Use of Selected Lactobacilli to Increase γ-Aminobutyric Acid (GABA) Content in Sourdough Bread Enriched with Amaranth Flour



Abstract
:1. Introduction
2. Materials and Methods
2.1. Lactobacilli Strains and Culture Conditions
2.2. Liquid Sourdough Fermentations
2.3. Sourdough Fermentation and Bread Making
2.4. Monitoring of Selected Lactobacilli Strains in Sourdough Fermentations
2.5. Determination of pH, Total Titratable Acidity, Volume Increase, and Enumeration of Cultivable Bacteria and Yeasts
2.6. Determination of Lactic Acid by HPLC
2.7. Total Phenols Assay by Folin–Ciocalteau Reagent
2.8. Antioxidant Capacity by DPPH Assay
2.9. HPLC Determination of GABA Content
2.10. Sensory Evaluation
2.11. Statistical Analysis
3. Results and Discussion
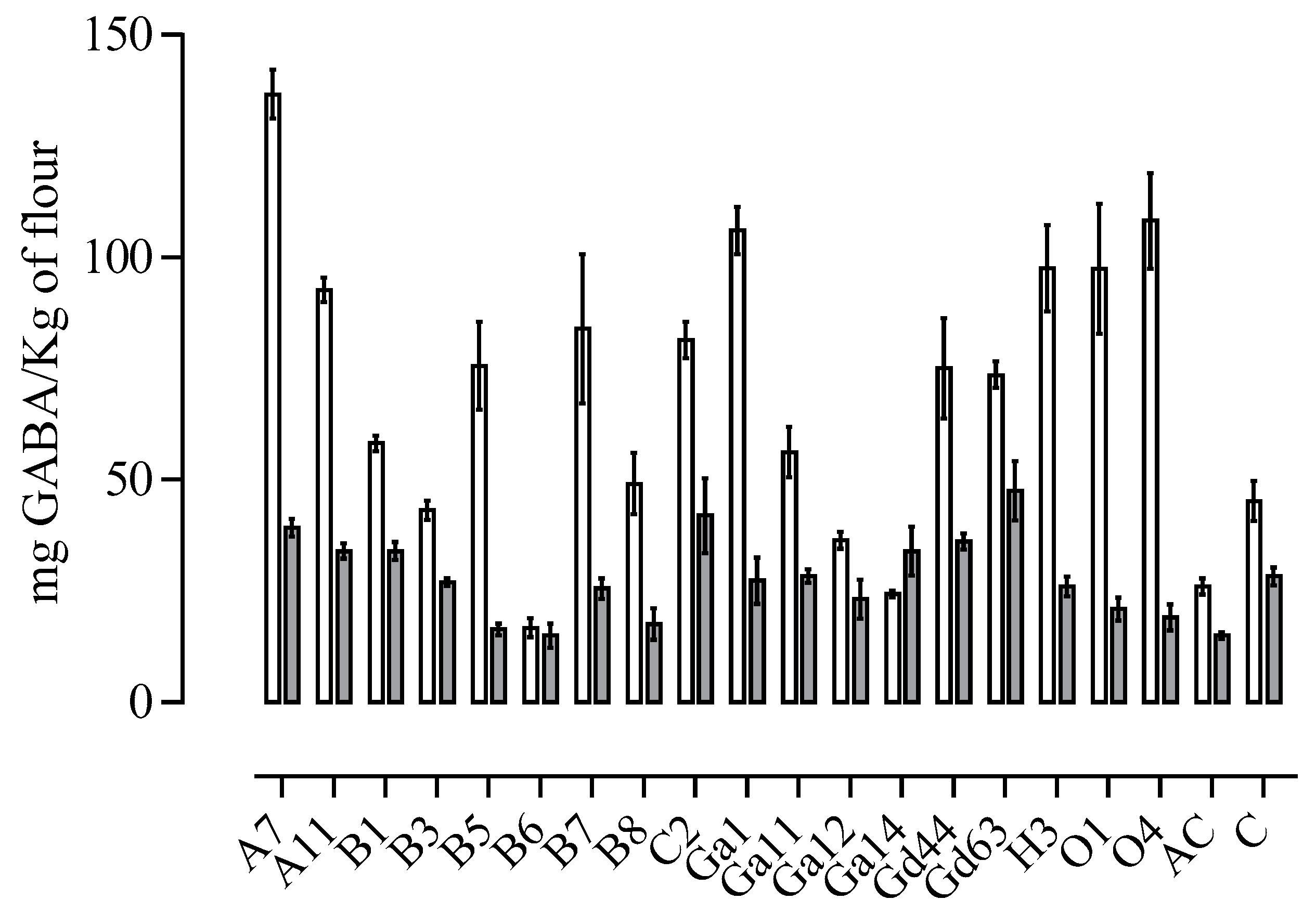
3.1. Selection of GABA-Producing Lactobacilli Strains
3.2. Sourdough Bread Fermentation with Selected Lactobacilli Strains
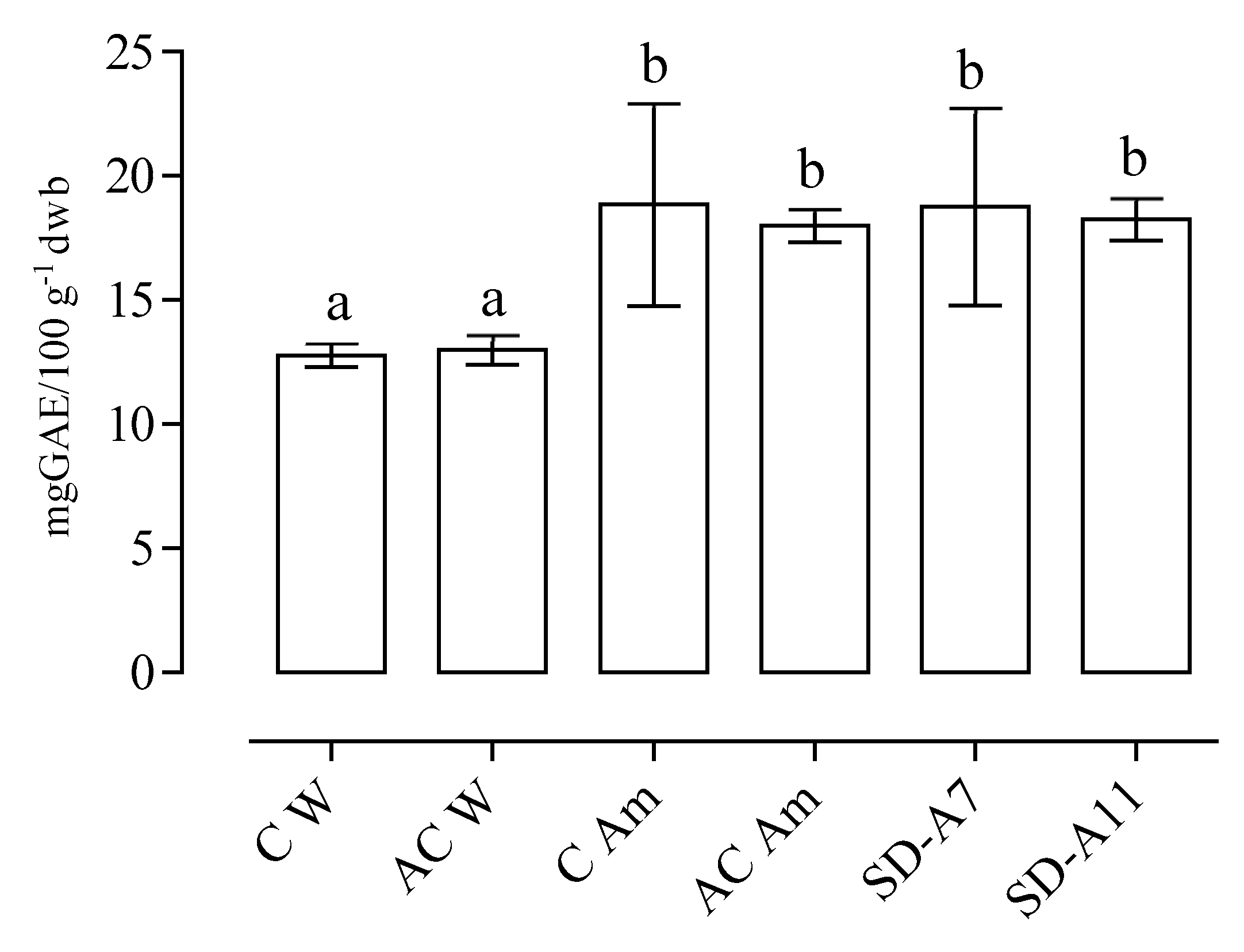
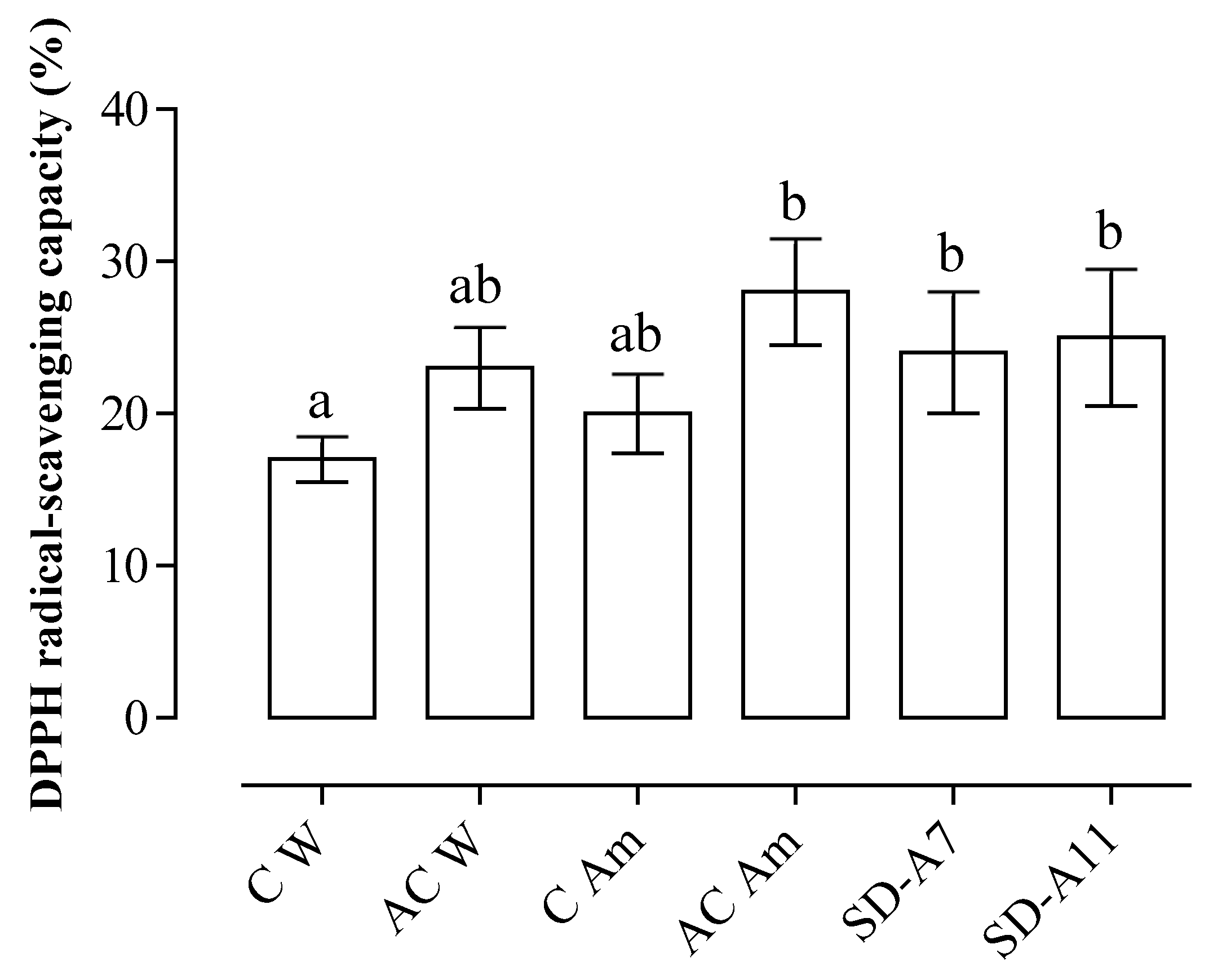
3.3. Phenolic Content and Antioxidant Capacity of Breads
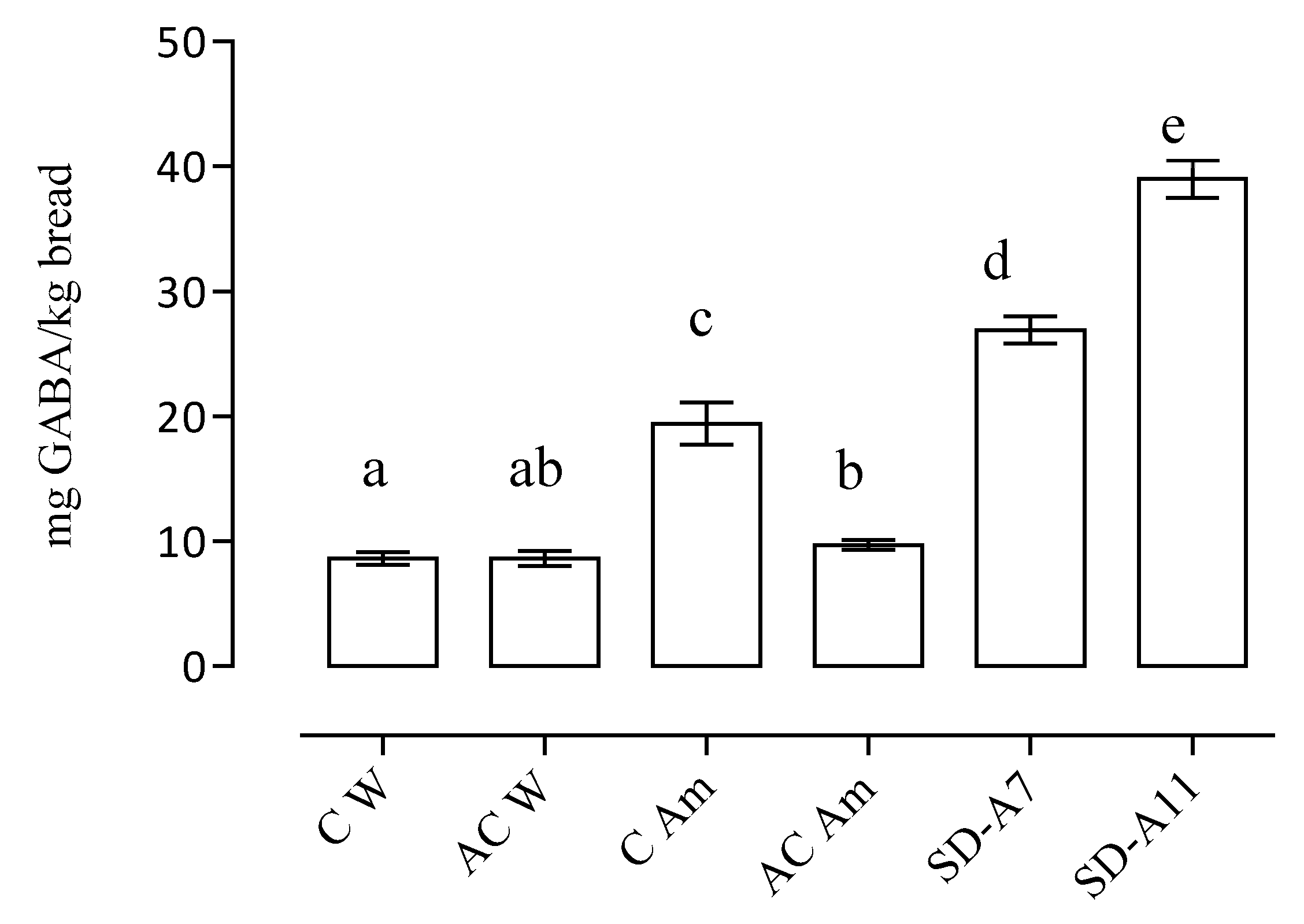
3.4. GABA Content of Breads
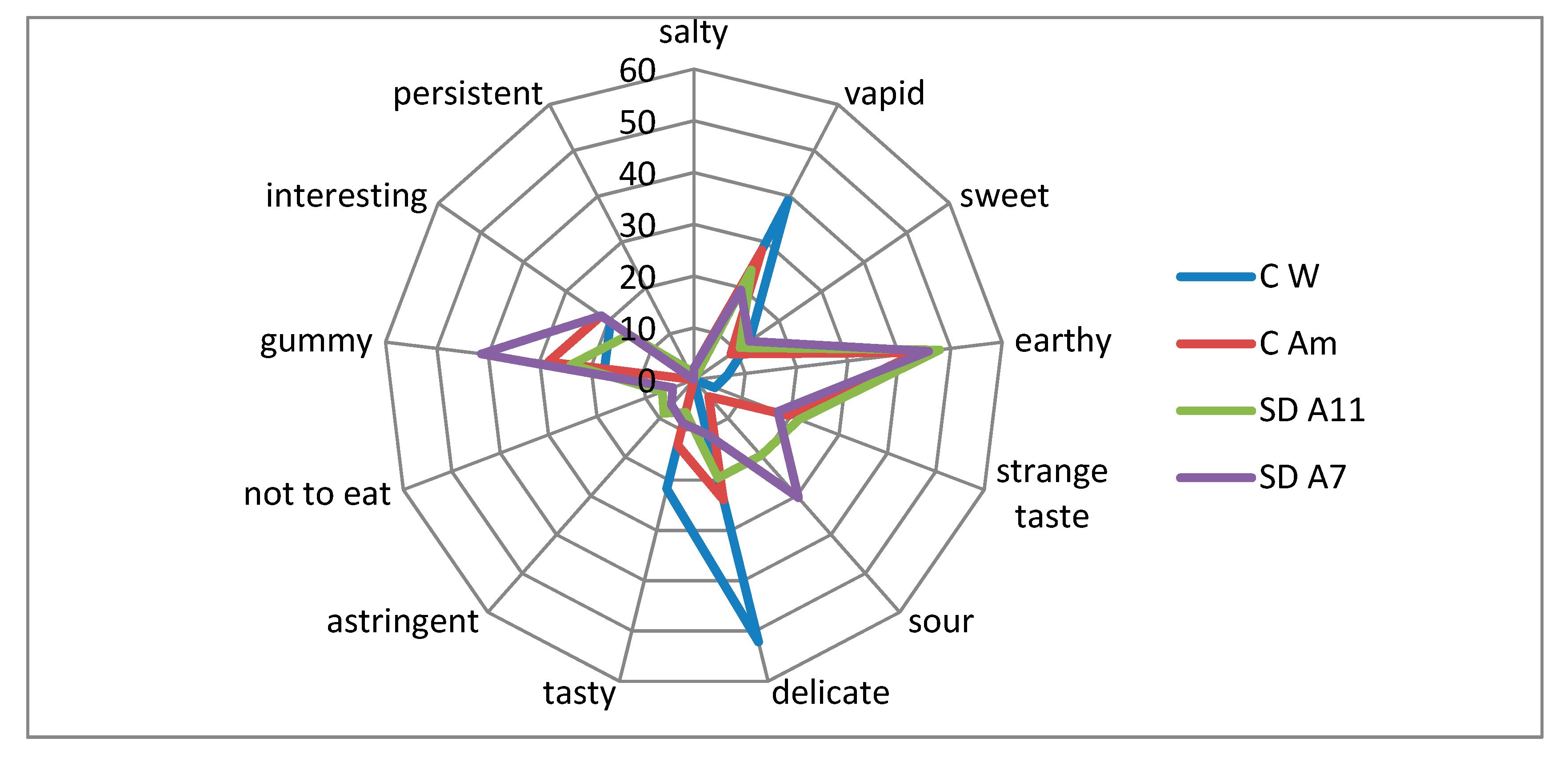
3.5. Sensory Evaluation
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chavan, R.S.; Chavan, S.R. Sourdough technology—A traditional way for wholesome foods: A review. Compr. Rev. Food Sci. Food Saf. 2011, 10, 169–182. [Google Scholar] [CrossRef]
- Arendt, E.K.; Ryan, L.A.; Dal Bello, F. Impact of sourdough on the texture of bread. Food Microbiol. 2007, 24, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Hammes, W.P.; Gänzle, M.G. Sourdough breads and related products. In Microbiology of Fermented Foods, 2nd ed.; Woods, B.J.B., Ed.; Blackie Academic: London, UK, 1998; pp. 199–216. [Google Scholar]
- Gobbetti, M.; De Angelis, M.; Di Cagno, R.; Calasso, M.; Archetti, G.; Rizzello, C.G. Novel insights on the functional/nutritional features of the sourdough fermentation. Int. J. Food Microbiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Diana, M.; Quílez, J.; Rafecas, M. Gamma-aminobutyric acid as a bioactive compound in foods: A review. J. Funct. Foods 2014, 10, 407–420. [Google Scholar] [CrossRef]
- Di Cagno, R.; De Angelis, M.; Lavermicocca, P.; De Vincenzi, M.; Giovannini, C.; Faccia, M.; Gobbetti, M. Proteolysis by sourdough lactic acid bacteria: Effects on wheat flour protein fractions and gliadin peptides involved in human cereal intolerance. Appl. Environ. Microbiol. 2002, 68, 623–633. [Google Scholar] [CrossRef]
- Gänzle, M.G.; Loponen, J.; Gobbetti, M. Proteolysis in sourdough fermentations: Mechanisms and potential for improved bread quality. Trends Food Sci. Technol. 2008, 19, 513–521. [Google Scholar] [CrossRef]
- Gobbetti, M.; Rizzello, C.G.; Di Cagno, R.; De Angelis, M. How the sourdough may affect the functional features of leavened baked goods. Food Microbiol. 2014, 37, 30–40. [Google Scholar] [CrossRef]
- Paterson, A.; Piggott, J.R. Flavour in sourdough breads: A review. Trends Food Sci. Technol. 2006, 17, 557–566. [Google Scholar] [CrossRef]
- Bhanwar, S.; Bamnia, M.; Ghosh, M.; Ganguli, A. Use of Lactococcus lactis to enrich sourdough bread with γ-aminobutyric acid. Int. J. Food Sci. Nutr. 2013, 64, 77–81. [Google Scholar] [CrossRef]
- Diana, M.; Rafecas, M.; Quílez, J. Free amino acids, acrylamide and biogenic amines in gamma-aminobutyric acid enriched sourdough and commercial breads. J. Cereal Sci. 2014, 60, 639–644. [Google Scholar] [CrossRef]
- Huang, J.; Mei, L.H.; Wu, H.; Lin, D.Q. Biosynthesis of γ-aminobutyric acid (GABA) using immobilized whole cells of Lactobacillus brevis. World J. Microbiol. Biot. 2007, 23, 865–871. [Google Scholar] [CrossRef]
- Komatsuzaki, N.; Shima, J.; Kawamoto, S.; Momose, H.; Kimura, T. Production of γ-aminobutyric acid (GABA) by Lactobacillus paracasei isolated from traditional fermented foods. Food Microbiol. 2005, 22, 497–504. [Google Scholar] [CrossRef]
- Komatsuzaki, N.; Tsukahara, K.; Toyoshima, H.; Suzuki, T.; Shimizu, N.; Kimura, T. Effect of soaking and gaseous treatment on GABA content in germinated brown rice. J. Food Eng. 2007, 78, 556–560. [Google Scholar] [CrossRef]
- Su, Y.C.; Wang, J.J.; Lin, T.T.; Pan, T.M. Production of the secondary metabolites γ-aminobutyric acid and monacolin K by Monascus. J. Ind. Microbiol. Biot. 2003, 30, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Okada, T.; Sugishita, T.; Murakami, T.; Murai, H.; Saikusa, T.; Horino, T.; Onoda, A.; Kajimoto, O.; Takahashi, R.; Takahashi, T. Effect of the defatted rice germ enriched with GABA for sleeplessness, depression, autonomic disorder by oral administration. J. Jpn. Soc. Food Sci. 2000, 47, 596–603. [Google Scholar] [CrossRef]
- Ueno, H. Enzymatic and structural aspects on glutamate decarboxylase. J. Mol. Catal. B-Enzym. 2000, 10, 67–79. [Google Scholar] [CrossRef]
- Coda, R.; Rizzello, C.G.; Gobbetti, M. Use of sourdough fermentation and pseudo-cereals and leguminous flours for the making of a functional bread enriched of γ-aminobutyric acid (GABA). Int. J. Food Microbiol. 2010, 137, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Rizzello, C.G.; Cassone, A.; Di Cagno, R.; Gobbetti, M. Synthesis of angiotensin I-converting enzyme (ACE)-inhibitory peptides and γ-aminobutyric acid (GABA) during sourdough fermentation by selected lactic acid bacteria. J. Agric. Food Chem. 2008, 56, 6936–6943. [Google Scholar] [CrossRef]
- Villegas, J.M.; Brown, L.; de Giori, G.S.; Hebert, E.M. Optimization of batch culture conditions for GABA production by Lactobacillus brevis CRL 1942, isolated from quinoa sourdough. LWT-Food Sci. Technol. 2016, 67, 22–26. [Google Scholar] [CrossRef]
- Lamberts, L.; Joye, I.J.; Beliën, T.; Delcour, J.A. Dynamics of γ-aminobutyric acid in wheat flour bread making. Food Chem. 2012, 130, 896–901. [Google Scholar] [CrossRef]
- Coda, R.; Di Cagno, R.; Gobbetti, M.; Rizzello, C.G. Sourdough lactic acid bacteria: Exploration of non-wheat cereal-based fermentation. Food Microbiol. 2014, 37, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Schoenlechner, R.; Siebenhandl, S.; Berghofer, E. Pseudocereals. In Gluten-Free Cereal Products and Beverages, 1st ed.; Arendt, E.K., Dal Bello, F., Eds.; Academic Press: New York, NY, USA, 2008; p. 149-VI. [Google Scholar]
- Chlopicka, J.; Pasko, P.; Gorinstein, S.; Jedryas, A.; Zagrodzki, P. Total phenolic and total flavonoid content, antioxidant activity and sensory evaluation of pseudocereal breads. LWT-Food Sci. Technol. 2012, 46, 548–555. [Google Scholar] [CrossRef]
- Gorinstein, S.; Vargas, O.J.M.; Jaramillo, N.O.; Salas, I.A.; Ayala, A.L.M.; Arancibia-Avila, P.; Toledo, F.; Katrich, E.; Trakhtenberg, S. The total polyphenols and the antioxidant potentials of some selected cereals and pseudocereals. Eur. Food Res. Technol. 2007, 225, 321–328. [Google Scholar] [CrossRef]
- Kalinova, J.; Dadakova, E. Rutin and total quercetin content in amaranth (Amaranthus spp.). Plant. Food. Hum. Nutr. 2009, 64, 68. [Google Scholar] [CrossRef]
- Li, S.Q.; Zhang, Q.H. Advances in the development of functional foods from buckwheat. Crit. Rev. Food Sci. 2001, 41, 451–464. [Google Scholar] [CrossRef]
- Lin, L.Y.; Liu, H.M.; Yu, Y.W.; Lin, S.D.; Mau, J.L. Quality and antioxidant property of buckwheat enhanced wheat bread. Food Chem. 2009, 112, 987–991. [Google Scholar] [CrossRef]
- Paśko, P.; Sajewicz, M.; Gorinstein, S.; Zachwieja, Z. Analysis of selected phenolic acids and flavonoids in Amaranthus cruentus and Chenopodium quinoa seeds and sprouts by HPLC. Acta Chromatogr. 2008, 20, 661–672. [Google Scholar] [CrossRef]
- Richardson, D.P. Wholegrain health claims in Europe. Proc. Nutr. Soc. 2003, 62, 161–169. [Google Scholar] [CrossRef] [Green Version]
- Nimbalkar, M.S.; Pai, S.R.; Pawar, N.V.; Oulkar, D.; Dixit, G.B. Free amino acid profiling in grain Amaranth using LC–MS/MS. Food Chem. 2012, 134, 2565–2569. [Google Scholar] [CrossRef]
- Jekle, M.; Houben, A.; Mitzscherling, M.; Becker, T. Effects of selected lactic acid bacteria on the characteristics of amaranth sourdough. J. Sci. Food Agric. 2010, 90, 2326–2332. [Google Scholar] [CrossRef]
- Galli, V.; Venturi, M.; Pini, N.; Guerrini, S.; Granchi, L.; Vincenzini, M. Liquid and firm sourdough fermentation: Microbial robustness and interactions during consecutive backsloppings. LWT-Food Sci. Technol. 2019, 105, 9–15. [Google Scholar] [CrossRef]
- Venturi, M.; Guerrini, S.; Granchi, L.; Vincenzini, M. Typing of Lactobacillus sanfranciscensis isolates from traditional sourdoughs by combining conventional and multiplex RAPD–PCR profiles. Int. J. Food Microbiol. 2012, 156, 122–126. [Google Scholar] [CrossRef] [PubMed]
- Seseña, S.; Sánchez, I.; Palop, L. Characterization of Lactobacillus strains and monitoring by RAPD-PCR in controlled fermentations of “Almagro” eggplants. Int. J. Food Microbiol. 2005, 104, 325–335. [Google Scholar] [CrossRef]
- Reguant, C.; Bordons, A. Typification of Oenococcus oeni strains by multiplex RAPD-PCR and study of population dynamics during malolactic fermentation. J. Appl. Microbiol. 2003, 95, 344–353. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Jubete, L.; Wijngaard, H.; Arendt, E.K.; Gallagher, E. Polyphenol composition and in vitro antioxidant activity of amaranth, quinoa buckwheat and wheat as affected by sprouting and baking. Food Chem. 2010, 119, 770–778. [Google Scholar] [CrossRef]
- Tuberoso, C.I.G.; Congiu, F.; Serreli, G.; Mameli, S. Determination of dansylated amino acids and biogenic amines in Cannonau and Vermentino wines by HPLC-FLD. Food Chem. 2015, 175, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Corona, O.; Alfonzo, A.; Ventimiglia, G.; Nasca, A.; Francesca, N.; Martorana, A.; Moschetti, G.; Settanni, L. Industrial application of selected lactic acid bacteria isolated from local semolinas for typical sourdough bread production. Food Microbial. 2016, 59, 43–56. [Google Scholar] [CrossRef] [Green Version]
- Siragusa, S.; De Angelis, M.; Di Cagno, R.; Rizzello, C.G.; Coda, R.; Gobbetti, M. Synthesis of γ-aminobutyric acid by lactic acid bacteria isolated from a variety of Italian cheeses. Appl. Environ. Microbiol. 2007, 73, 7283–7290. [Google Scholar] [CrossRef]
- Ayo, J.A. The effect of amaranth grain flour on the quality of bread. Int. J. Food Prop. 2001, 4, 341–351. [Google Scholar] [CrossRef]
- Sanz-Penella, J.M.; Wronkowska, M.; Soral-Smietana, M.; Haros, M. Effect of whole amaranth flour on bread properties and nutritive value. LWT-Food Sci. Technol. 2013, 50, 679–685. [Google Scholar] [CrossRef] [Green Version]
- Sindhuja, A.; Sudha, M.L.; Rahim, A. Effect of incorporation of amaranth flour on the quality of cookies. Eur. Food Res. Technol. 2005, 221, 597. [Google Scholar] [CrossRef]
- Minervini, F.; Lattanzi, A.; De Angelis, M.; Di Cagno, R.; Gobbetti, M. Influence of artisan bakery-or laboratory-propagated sourdoughs on the diversity of lactic acid bacterium and yeast microbiotas. Appl. Environ. Microbiol. 2012, 78, 5328–5340. [Google Scholar] [CrossRef] [PubMed]
- Ventimiglia, G.; Alfonzo, A.; Galluzzo, P.; Corona, O.; Francesca, N.; Caracappa, S.; Moschetti, G.; Settanni, L. Codominance of Lactobacillus plantarum and obligate heterofermentative lactic acid bacteria during sourdough fermentation. Food Microbiol. 2015, 51, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Lattanzi, A.; Minervini, F.; Di Cagno, R.; Diviccaro, A.; Antonielli, L.; Cardinali, G.; Cappelle, S.; De Angelis, M.; Gobbetti, M. The lactic acid bacteria and yeast microbiota of eighteen sourdoughs used for the manufacture of traditional Italian sweet leavened baked goods. Int. J. Food Microbiol. 2013, 163, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Vollmannova, A.; Margitanova, E.; Tóth, T.; Timoracka, M.; Urminska, D.; Bojňanská, T.; Čičová, I. Cultivar influence on total polyphenol and rutin contents and total antioxidant capacity in buckwheat, amaranth, and quinoa seeds. Czech J. Food Sci. 2013, 31, 589–595. [Google Scholar] [CrossRef] [Green Version]
- Liukkonen, K.H.; Katina, K.; Wilhelmsson, A.; Myllymaki, O.; Lampi, A.M.; Kariluoto, S.; Piironen, V.; Heinonen, S.M.; Nurmi, T.; Adlercreutz, H.; et al. Process-induced changes on bioactive compounds in whole grain rye. Proc. Nutr. Soc. 2003, 62, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Poutanen, K.; Flander, L.; Katina, K. Sourdough and cereal fermentation in a nutritional perspective. Food Microbiol. 2009, 26, 693–699. [Google Scholar] [CrossRef]
- Park, K.B.; Oh, S.H. Production of yogurt with enhanced levels of gamma-aminobutyric acid and valuable nutrients using lactic acid bacteria and germinated soybean extract. Bioresour. Technol. 2007, 98, 1675–1679. [Google Scholar] [CrossRef]
- Kim, J.Y.; Lee, M.Y.; Ji, G.E.; Lee, Y.S.; Hwang, K.T. Production of γ-aminobutyric acid in black raspberry juice during fermentation by Lactobacillus brevis GABA100. Int. J. Food Microbiol. 2009, 130, 12–16. [Google Scholar] [CrossRef]
- Thwe, S.M.; Kobayashi, T.; Luan, T.; Shirai, T.; Onodera, M.; Hamada-Sato, N.; Imada, C. Isolation, characterization, and utilization of γ-aminobutyric acid (GABA)-producing lactic acid bacteria from Myanmar fishery products fermented with boiled rice. Fish. Sci. 2011, 77, 279–288. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lactobacilli Strains | Italian Sourdough |
|---|---|
| L. brevis A7; L. farciminis A11 | Schiacciata (flat, salty bread) |
| L. brevis B1; L. sanfranciscensis B3; L. farciminis B5; L. farciminis B7; L. rossiae B6; L. farciminis B8 | Bread |
| L. plantarum C2 | Tuscan bread |
| L. brevis Ga1; L. rossiae Ga11; L. rossiae Ga12; L. rossiae Ga14; L. sanfranciscensis Gd44; L. rossiae Gd63 | Lagaccio biscuit |
| L. farciminis H3 | Ancient grain bread |
| L. rossiae O1; L. plantarum O4 | Panettone cake |
| Prefermented Dough | ||||||
| Ingredients (% on Total Flour) | PFC W | PFAC W | PFC Am | PFAC Am | PFSD A7 | PFSD A11 |
| Wheat flour | 100 | 100 | 80 | 80 | 80 | 80 |
| Amaranth flour | -- | -- | 20 | 20 | 20 | 20 |
| Water | 58 | 58 | 58 | 58 | 58 | 58 |
| Baker’s yeast | 1 | 1 | 1 | 1 | 1 | 1 |
| L. brevis A7 (log (CFU/g)) | -- | -- | -- | -- | 9 | -- |
| L. farciminis A11 (log (CFU/g)) | -- | -- | -- | -- | -- | 9 |
| Bread | ||||||
| Ingredients (% on total flour) | C W | AC W | C Am | AC Am | SD-A7 | SD-A11 |
| Prefermented dough | 50 | 50 | 50 | 50 | 50 | 50 |
| Wheat flour | 100 | 100 | 80 | 80 | 80 | 80 |
| Amaranth flour | -- | -- | 20 | 20 | 20 | 20 |
| Water | 58 | 58 | 58 | 58 | 58 | 58 |
| Dough yield | 158 | 158 | 158 | 158 | 158 | 158 |
| Dough | Final pH | ΔpH | Final TTA | ΔTTA | Lactic acid | ΔV/V0 × 100 | Lactobacilli | Yeasts |
|---|---|---|---|---|---|---|---|---|
| SD-A7 | 4.35 ± 0.03 a | 0.95 ± 0.39 a | 6.90 ± 0.14 b | 2.80 ± 0.66 a | 4.60 ± 0.63 b | 105 ± 7.07 a | 8.50 ± 0.72 a | 7.35 ± 0.64 a |
| SD-A11 | 4.60 ± 0.18 b | 0.82 ± 0.10 a | 6.45 ± 0.07 a | 2.95 ± 0.78 a | 3.20 ± 0.56 a | 105 ± 21.2 a | 8.59 ± 0.73 a | 7.40 ± 0.62 a |
| C W | C Am | SD-A7 | SD-A11 | |
|---|---|---|---|---|
| Colour | 6.7 ± 1.3 a | 6.2 ± 1.2 a | 6.1 ± 1.2 a | 6.0 ± 1.5 a |
| Aroma | 6.1 ± 1.5 a | 5.6 ± 1.6 a | 5.6 ± 1.5 a | 5.3 ± 2.0 a |
| Consistency | 6.3 ± 1.4 b | 5.8 ± 1.6 ab | 5.1 ± 1.6 a | 5.2 ± 1.5 a |
| General liking | 6.3 ± 1.5 b | 5.3 ± 1.6 a | 4.9 ± 1.6 a | 4.9 ± 1.8 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Venturi, M.; Galli, V.; Pini, N.; Guerrini, S.; Granchi, L. Use of Selected Lactobacilli to Increase γ-Aminobutyric Acid (GABA) Content in Sourdough Bread Enriched with Amaranth Flour. Foods 2019, 8, 218. https://doi.org/10.3390/foods8060218
Venturi M, Galli V, Pini N, Guerrini S, Granchi L. Use of Selected Lactobacilli to Increase γ-Aminobutyric Acid (GABA) Content in Sourdough Bread Enriched with Amaranth Flour. Foods. 2019; 8(6):218. https://doi.org/10.3390/foods8060218
Chicago/Turabian StyleVenturi, Manuel, Viola Galli, Niccolò Pini, Simona Guerrini, and Lisa Granchi. 2019. "Use of Selected Lactobacilli to Increase γ-Aminobutyric Acid (GABA) Content in Sourdough Bread Enriched with Amaranth Flour" Foods 8, no. 6: 218. https://doi.org/10.3390/foods8060218
APA StyleVenturi, M., Galli, V., Pini, N., Guerrini, S., & Granchi, L. (2019). Use of Selected Lactobacilli to Increase γ-Aminobutyric Acid (GABA) Content in Sourdough Bread Enriched with Amaranth Flour. Foods, 8(6), 218. https://doi.org/10.3390/foods8060218