [Google Scholar] [CrossRef]
- Agata, N.; Ohta, M.; Mori, M.; Isobe, M. A novel dodecadepsipeptide, cereulide, is an emetic toxin of Bacillus cereus. FEMS Microbiol. Lett. 1995, 129, 17–20. [Google Scholar] [CrossRef] [PubMed]
- Thery, M.; Cousin, V.L.; Tissieres, P.; Enault, M.; Morin, L. Multi-organ failure caused by lasagnas: A case report of Bacillus cereus food poisoning. Front. Pediatr. 2022, 10, 978250. [Google Scholar] [CrossRef] [PubMed]
- Naranjo, M.; Denayer, S.; Botteldoorn, N.; Delbrassinne, L.; Veys, J.; Waegenaere, J.; Sirtaine, N.; Driesen, R.B.; Sipido, K.R.; Mahillon, J.; et al. Sudden death of a young adult associated with Bacillus cereus food poisoning. J. Clin. Microbiol. 2011, 49, 4379–4381. [Google Scholar] [CrossRef] [PubMed]
- Beecher, D.J.; Macmillan, J.D. Characterization of the components of hemolysin BL from Bacillus cereus. Infect. Immun. 1991, 59, 1778–1784. [Google Scholar] [CrossRef] [PubMed]
- Lund, T.; Granum, P.E. Characterisation of a non-haemolytic enterotoxin complex from Bacillus cereus isolated after a foodborne outbreak. FEMS Microbiol. Lett. 1996, 141, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Lund, T.; De Buyser, M.L.; Granum, P.E. A new cytotoxin from Bacillus cereus that may cause necrotic enteritis. Mol. Microbiol. 2000, 38, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Jessberger, N.; Krey, V.M.; Rademacher, C.; Bohm, M.E.; Mohr, A.K.; Ehling-Schulz, M.; Scherer, S.; Martlbauer, E. From genome to toxicity: A combinatory approach highlights the complexity of enterotoxin production in Bacillus cereus. Front. Microbiol. 2015, 6, 560. [Google Scholar] [CrossRef]
- Tran, S.L.; Guillemet, E.; Gohar, M.; Lereclus, D.; Ramarao, N. CwpFM (EntFM) is a Bacillus cereus potential cell wall peptidase implicated in adhesion, biofilm formation, and virulence. J. Bacteriol. 2010, 192, 2638–2642. [Google Scholar] [CrossRef] [PubMed]
- Fox, D.; Mathur, A.; Xue, Y.; Liu, Y.; Tan, W.H.; Feng, S.; Pandey, A.; Ngo, C.; Hayward, J.A.; Atmosukarto, I.I.; et al. Bacillus cereus non-haemolytic enterotoxin activates the NLRP3 inflammasome. Nat. Commun. 2020, 11, 760. [Google Scholar] [CrossRef] [PubMed]
- Andersson, A.; Granum, P.E.; Ronner, U. The adhesion of Bacillus cereus spores to epithelial cells might be an additional virulence mechanism. Int. J. Food Microbiol. 1998, 39, 93–99. [Google Scholar] [CrossRef]
- Ramarao, N.; Lereclus, D. Adhesion and cytotoxicity of Bacillus cereus and Bacillus thuringiensis to epithelial cells are FlhA and PlcR dependent, respectively. Microbes Infect. 2006, 8, 1483–1491. [Google Scholar] [CrossRef] [PubMed]
- Mazzantini, D.; Celandroni, F.; Salvetti, S.; Gueye, S.A.; Lupetti, A.; Senesi, S.; Ghelardi, E. FlhF Is Required for Swarming Motility and Full Pathogenicity of Bacillus cereus. Front. Microbiol. 2016, 7, 1644. [Google Scholar] [CrossRef] [PubMed]
- Ceuppens, S.; Uyttendaele, M.; Drieskens, K.; Rajkovic, A.; Boon, N.; Wiele, T.V. Survival of Bacillus cereus vegetative cells and spores during in vitro simulation of gastric passage. J. Food Prot. 2012, 75, 690–694. [Google Scholar] [CrossRef] [PubMed]
- Clavel, T.; Carlin, F.; Lairon, D.; Nguyen-The, C.; Schmitt, P. Survival of Bacillus cereus spores and vegetative cells in acid media simulating human stomach. J. Appl. Microbiol. 2004, 97, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Hachfi, S.; Brun-Barale, A.; Munro, P.; Nawrot-Esposito, M.-P.; Michel, G.; Fichant, A.; Bonis, M.; Ruimy, R.; Boyer, L.; Gallet, A. Ingestion of Bacillus cereus spores dampens the immune response to favor bacterial persistence. bioRxiv 2023. [Google Scholar] [CrossRef]
- Rolny, I.S.; Minnaard, J.; Racedo, S.M.; Perez, P.F. Murine model of Bacillus cereus gastrointestinal infection. J. Med. Microbiol. 2014, 63, 1741–1749. [Google Scholar] [CrossRef] [PubMed]
- Jessberger, N.; Kranzler, M.; Da Riol, C.; Schwenk, V.; Buchacher, T.; Dietrich, R.; Ehling-Schulz, M.; Martlbauer, E. Assessing the toxic potential of enteropathogenic Bacillus cereus. Food Microbiol. 2019, 84, 103276. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Biological Hazards (BIOHAZ). Risks for public health related to the presence of Bacillus cereus and other Bacillus spp. including Bacillus thuringiensis in foodstuffs. EFSA J. 2016, 14, e04524. [Google Scholar] [CrossRef]
- Palma, L.; Munoz, D.; Berry, C.; Murillo, J.; Caballero, P. Bacillus thuringiensis toxins: An overview of their biocidal activity. Toxins 2014, 6, 3296–3325. [Google Scholar] [CrossRef] [PubMed]
- Lacey, L.A.; Grzywacz, D.; Shapiro-Ilan, D.I.; Frutos, R.; Brownbridge, M.; Goettel, M.S. Insect pathogens as biological control agents: Back to the future. J. Invertebr. Pathol. 2015, 132, 1–41. [Google Scholar] [CrossRef]
- Casida, J.E.; Bryant, R.J. The ABCs of pesticide toxicology: Amounts, biology, and chemistry. Toxicol. Res. 2017, 6, 755–763. [Google Scholar] [CrossRef]
- Jackson, S.G.; Goodbrand, R.B.; Ahmed, R.; Kasatiya, S. Bacillus cereus and Bacillus thuringiensis isolated in a gastroenteritis outbreak investigation. Lett. Appl. Microbiol. 1995, 21, 103–105. [Google Scholar] [CrossRef] [PubMed]
- Johler, S.; Kalbhenn, E.M.; Heini, N.; Brodmann, P.; Gautsch, S.; Bagcioglu, M.; Contzen, M.; Stephan, R.; Ehling-Schulz, M. Enterotoxin Production of Bacillus thuringiensis Isolates From Biopesticides, Foods, and Outbreaks. Front. Microbiol. 2018, 9, 1915. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, L.; Bernard, K.; Beniac, D.; Isaac-Renton, J.L.; Naseby, D.C. Identification of Bacillus cereus group species associated with food poisoning outbreaks in British Columbia, Canada. Appl. Environ. Microbiol. 2008, 74, 7451–7453. [Google Scholar] [CrossRef] [PubMed]
- Bonis, M.; Felten, A.; Pairaud, S.; Dijoux, A.; Maladen, V.; Mallet, L.; Radomski, N.; Duboisset, A.; Arar, C.; Sarda, X.; et al. Comparative phenotypic, genotypic and genomic analyses of Bacillus thuringiensis associated with foodborne outbreaks in France. PLoS ONE 2021, 16, e0246885. [Google Scholar] [CrossRef] [PubMed]
- Balls, M.; Parascandola, J. The Emergence and Early Fate of the Three Rs Concept. Altern. Lab. Anim. 2019, 47, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Office of Laboratory Animal Welfare, National Institutes of Health. Public Health Service Policy on Humane Care and Use of Laboratory Animals; Office of Laboratory Animal Welfare, National Institutes of Health: Bethesda, MD, USA, 2002.
- Fedi, A.; Vitale, C.; Ponschin, G.; Ayehunie, S.; Fato, M.; Scaglione, S. In vitro models replicating the human intestinal epithelium for absorption and metabolism studies: A systematic review. J. Control. Release 2021, 335, 247–268. [Google Scholar] [CrossRef] [PubMed]
- Andrews, C.; McLean, M.H.; Durum, S.K. Cytokine Tuning of Intestinal Epithelial Function. Front. Immunol. 2018, 9, 1270. [Google Scholar] [CrossRef] [PubMed]
- Cotton, J.A.; Platnich, J.M.; Muruve, D.A.; Jijon, H.B.; Buret, A.G.; Beck, P.L. Interleukin-8 in gastrointestinal inflammation and malignancy: Induction and clinical consequences. Int. J. Interferon Cytokine Mediat. Res. 2016, 8, 13–34. [Google Scholar] [CrossRef]
- Schwenk, V.; Riegg, J.; Lacroix, M.; Martlbauer, E.; Jessberger, N. Enteropathogenic Potential of Bacillus thuringiensis Isolates from Soil, Animals, Food and Biopesticides. Foods 2020, 9, 1484. [Google Scholar] [CrossRef] [PubMed]
- Buchon, N.; Osman, D. All for one and one for all: Regionalization of the Drosophila intestine. Insect Biochem. Mol. Biol. 2015, 67, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Younes, S.; Al-Sulaiti, A.; Nasser, E.A.A.; Najjar, H.; Kamareddine, L. Drosophila as a Model Organism in Host-Pathogen Interaction Studies. Front. Cell Infect. Microbiol. 2020, 10, 214. [Google Scholar] [CrossRef] [PubMed]
- Cadot, C.; Tran, S.L.; Vignaud, M.L.; De Buyser, M.L.; Kolsto, A.B.; Brisabois, A.; Nguyen-The, C.; Lereclus, D.; Guinebretiere, M.H.; Ramarao, N. InhA1, NprA, and HlyII as candidates for markers to differentiate pathogenic from nonpathogenic Bacillus cereus strains. J. Clin. Microbiol. 2010, 48, 1358–1365. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, M.; Foley, E. Microbial recognition regulates intestinal epithelial growth in homeostasis and disease. FEBS J. 2022, 289, 3666–3691. [Google Scholar] [CrossRef] [PubMed]
- Babin, A.; Nawrot-Esposito, M.P.; Gallet, A.; Gatti, J.L.; Poirie, M. Differential side-effects of Bacillus thuringiensis bioinsecticide on non-target Drosophila flies. Sci. Rep. 2020, 10, 16241. [Google Scholar] [CrossRef] [PubMed]
- Jneid, R.; Loudhaief, R.; Zucchini-Pascal, N.; Nawrot-Esposito, M.P.; Fichant, A.; Rousset, R.; Bonis, M.; Osman, D.; Gallet, A. Bacillus thuringiensis toxins divert progenitor cells toward enteroendocrine fate by decreasing cell adhesion with intestinal stem cells in Drosophila. eLife 2023, 12, e80179. [Google Scholar] [CrossRef] [PubMed]
- Fichant, A.; Felten, A.; Gallet, A.; Firmesse, O.; Bonis, M. Identification of Genetic Markers for the Detection of Bacillus thuringiensis Strains of Interest for Food Safety. Foods 2022, 11, 3924. [Google Scholar] [CrossRef] [PubMed]
- Criscuolo, A. A fast alignment-free bioinformatics procedure to infer accurate distance-based phylogenetic trees from genome assemblies. Res. Ideas Outcomes 2019, 5, e36178. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [PubMed]
- Lecadet, M.M.; Blondel, M.O.; Ribier, J. Generalized transduction in Bacillus thuringiensis var. berliner 1715 using bacteriophage CP-54Ber. J. Gen. Microbiol. 1980, 121, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Martins, R.R.; McCracken, A.W.; Simons, M.J.P.; Henriques, C.M.; Rera, M. How to Catch a Smurf?—Ageing and Beyond… In vivo Assessment of Intestinal Permeability in Multiple Model Organisms. Bio Protoc. 2018, 8, e2722. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, N.M.; Sestelo, M.; Meira-Machado, L.; Roca-Pardiñas, J. Clustcurv: An R Package for Determining Groups in Multiple Curves. R. J. 2021, 13, 164–183. [Google Scholar] [CrossRef]
- Buchacher, T.; Digruber, A.; Kanzler, M.; Del Favero, G.; Ehling-Schulz, M. Bacillus cereus extracellular vesicles act as shuttles for biologically active multicomponent enterotoxins. Cell Commun. Signal 2023, 21, 112. [Google Scholar] [CrossRef] [PubMed]
- Glasset, B.; Sperry, M.; Dervyn, R.; Herbin, S.; Brisabois, A.; Ramarao, N. The cytotoxic potential of Bacillus cereus strains of various origins. Food Microbiol. 2021, 98, 103759. [Google Scholar] [CrossRef] [PubMed]
- Jessberger, N.; Dietrich, R.; Bock, S.; Didier, A.; Martlbauer, E. Bacillus cereus enterotoxins act as major virulence factors and exhibit distinct cytotoxicity to different human cell lines. Toxicon 2014, 77, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Kone, K.M.; Hinnekens, P.; Jovanovic, J.; Rajkovic, A.; Mahillon, J. New Insights into the Potential Cytotoxic Role of Bacillus cytotoxicus Cytotoxin K-1. Toxins 2021, 13, 698. [Google Scholar] [CrossRef] [PubMed]
- Stenfors, L.P.; Mayr, R.; Scherer, S.; Granum, P.E. Pathogenic potential of fifty Bacillus weihenstephanensis strains. FEMS Microbiol. Lett. 2002, 215, 47–51. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Biological Hazards (BIOHAZ); Ricci, A.; Allende, A.; Bolton, D.; Chemaly, M.; Davies, R.; Girones, R.; Herman, L.; Koutsoumanis, K.; Lindqvist, R.; et al. Scientific Opinion on the update of the list of QPS-recommended biological agents intentionally added to food or feed as notified to EFSA. EFSA J. 2017, 15, e04664. [Google Scholar] [CrossRef]
- Castiaux, V.; Laloux, L.; Schneider, Y.-J.; Mahillon, J. Screening of Cytotoxic B. cereus on Differentiated Caco-2 Cells and in Co-Culture with Mucus-Secreting (HT29-MTX) Cells. Toxins 2016, 8, 320. [Google Scholar] [CrossRef] [PubMed]
- Jessberger, N.; Rademacher, C.; Krey, V.M.; Dietrich, R.; Mohr, A.K.; Bohm, M.E.; Scherer, S.; Ehling-Schulz, M.; Martlbauer, E. Simulating Intestinal Growth Conditions Enhances Toxin Production of Enteropathogenic Bacillus cereus. Front. Microbiol. 2017, 8, 627. [Google Scholar] [CrossRef] [PubMed]
- Hansen, B.M.; Thorsen, L.; Nielsen-LeRoux, C.; Wilcks, A.; Hendriksen, N.B. New experimental approaches for human risk assessment of microbial pest control agents—Exemplified by the bacterium Bacillus thuringiensis. Pestic. Res. 2011, 136, 1–128. [Google Scholar]
- Brasseur, K.; Auger, P.; Asselin, E.; Parent, S.; Cote, J.C.; Sirois, M. Parasporin-2 from a New Bacillus thuringiensis 4R2 Strain Induces Caspases Activation and Apoptosis in Human Cancer Cells. PLoS ONE 2015, 10, e0135106. [Google Scholar] [CrossRef] [PubMed]
- Ito, A.; Sasaguri, Y.; Kitada, S.; Kusaka, Y.; Kuwano, K.; Masutomi, K.; Mizuki, E.; Akao, T.; Ohba, M. A Bacillus thuringiensis crystal protein with selective cytocidal action to human cells. J. Biol. Chem. 2004, 279, 21282–21286. [Google Scholar] [CrossRef] [PubMed]
- Mendoza-Almanza, G.; Esparza-Ibarra, E.L.; Ayala-Lujan, J.L.; Mercado-Reyes, M.; Godina-Gonzalez, S.; Hernandez-Barrales, M.; Olmos-Soto, J. The Cytocidal Spectrum of Bacillus thuringiensis Toxins: From Insects to Human Cancer Cells. Toxins 2020, 12, 301. [Google Scholar] [CrossRef] [PubMed]
- Sonnier, D.I.; Bailey, S.R.; Schuster, R.M.; Lentsch, A.B.; Pritts, T.A. TNF-alpha induces vectorial secretion of IL-8 in Caco-2 cells. J. Gastrointest. Surg. 2010, 14, 1592–1599. [Google Scholar] [CrossRef] [PubMed]
- Codemo, M.; Muschiol, S.; Iovino, F.; Nannapaneni, P.; Plant, L.; Wai, S.N.; Henriques-Normark, B. Immunomodulatory Effects of Pneumococcal Extracellular Vesicles on Cellular and Humoral Host Defenses. mBio 2018, 9, e00559-18. [Google Scholar] [CrossRef] [PubMed]
- Tartaglia, N.R.; Breyne, K.; Meyer, E.; Cauty, C.; Jardin, J.; Chretien, D.; Dupont, A.; Demeyere, K.; Berkova, N.; Azevedo, V.; et al. Staphylococcus aureus Extracellular Vesicles Elicit an Immunostimulatory Response in vivo on the Murine Mammary Gland. Front. Cell Infect. Microbiol. 2018, 8, 277. [Google Scholar] [CrossRef]
- Jensen, G.B.; Larsen, P.; Jacobsen, B.L.; Madsen, B.; Smidt, L.; Andrup, L. Bacillus thuringiensis in fecal samples from greenhouse workers after exposure to B. thuringiensis-based pesticides. Appl. Environ. Microbiol. 2002, 68, 4900–4905. [Google Scholar] [CrossRef]
- Oliveira-Filho, E.C.; Oliveira, R.S.; Lopes, M.C.; Ramos, F.R.; Grisolia, C.K.; Alves, R.T.; Monnerat, R.G. Toxicity assessment and clearance of Brazilian microbial pest control agents in mice. Bull. Environ. Contam. Toxicol. 2009, 83, 570–574. [Google Scholar] [CrossRef]
- Wilcks, A.; Smidt, L.; Bahl, M.I.; Hansen, B.M.; Andrup, L.; Hendriksen, N.B.; Licht, T.R. Germination and conjugation of Bacillus thuringiensis subsp. israelensis in the intestine of gnotobiotic rats. J. Appl. Microbiol. 2008, 104, 1252–1259. [Google Scholar] [CrossRef] [PubMed]
- Loudhaief, R.; Brun-Barale, A.; Benguettat, O.; Nawrot-Esposito, M.P.; Pauron, D.; Amichot, M.; Gallet, A. Apoptosis restores cellular density by eliminating a physiologically or genetically induced excess of enterocytes in the Drosophila midgut. Development 2017, 144, 808–819. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.B.; Choi, O.K.; Kwon, S.M.; Cho, S.H.; Park, B.J.; Jin, N.Y.; Yu, Y.M.; Oh, D.H. Prevalence and Toxin Characteristics of Bacillus thuringiensis Isolated from Organic Vegetables. J. Microbiol. Biotechnol. 2017, 27, 1449–1456. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Hendriks, M.; Deleu, E.; Spanoghe, P.; Hofte, M.; van Overbeek, L.; Uyttendaele, M. Prevalence, attachment ability and strength of the biological control agent Bacillus thuringiensis on tomato. Food Microbiol. 2023, 112, 104235. [Google Scholar] [CrossRef] [PubMed]
- Ruder, B.; Atreya, R.; Becker, C. Tumour Necrosis Factor Alpha in Intestinal Homeostasis and Gut Related Diseases. Int. J. Mol. Sci. 2019, 20, 1887. [Google Scholar] [CrossRef] [PubMed]
- Panayidou, S.; Ioannidou, E.; Apidianakis, Y. Human pathogenic bacteria, fungi, and viruses in Drosophila: Disease modeling, lessons, and shortcomings. Virulence 2014, 5, 253–269. [Google Scholar] [CrossRef] [PubMed]
- Benguettat, O.; Jneid, R.; Soltys, J.; Loudhaief, R.; Brun-Barale, A.; Osman, D.; Gallet, A. The DH31/CGRP enteroendocrine peptide triggers intestinal contractions favoring the elimination of opportunistic bacteria. PLoS Pathog. 2018, 14, e1007279. [Google Scholar] [CrossRef] [PubMed]
- Broderick, N.A.; Lemaitre, B. Gut-associated microbes of Drosophila melanogaster. Gut Microbes 2012, 3, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Edgar, B.A. Insect Gut Regeneration. Cold Spring Harb. Perspect. Biol. 2022, 14, a040915. [Google Scholar] [CrossRef] [PubMed]
- Anastassiadou, M.; Arena, M.; Auteri, D.; Brancato, A.; Bura, L.; Carrasco Cabrera, L.; Chaideftou, E.; Chiusolo, A.; Crivellente, F.; De Lentdecker, C.; et al. Peer review of the pesticide risk assessment of the active substance Bacillus thuringiensis subsp. kurstaki strain SA-11. EFSA J. 2020, 18, e06261. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Global Dataset (n = 48) | Subdataset 1 (Caco-2 Cell Model) | Subdataset 2 (D. melanogaster Model) | |
|---|---|---|---|
| Bc non-Bt (n = 18) | 8 | 17 | |
| Bt (n = 27) | Bt pesticide (or mutant) | 11 | 10 |
| FBO-Bt “pesticide-like” * | 4 | 5 | |
| Bt non-pesticide (or mutant) | 11 | 4 | |
| Bacillus spp. non-Bc (n = 3) | 3 | 3 | |
| Total | 37 | 39 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fichant, A.; Lanceleur, R.; Hachfi, S.; Brun-Barale, A.; Blier, A.-L.; Firmesse, O.; Gallet, A.; Fessard, V.; Bonis, M. New Approach Methods to Assess the Enteropathogenic Potential of Strains of the Bacillus cereus Group, including Bacillus thuringiensis. Foods 2024, 13, 1140. https://doi.org/10.3390/foods13081140
Fichant A, Lanceleur R, Hachfi S, Brun-Barale A, Blier A-L, Firmesse O, Gallet A, Fessard V, Bonis M. New Approach Methods to Assess the Enteropathogenic Potential of Strains of the Bacillus cereus Group, including Bacillus thuringiensis. Foods. 2024; 13(8):1140. https://doi.org/10.3390/foods13081140
Chicago/Turabian StyleFichant, Arnaud, Rachelle Lanceleur, Salma Hachfi, Alexandra Brun-Barale, Anne-Louise Blier, Olivier Firmesse, Armel Gallet, Valérie Fessard, and Mathilde Bonis. 2024. "New Approach Methods to Assess the Enteropathogenic Potential of Strains of the Bacillus cereus Group, including Bacillus thuringiensis" Foods 13, no. 8: 1140. https://doi.org/10.3390/foods13081140
APA StyleFichant, A., Lanceleur, R., Hachfi, S., Brun-Barale, A., Blier, A. -L., Firmesse, O., Gallet, A., Fessard, V., & Bonis, M. (2024). New Approach Methods to Assess the Enteropathogenic Potential of Strains of the Bacillus cereus Group, including Bacillus thuringiensis. Foods, 13(8), 1140. https://doi.org/10.3390/foods13081140