Flavonoid-Rich Extract of Actinidia macrosperma (A Wild Kiwifruit) Inhibits Angiotensin-Converting Enzyme In Vitro



Abstract
:1. Introduction
2. Methodology
2.1. Chemicals
2.2. Plant Materials
2.3. Preparation of Flavonoid-Rich Kiwifruit Extracts
2.4. UPLC-MS/MS Analysis of Phenolics in the Kiwifruit Extracts
2.5. Assay for ACE Inhibitory Activity
2.6. Determination of Kinetic Parameters of ACE Inhibition
2.7. Statistical Analysis
3. Results and Discussion
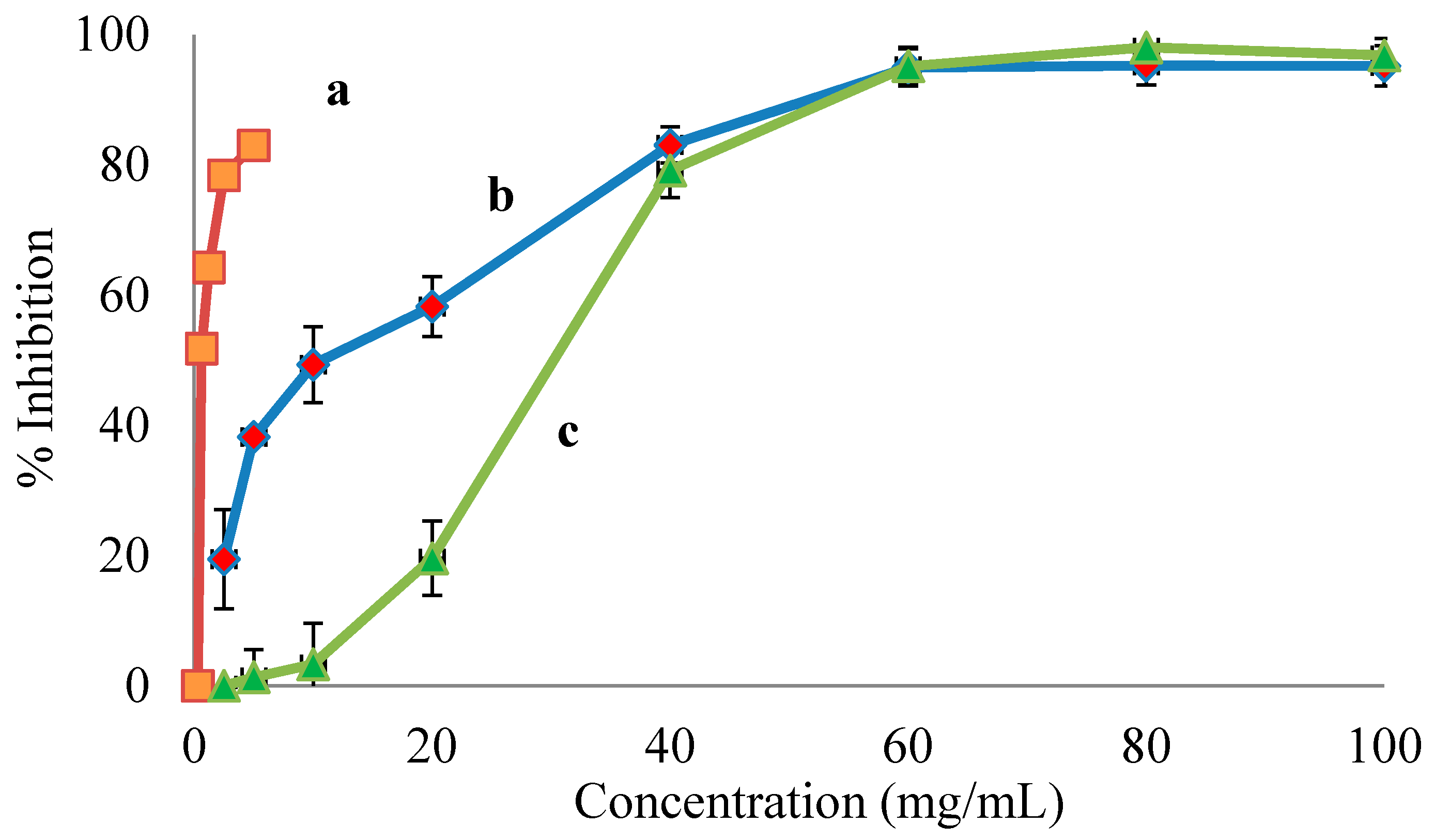
3.1. ACE Inhibition
3.2. Determination of Kinetic Parameters of ACE Inhibition
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Dojki, F.K.; Bakris, G.L. Blood Pressure Control and Cardiovascular/Renal Outcomes. Endocrinol. Metab. Clin. N. Am. 2018, 47, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Atlas, S.A. The renin-angiotensin aldosterone system: Pathophysiological role and pharmacologic inhibition. J. Manag. Care Pharm. 2007, 13, S9–S20. [Google Scholar] [CrossRef] [PubMed]
- Balasuriya, B.W.N.; Rupasinghe, H.P.V. Plant flavonoids as angiotensin-converting enzyme inhibitors in regulation of hypertension. Funct. Foods Health Dis. 2011, 5, 172–188. [Google Scholar]
- Quan, A. Fetopathy associated with exposure to angiotensin-converting enzyme inhibitors and angiotensin receptor antagonists. Early Hum. Dev. 2006, 82, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.; Kang, D.G.; Kwon, J.W.; Kwon, T.O.; Lee, S.Y.; Lee, D.B.; Lee, H.S. Isolation of angiotensin-converting enzyme (ACE) inhibitory flavonoids from Sedum sarmentosum. Biol. Pharm. Bull. 2004, 27, 2035–2037. [Google Scholar] [CrossRef] [PubMed]
- Actis-Goretta, L.; Ottaviani, J.I.; Fraga, C.G. Inhibition of angiotensin-converting enzyme activity by flavanol-rich foods. J. Agric. Food Chem. 2006, 54, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Ottaviani, J.I.; Actis-Goretta, L.; Villordo, J.J.; Fraga, C.G. Procyanidin structure defines the extent and specificity of angiotensin I converting enzyme inhibition. Biochimie 2006, 88, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Braga, F.C.; Serra, C.P.; Viana Júnior, N.S.; Oliveira, A.B.; Côrtes, S.F.; Lombardi, J.A. Angiotensin-converting enzyme inhibition by Brazilian plants. Fitoterapia 2007, 78, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Cinq-Mars, C.D.; Li-Chan, E.C.Y. Optimizing angiotensin I-converting enzyme inhibitory activity of Pacific hake (Merluccius productus) fillet hydrolysate using response surface methodology and ultrafiltration. J. Agric. Food Chem. 2007, 55, 9380–9388. [Google Scholar] [CrossRef] [PubMed]
- Loizzo, M.R.; Said, A.; Tundis, R.; Rashed, K.; Statti, G.A.; Hufner, A.; Menichini, F. Inhibition of angiotensin-converting enzyme (ACE) by flavonoids isolated from Ailanthus excelsa (Roxb) (simaroubaceae). Phytother. Res. 2007, 21, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Farzamirad, V.; Aluko, R.E. Angiotensin-converting enzyme inhibition and free-radical scavenging properties of cationic peptides derived from soybean protein hydrolysates. Int. J. Food Sci. Nutr. 2008, 59, 428–437. [Google Scholar] [CrossRef] [PubMed]
- Hong, F.; Ming, L.; Yi, S.; Zhanxia, L.; Yongquan, W.; Chi, L. The antihypertensive effect of peptides: A novel alternative to drugs? Peptides 2008, 29, 1062–1071. [Google Scholar] [CrossRef] [PubMed]
- Guang, C.; Phillips, R.D. Plant food-derived angiotensin I converting enzyme inhibitory peptides. J. Agric. Food Chem. 2009, 57, 5113–5120. [Google Scholar] [CrossRef] [PubMed]
- Udenigwe, C.C.; Lin, Y.S.; Hou, W.C.; Aluko, R.E. Kinetics of the inhibition of renin and angiotensin I-converting enzyme by flaxseed protein hydrolysate fractions. J. Funct. Foods 2009, 1, 199–207. [Google Scholar] [CrossRef]
- Balasuriya, N.; Rupasinghe, H.P.V. Antihypertensive properties of flavonoid-rich apple peel extract. Food Chem. 2012, 135, 2320–2325. [Google Scholar] [CrossRef] [PubMed]
- Aluko, R.E. Structure and function of plant protein-derived antihypertensive peptides. Curr. Opin. Food Sci. 2015, 4, 44–50. [Google Scholar] [CrossRef]
- Hodgson, J.M.; Devine, A.; Puddey, I.B.; Chan, S.Y.; Beilin, L.J.; Prince, R.L. Tea intake is inversely related to blood pressure in older women. J. Nutr. 2003, 133, 2883–2886. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, F.; Siddesha, J.M.; Urooj, A.; Vishwanath, B.S. Radical scavenging and angiotensin-converting enzyme inhibitory activities of standardized extracts of Ficus racemosa stem bark. Phytother. Res. 2010, 24, 1839–1843. [Google Scholar] [CrossRef] [PubMed]
- Lecour, S.; Lamont, K.T. Natural polyphenols and cardioprotection. Mini-Rev. Med. Chem. 2011, 11, 1191–1199. [Google Scholar] [PubMed]
- Balasuriya, N.; Rupasinghe, H.P.V.; Sweeney, M.; McCarron, S.; Gottschall-Pass, K. Antihypertensive effects of apple peel extract on spontaneously hypertensive rats. Pharmacologia 2015, 6, 371–376. [Google Scholar]
- Ferguson, A.R. The need for characterization and evaluation of germplasm: Kiwifruit as an example. Euphytica 2007, 154, 371–382. [Google Scholar] [CrossRef]
- Nishiyama, I. Fruits of the Actinidia Genus. Adv. Food Nutr. Res. 2007, 52, 293–324. [Google Scholar] [PubMed]
- Fiorentino, A.; D’Abrosca, B.; Pacifico, S.; Mastellone, C.; Scognamiglio, M.; Monaco, P. Identification and assessment of antioxidant capacity of phytochemicals from kiwi fruits. J. Agric. Food Chem. 2009, 57, 4148–4155. [Google Scholar] [CrossRef] [PubMed]
- Hunter, D.C.; Greenwood, J.; Zhang, J.; Skinner, M.A. Antioxidant and ‘natural protective’ properties of kiwifruit. Curr. Top. Med. Chem. 2011, 11, 1811–1820. [Google Scholar] [CrossRef] [PubMed]
- Stonehouse, W.; Gammon, C.S.; Beck, K.L.; Conlon, C.A.; von Hurst, P.R.; Kruger, R. Kiwifruit: Our daily prescription for health. Can. J. Physiol. Pharmacol. 2012, 91, 442–447. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zhao, Y.; Fu, C. Biological activities of extracts from a naturally wild kiwifruit, Actinidia macrosperma. Afr. J. Agric. Res. 2011, 6, 2231–2234. [Google Scholar]
- Rupasinghe, H.P.V.; Wang, L.; Huber, G.M.; Pitts, N.L. Effect of baking on dietary fibre and phenolics of muffins incorporated with apple skin powder. Food Chem. 2008, 107, 1217–1224. [Google Scholar] [CrossRef]
- Lu, Y.; Zhao, Y.; Fu, C. Preliminary evaluation of antimicrobial and cytotoxic activities of extracts from Actinidia macrosperma. Adv. Mater. Res. 2012, 455, 1200–1203. [Google Scholar] [CrossRef]
- Latocha, P.; Krupa, T.; Wołosiak, R.; Worobiej, E.; Wilczak, J. Antioxidant activity and chemical difference in fruit of different Actinidia sp. Int. J. Food Sci. Nutr. 2010, 61, 381–394. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Fan, J.; Zhao, Y.; Chen, S.; Zheng, X.; Yin, Y.; Fu, C. Immunomodulatory activity of aqueous extract of Actinidia macrosperma. Asia Pac. J. Clin. Nutr. 2007, 16 (Suppl. 1), 261–265. [Google Scholar]
- Actis-Goretta, L.; Ottaviani, J.I.; Keen, C.L.; Fraga, C.G. Inhibition of angiotensin-converting enzyme (ACE) activity by flavan-3-ols and procyanidins. FEBS Lett. 2003, 555, 597–600. [Google Scholar] [CrossRef]
- Persson, I.A.L.; Josefsson, M.; Persson, K.; Andersson, R.G. Tea flavanols inhibit angiotensin-converting enzyme activity and increase nitric oxide production in human endothelial cells. J. Pharm. Pharmacol. 2006, 58, 1139–1144. [Google Scholar] [CrossRef] [PubMed]
- Pinto, M.D.S.; Kwon, Y.I.; Apostolidis, E.; Lajolo, F.M.; Genovese, M.I.; Shetty, K. Potential of Ginkgo biloba L. leaves in the management of hyperglycemia and hypertension using in vitro models. Bioresour. Technol. 2009, 100, 6599–6609. [Google Scholar] [CrossRef] [PubMed]
- Ojeda, D.; Jiménez-Ferrer, E.; Zamilpa, A.; Herrera-Arellano, A.; Tortoriello, J.; Alvarez, L. Inhibition of angiotensin converting enzyme (ACE) activity by the anthocyanins delphinidin- and cyanidin-3-O-sambubiosides from Hibiscus sabdariffa. J. Ethnopharmacol. 2010, 127, 7–10. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Kiwifruit Genotypes | IC50 (mg/mL) |
|---|---|
| A.macrosperma | 0.49 |
| A.chinesis cv Hort 16A | 12.81 |
| A.deliciosa cv Hayward | 30.49 |
| Group/Name of the Flavonoid | Concentration of Phenolics (µg/g DW) | ||
|---|---|---|---|
| A. macrosperma | A. chinensis | A. deliciosa | |
| Flavonol | |||
| Quercetin-3-O-Galactoside | 470.9 | 205.19 | 441.39 |
| Quercetin-3-O-Glucoside | 4.16 | 0.45 | 0.14 |
| Quercetin Arabinoglucoside | 2.53 | 0.06 | 0.21 |
| Quercetin-3-O-Rhamnoside | 2.99 | 0.61 | 0.31 |
| Quercetin | 2.56 | nd | 0.17 |
| Quercetin-3-O-Rutinoside | 1.96 | 0.29 | 0.55 |
| Flavanol | |||
| Epigallocatechin | 1.55 | 0.61 | 0.46 |
| Catechin | 54.31 | 0.75 | 0.30 |
| Epicatechin | 0.91 | 5.15 | 0.74 |
| Epigallocatechingallate | 0.75 | nd | 0.40 |
| Dihydrochalcones | |||
| Phloridzin | 3.12 | 2.03 | 5.08 |
| Phloritin | 0.14 | 0.21 | 0.14 |
| Phenolic acids | |||
| Chlorogenic acid | 1.97 | 0.39 | 0.28 |
| Caffeic acid | 1.64 | 0.04 | 0.08 |
| Ferulic acid | 4.70 | 0.42 | 0.60 |
| Isoferulic acid | 32.71 | 15.12 | 28.17 |
| Total Phenolics | 586.9 | 231.32 | 479.02 |
| Extract | Concentration Tested (mg/mL) | Km (mM) | Vmax (mM/min) | Ki (mg/mL) |
|---|---|---|---|---|
| No inhibitor | 0 | 0.074 | 0.024 | |
| A. chinesis cv Hort 16A | 13.00 | 2.036 | 0.033 | 44.516 |
| A. deliciosa cv Hayward | 31.00 | 4.849 | 0.033 | 64.041 |
| A. macrosperma | 2.60 | 7.258 | 0.069 | 78.312 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hettihewa, S.K.; Hemar, Y.; Rupasinghe, H.P.V. Flavonoid-Rich Extract of Actinidia macrosperma (A Wild Kiwifruit) Inhibits Angiotensin-Converting Enzyme In Vitro. Foods 2018, 7, 146. https://doi.org/10.3390/foods7090146
Hettihewa SK, Hemar Y, Rupasinghe HPV. Flavonoid-Rich Extract of Actinidia macrosperma (A Wild Kiwifruit) Inhibits Angiotensin-Converting Enzyme In Vitro. Foods. 2018; 7(9):146. https://doi.org/10.3390/foods7090146
Chicago/Turabian StyleHettihewa, Sujeewa K., Yacine Hemar, and H. P. Vasantha Rupasinghe. 2018. "Flavonoid-Rich Extract of Actinidia macrosperma (A Wild Kiwifruit) Inhibits Angiotensin-Converting Enzyme In Vitro" Foods 7, no. 9: 146. https://doi.org/10.3390/foods7090146
APA StyleHettihewa, S. K., Hemar, Y., & Rupasinghe, H. P. V. (2018). Flavonoid-Rich Extract of Actinidia macrosperma (A Wild Kiwifruit) Inhibits Angiotensin-Converting Enzyme In Vitro. Foods, 7(9), 146. https://doi.org/10.3390/foods7090146