Nutraceutic Potential of Two Allium Species and Their Distinctive Organosulfur Compounds: A Multi-Assay Evaluation

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Allium Vegetables and Single Compounds
2.2. Preparation of the Samples
2.3. In Vivo Assays
2.3.1. Somatic Mutation and Recombination Test (SMART)
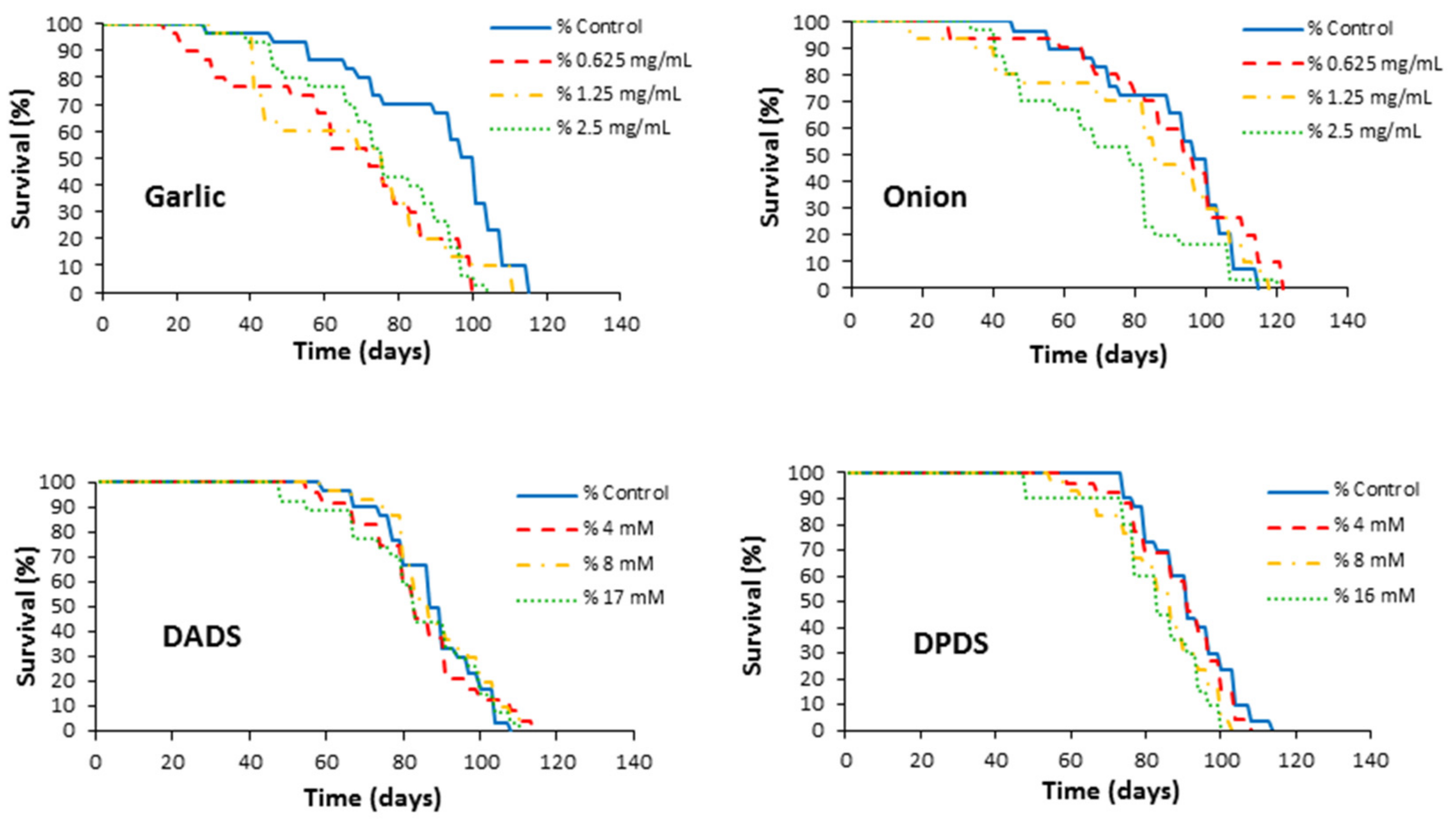
2.3.2. Longevity Assays
2.4. In Vitro Assays
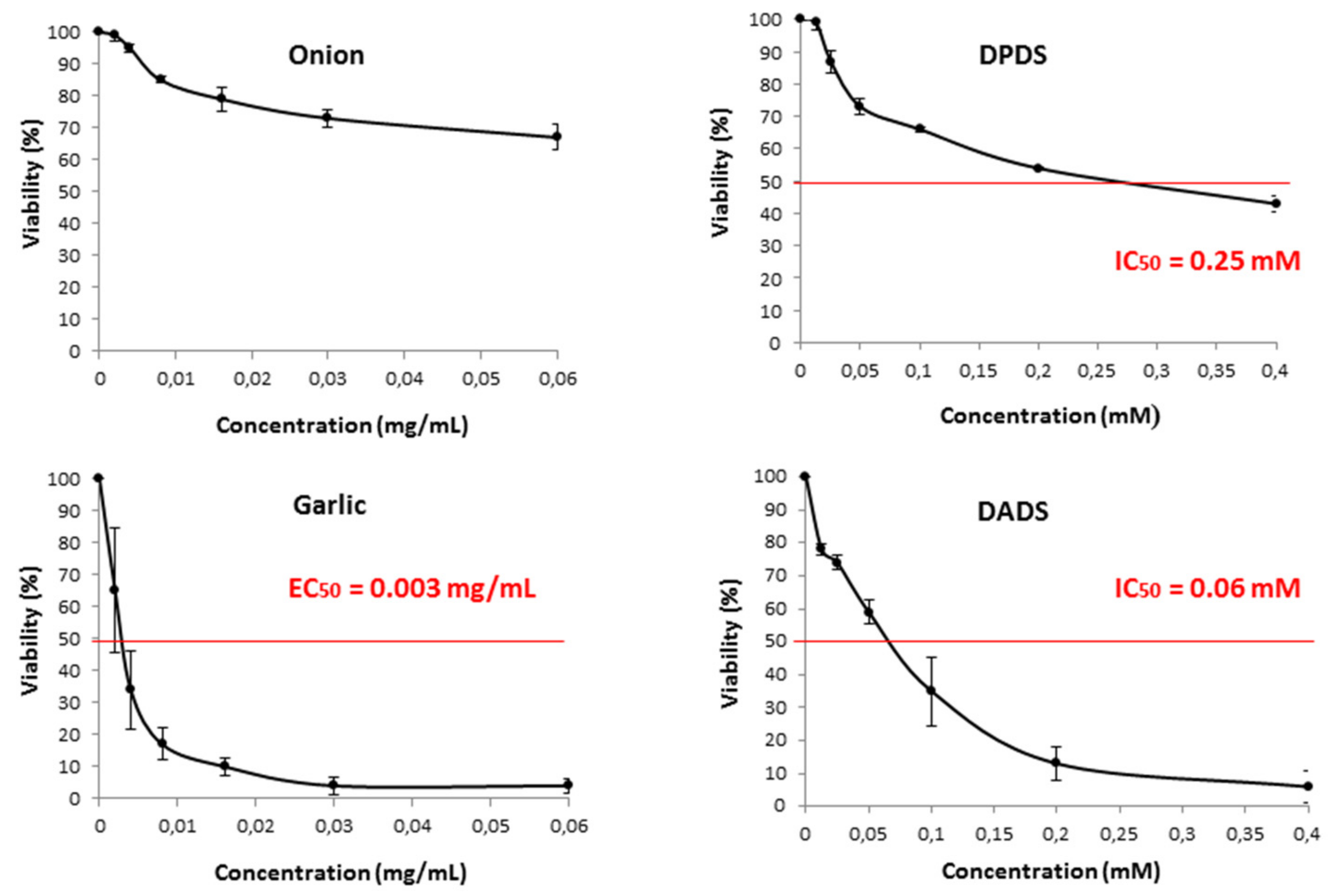
2.4.1. Cell Line Cultures and Cytotoxicity Assay
2.4.2. Inter-Nucleosomal DNA Fragmentation Assay
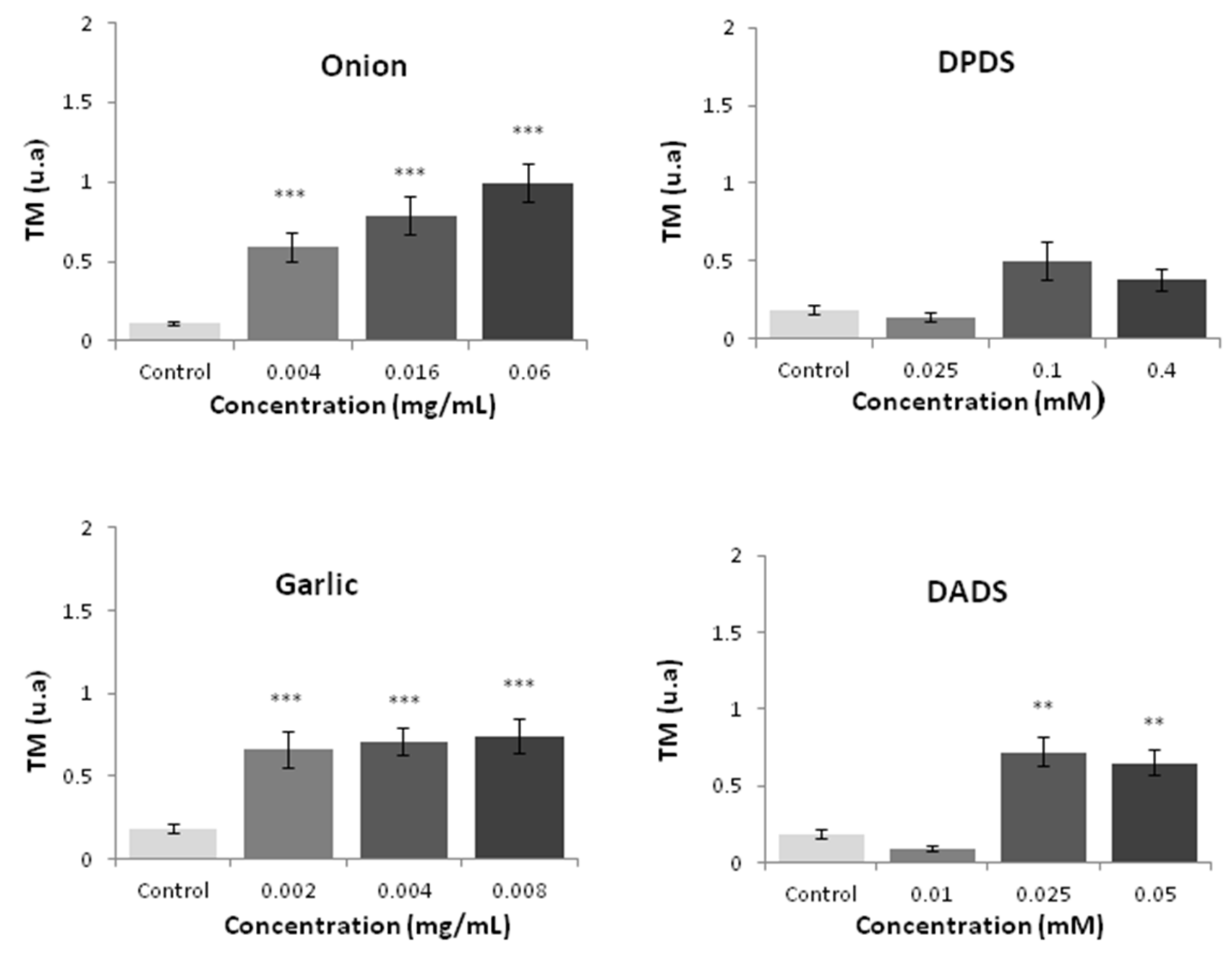
2.4.3. Evaluation of DNA Breakage Ability: Comet Assay
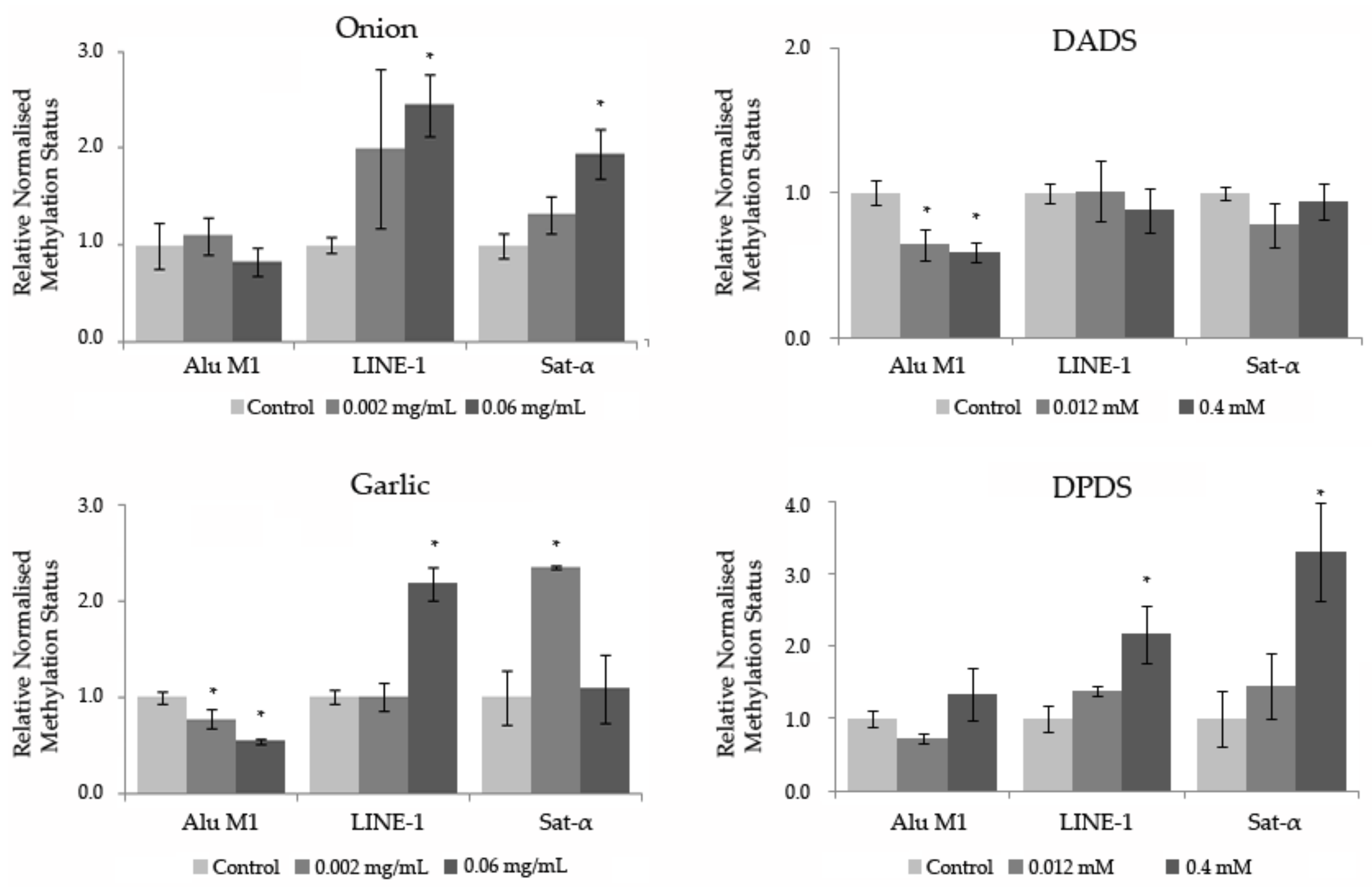
2.4.4. Epigenetic Analysis of Repetitive Sequences on DNA of HL60 Cells
- -
- CT of the target gene was normalized with respect to the referent gene (ΔCT).
- -
- ΔCT of each experimental sample or reference (ΔCT,r) were compared with ΔCT of the calibrator sample (ΔCT,cb): ΔΔCT.
- -
- The relative value of each sample is defined by the formula below.
3. Results
3.1. SMART Test
3.2. Longevity Assays
3.3. Cytotoxicity and Proapoptotic Assays in Leukemia Cells
3.4. DNA Single Strand Breaks
3.5. Methylation Status
4. Discussion
4.1. In Vivo Assessment of the Safety, Protection, and Lifespan Modulation
4.2. In Vitro Assessment of the Cytotoxic, Clastogenic Activities and Methylation Status
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- De Lorgeril, M.; Salen, P. The Mediterranean-style diet for the prevention of cardiovascular diseases. Public Health Nutr. 2006, 9, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Trichopoulou, A.; Vasilopoulou, E. Mediterranean diet and longevity. Br. J. Nutr. 2000, 84, S205–S209. [Google Scholar] [CrossRef] [PubMed]
- Ambrosone, C.B.; Freudenheim, J.L.; Thompson, P.A.; Bowman, E.; Vena, J.E.; Marshall, J.R.; Graham, S.; Laughlin, R.; Nemoto, T.; Shields, P.G. Manganese superoxide dismutase (MnSOD) genetic polymorphisms, dietary antioxidants, and risk of breast cancer. Cancer Res. 1999, 59, 602–606. [Google Scholar] [PubMed]
- Rivlin, R.S. Historical perspective on the use of garlic. J. Nutr. 2001, 131, 951S–954S. [Google Scholar] [CrossRef] [PubMed]
- Lanzotti, V. The analysis of onion and garlic. J. Chromatogr. A 2006, 1112, 3–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santhosha, S.G.; Jamuna, P.; Prabhavathi, S.N. Bioactive components of garlic and their physiological role in health maintenance: A review. Food Biosci. 2013, 3, 59–74. [Google Scholar] [CrossRef]
- Corzo-Martínez, M.; Corzo, N.; Villamiel, M. Biological propierties of onions and garlic. Trends Food Sci. Technol. 2007, 18, 609–625. [Google Scholar] [CrossRef]
- Khadem, S.; Marles, R.J. Monocyclic phenolic acids; hydroxi-polihidroxibenzoic acids: Ocurrence and recent bioactivity studies. Molecules 2010, 15, 7985–8005. [Google Scholar] [CrossRef]
- Heleno, S.A.; Martins, A.; Queiroz, M.J.R.P.; Ferreira, I.C.F.R. Bioactivities of phenolic acids: Metabolites versus parent compounds: A review. Food Chem. 2015, 173, 501–513. [Google Scholar] [CrossRef]
- Yoshinari, O.; Shiojima, Y.; Igarashi, K. Anti-obesity effects of onion extract in Zucker diabetic fatty rats. Nutrients 2012, 4, 1518–1526. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Aoyama, S.; Hamaguchi, N.; Rhi, G.S. Antioxidative and antihypertensive effects of Welsh onion on rats fed with a high-fat high-sucrose diet. Biosci. Biotechnol. Biochem. 2005, 69, 1311–1317. [Google Scholar] [CrossRef]
- Sengupta, A.; Ghosh, S.; Bhattacharjee, S. Allium vegetables in cancer prevention: An overview. Asian Pac. J. Cancer Prev. 2004, 5, 237–245. [Google Scholar]
- Cerella, C.; Dicato, M.; Jacob, C.; Diederich, M. Chemical properties and mechanisms determining the anti-cancer action of garlic-derived organic sulfur compounds. Anti-Cancer Agents Med. Chem. 2011, 11, 267–271. [Google Scholar] [CrossRef]
- Amagase, H.; Petesch, B.L.; Matsuura, H.; Kasuga, S.; Itakura, Y. Intake of garlic and its bioactive components. J. Nutr. 2001, 131, 955S–962S. [Google Scholar] [CrossRef] [PubMed]
- Brodnitz, M.H.; Pascale, J.V.; Van Derslice, L. Flavor components of garlic extract. J. Agric. Food Chem. 1971, 19, 273–275. [Google Scholar] [CrossRef]
- Shukla, Y.; Kalra, N. Cancer chemoprevention with garlic and its constituents. Cancer Lett. 2007, 247, 167–181. [Google Scholar] [CrossRef] [PubMed]
- Shaath, N.A.; Flores, F.B. Egyptian onion oil. Dev. Food Sci. 1998, 40, 443–453. [Google Scholar]
- Benkeblia, N. Free-radical scavenging capacity and antioxidant properties of some selected onions (Allium cepa L.) and garlic (Allium sativum L.) extracts. Braz. Arch. Biol. Technol. 2005, 48, 753–759. [Google Scholar] [CrossRef]
- Park, J.H.; Park, Y.K.; Park, E. Antioxidative and antigenotoxic effects of garlic (Allium sativum L.) prepared by different processing methods. Plant Foods Hum. Nutr. 2009, 64, 244–249. [Google Scholar] [CrossRef]
- Tanaka, S.; Haruma, K.; Yoshihara, M.; Kajilyama, G.; Kira, K.; Amagase, H.; Chayama, K. Aged garlic extract has potential suppressive effect on colorectal adenomas in humans. J. Nutr. 2006, 136, 821S–826S. [Google Scholar] [CrossRef]
- Huang, C.H.; Hsu, F.Y.; Wu, Y.H.; Zhong, L.; Tseng, M.Y.; Kuo, C.J.; Hsu, A.L.; Liang, S.S.; Chiou, S.H. Analysis of lifespan-promoting effect of garlic extract by an integrated metabolo-proteomics approach. J. Nutr. Biochem. 2015, 26, 808–817. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.-J.; Kassie, F.; Mersch-Sundermann, V. The role of reactive oxygen species (ROS) production on diallyl disulfide (DADS) induced apoptosis and cell cycle arrest in human A549 lung carcinoma cells. Mutat. Res. 2005, 579, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.-S.; Chen, G.-W.; Hsia, T.-C.; Ho, H.-C.; Ho, C.-C.; Lin, M.-W.; Lin, S.-S.; Yeh, R.-D.; Ip, S.-W.; Lu, H.-F.; et al. Diallyl disulfide induces apoptosis in human colon cancer cell line (COLO 205) through the induction of reactive oxygen species, endoplasmic reticulum stress, caspases casade and mitochondrial-dependent pathways. Food Chem. Toxicol. 2009, 47, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Jakubikova, J.; Sedlak, J. Garlic-derived organosulfides induce cytotoxicity, apoptosis, cell cycle arrest and oxidative stress in human colon carcinoma cell lines. Neoplasma 2005, 53, 191–199. [Google Scholar]
- Tan, H.; Ling, H.; He, J.; Yi, L.; Zhou, J.; Lin, M.; Su, Q. Inhibition of ERK and activation of p38 are involved in diallyl disulfide induced apoptosis of leukemia HL-60 cells. Arch. Pharm. Res. 2009, 31, 786–793. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Sue, C.C.; Yu, C.S.; Chen, S.C.; Chen, G.W.; Chung, J.G. Diallyl disulfide (DADS) induced apoptosis undergo caspase-3 activity in human bladder cancer T24 cells. Food Chem. Toxicol. 2008, 42, 1543–1552. [Google Scholar] [CrossRef]
- Lin, Y.T.; Yang, J.S.; Lin, S.Y.; Tan, T.W.; Ho, C.C.; Hsia, T.C.; Chiu, T.H.; Yu, C.S.; Lu, H.F.; Weng, Y.S.; et al. Diallyl disulfide (DADS) induces apoptosis in human cervical cancer Ca Ski cells via reactive oxygen species and Ca2+-dependent mitochondria-dependent pathway. Anticancer Res. 2008, 28, 2791–2799. [Google Scholar]
- Gayathri, R.; Gunadharini, D.-N.; Arunkumar, A.; Senthilkumar, K.; Krishnamoorthy, G.; Banudevi, S.; Vignesh, R.C.; Arunakaran, J. Effects of diallyl disulfide (DADS) on expression of apoptosis associated proteins in androgen independent human prostate cancer cells (PC-3). Mol. Cell. Biochem. 2009, 320, 197–203. [Google Scholar] [CrossRef]
- Sundaram, S.-G.; Milner, J.-A. Diallyl disulfide inhibits the proliferation of human tumor cells in culture. BBA-Mol. Basis Dis. 1996, 1315, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Meyers, K.-J.; van der Heide, J.; Liu, R.H. Varietal differences in phenolic content and antioxidant and antiproliferative activities of onions. J. Agric. Food Chem. 2004, 52, 6787–6793. [Google Scholar] [CrossRef]
- Aunanan, A.; Kangsadalampai, K. Effect of Preparation and Temperature treatments on Antimutagenicity against Urethane in Drosophila melanogaster and Antioxidant activity of Three Allium members. Thai J. Toxicol. 2010, 23, 108–116. [Google Scholar]
- Ikken, Y.; Morales, P.; Martinez, A.; Marín, M.L.; Haza, A.I.; Cambero, M.I. Antimutagenic effect of fruit and vegetable ethanolic extracts against N-nitrosamines evaluated by the Ames test. J. Agric. Food Chem. 1999, 47, 3257–3264. [Google Scholar] [CrossRef] [PubMed]
- Seki, T.; Tsuji, K.; Hayato, Y.; Moritomo, T.; Ariga, T. Garlic and onion oils inhibit proliferation and induce differentiation of HL-60 cells. Cancer Lett. 2000, 160, 29–35. [Google Scholar] [CrossRef]
- Wang, Y.; Tian, W.-X.; Ma, X.-F. Inhibitory effects of onion (Allium cepa L.) extract on proliferation of cancer cells and adipocytes via inhibiting fatty acid synthase. Asian Pac. J. Cancer Prev. 2012, 13, 5573–5579. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.-J.; Stahl, T.; Hu, Y.; Kassie, F.; Mersch-Sundermann, V. The production of reactive oxygen species and the mitochondrial membrane potential are modulated during onion oil–induced cell cycle arrest and apoptosis in A549 cells. J. Nutr. 2006, 136, 608–613. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, S.; Ganesh, N. Tumor inhibition and Cytotoxicity assay by aqueous extract of onion (Allium cepa) & Garlic (Allium sativum): An in-vitro analysis. Int. J. Phytomed. 2010, 2, 80–84. [Google Scholar]
- Guyonnet, D.; Belloir, C.; Suschetet, M.; Siess, M.-H.; Le Bon, A.-M. Liver subcellular fractions from rats treated by organosulfur compounds from Allium modulate mutagen activation. Mutat. Res. 2000, 466, 17–26. [Google Scholar] [CrossRef]
- Arranz, N.; Haza, A.I.; García, A.; Möller, L.; Rafter, J.; Morales, P. Protective effects of organosulfur compounds towards N-nitrosamine-induced DNA damage in the single-cell gel electrophoresis (SCGE)/HepG2 assay. Food Chem. Toxicol. 2007, 45, 1662–1669. [Google Scholar] [CrossRef]
- Arranz, N.; Haza, A.I.; García, A.; Delgado, E.; Rafter, J.; Morales, P. Effects of organosulfurs, isothiocyanates and vitamin C towards hydrogen peroxide-induced oxidative DNA damage (strand breaks and oxidized purines/pyrimidines) in human hepatoma cells. Chem. Biol. Interact. 2007, 169, 63–71. [Google Scholar] [CrossRef]
- Singh, S.V.; Pan, S.S.; Srivastava, S.K.; Xia, H.; Hu, X.; Zaren, H.A.; Orchard, J.L. Differential induction of NAD (P) H: Quinone oxidoreductase by anti-carcinogenic organosulfides from garlic. Biochem. Biophys. Res. Commun. 1998, 244, 917–920. [Google Scholar] [CrossRef]
- Merhi, F.; Auger, J.; Rendu, F.; Bauvois, B. Allium compounds, dipropyl and dimethyl thiosulfinates as antiproliferative and differentiating agents of human acute myeloid leukemia cell lines. Biol. Targets Ther. 2008, 2, 885–895. [Google Scholar]
- García, A.; Morales, P.; Arranz, N.; Delgado, M.A.; Rafter, J.; Haza, A.I. Antiapoptotic effects of dietary antioxidants towards N-nitrosopiperidine and N-nitrosodibutylamine-induced apoptosis in HL-60 and HepG2 cells. J. Appl. Toxicol. 2009, 29, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Huen, D.; Morely, T.; Johnson, G.; Gubb, D.; Roote, J.; Adler, P.N. The multiple-wing-hairs gene encodes a novel GBD–FH3 domain-containing protein that functions both prior to and after wing hair initiation. Genetics 2008, 180, 219–228. [Google Scholar] [CrossRef]
- Ren, N.; Charlton, J.; Adler, P.N. The flare gene, which encodes the AIP1 protein of Drosophila, functions to regulate F-actin disassembly in pupal epidermal cells. Genetics 2007, 176, 2223–2234. [Google Scholar] [CrossRef] [PubMed]
- Graf, U.; Wurgler, F.E.; Katz, A.J.; Frei, H.; Juon, H.; Hall, C.B.; Kale, P.G. Somatic mutation and recombination test in Drosophila melanogaster. Environ. Mutagen. 1984, 6, 153–188. [Google Scholar] [CrossRef] [PubMed]
- Anter, J.; Fernández-Bedmar, Z.; Villatoro-Pulido, M.; Demyda-Peyrás, S.; Moreno-Millán, M.; Alonso-Moraga, A.; Muñoz-Serrano, A.; Luque de Castro, M.D. A pilot study on the DNA-protective, cytotoxic, and apoptosis-inducing properties of olive-leaf extracts. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2011, 723, 165–170. [Google Scholar] [CrossRef]
- Block, E. The Organosulfur Chemistry of the Genus AZZium-Implications for the Organic Chemistry of Sulfur. Angew. Chem. Int. Ed. Engl. 1992, 31, 1135–1178. [Google Scholar] [CrossRef]
- Frei, H.; Würgler, F.E. Optimal experimental design and sample size for the statistical evaluation of data from somatic mutation and recombination tests (SMART) in Drosophila. Mutat. Res. 1995, 334, 247–258. [Google Scholar] [CrossRef]
- Abraham, S.K. Antigenotoxicity of coffee in the Drosophila assay for somatic mutation and recombination. Mutagenesis 1994, 9, 383–386. [Google Scholar] [CrossRef]
- Fernández-Bedmar, Z.; Anter, J.; de la Cruz-Ares, S.; Muñoz-Serrano, A. Role of citrus juices and distinctive components in the modulation of degenerative processes: Genotoxicity, antigenotoxicity, cytotoxicity, and longevity in Drosophila. J. Toxicol. Environ. Health A 2011, 74, 1052–1066. [Google Scholar] [CrossRef]
- Lee, K.S.; Lee, B.S.; Semnani, S.; Avanesian, A.; Um, C.Y.; Jeon, H.J.; Seong, K.M.; Yu, K.; Min, K.J.; Jafari, M. Curcumin extends life span, improves health span, and modulates the expression of age-associated aging genes in Drosophila melanogaster. Rejuvenation Res. 2010, 13, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Birnie, G. The HL60 cell line: A model system for studying human myeloid cell differentiation. Br. J. Cancer 1988, 9, 41–45. [Google Scholar]
- Anter, J.; Demyda-Peyrás, S.; Delgado de la Torre, M.P.; Campos-Sánchez, J.; Luque de Castro, M.D.; Munoz-Serrano, A.; Alonso-Moraga, A. Biological and health promoting activity of vinification byproducts produced in Spanish vineyards. S. Afr. J. Enol. Vitic. 2015, 36, 126–133. [Google Scholar] [CrossRef]
- Olive, P.L.; Banáth, J.P. The comet assay: A method to measure DNA damage in individual cells. Nat. Protoc. 2006, 1, 23–29. [Google Scholar] [CrossRef]
- Gyori, B.M.; Venkatachalan, G.; Thiagarajan, P.S.; Hsu, D.; Clement, M.V. OpenComet: An automated tool for comet assay image analysis. Redox Biol. 2014, 2, 457–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serpeloni, J.M.; Bisarro dos Reis, M.; Rodrigues, J.; Campaner dos Santos, L.; Vilegas, W.; Varanda, E.A.; Dokkedal, A.L.; Cólus, I.M.S. In vivo assessment of DNA damage and protective effects of extracts from Miconia species using the comet assay and micronucleus test. Mutagenesis 2008, 23, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Merinas-Amo, T.; Merinas-Amo, R.; Alonso-Moraga, A. A clinical pilot assay of beer consumption: Modulation in the methylation status patterns of repetitive sequences. Sylwan 2017, 161, 134–156. [Google Scholar]
- Deininger, P.L.; Moran, J.V.; Batzer, M.A.; Kazazian, H.H. Mobile elements and mammalian genome evolution. Curr. Opin. Genet. Dev. 2003, 13, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, M. DNA hypomethylation, cancer, the immunodeficiency, centromeric region instability, facial anomalies syndrome and chromosomal rearrangements. J. Nutr. 2002, 132, 2424S–2429S. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Wevrick, R.; Fisher, R.; Ferguson-Smith, M.; Lin, C. Human centromeric DNAs. Hum. Genet. 1997, 100, 291–304. [Google Scholar] [CrossRef]
- Weiner, A.M. SINEs and LINEs: The art of biting the hand that feeds you. Curr. Opin. Cell Biol. 2002, 14, 343–350. [Google Scholar] [CrossRef]
- Weisenberger, D.J.; Campan, M.; Long, T.I.; Kim, M.; Woods, C.; Fiala, E.; Ehrlich, M.; Laird, P.W. Analysis of repetitive element DNA methylation by MethyLight. Nucleic Acids Res. 2005, 33, 6823–6836. [Google Scholar] [CrossRef] [Green Version]
- Nikolaidis, G.; Raji, O.Y.; Markopoulou, S.; Gosney, J.R.; Bryan, J.; Warburton, C.; Walshaw, M.; Sheard, J.; Field, J.K.; Liloglou, T. DNA methylation biomarkers offer improved diagnostic efficiency in lung cancer. Cancer Res. 2012, 72, 5692–5701. [Google Scholar] [CrossRef]
- Liloglou, T.; Bediaga, N.G.; Brown, B.R.; Field, J.K.; Davies, M.P. Epigenetic biomarkers in lung cancer. Cancer Lett. 2014, 342, 200–212. [Google Scholar] [CrossRef] [PubMed]
- Romero-Jimenez, M.; Campos-Sanchez, J.; Analla, M.; Muñoz-Serrano, A.; Alonso-Moraga, A. Genotoxicity and anti-genotoxicity of some traditional medicinal herbs. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2005, 585, 147–155. [Google Scholar] [CrossRef]
- Masuda, Y.; Asada, K.; Satoh, R.; Takada, K.; Kitajima, J. Capillin, a major constituent of Artemisia capillaris Thunb. flower essential oil, induces apoptosis through the mitochondrial pathway in human leukemia HL-60 cells. Phytomedicine 2015, 22, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Reiter, L.T.; Potocki, L.; Chien, S.; Gribskow, M.; Bier, E. A systematic analysis of human disease-associated gene sequences in Drosophila melanogaster. Genome Res. 2001, 11, 1114–1125. [Google Scholar] [CrossRef]
- Jones, M.A.; Grotewiel, M. Drosophila as a model for age-related impairment in locomotor and other behaviors. Exp. Gerontol. 2011, 46, 320–325. [Google Scholar] [CrossRef]
- Pandey, U.B.; Nichols, C.D. Human disease models in Drosophila melanogaster and the role of the fly in therapeutic drug discovery. Pharmacol. Rev. 2011, 63, 411–436. [Google Scholar] [CrossRef]
- Mateo-Fernández, M.; Merinas-Amo, T.; Moreno-Millán, M.; Alonso-Moraga, A.; Demyda Peyrás, S. In Vivo and in vitro genotoxic and epigenetic effects of two types of cola beverages and caffeine: A multiassay approach. BioMed Res. Int. 2016, 2016, 7574843. [Google Scholar] [CrossRef]
- Mukhopadhyay, I.; Chowdhuri, D.K.; Bajpayee, M.; Dhawan, A. Evaluation of in vivo genotoxicity of cypermethrin in Drosophila melanogaster using the alkaline Comet assay. Mutagenesis 2004, 19, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Siddique, H.R.; Chowdhuri, D.K.; Saxena, D.K.; Dhawan, A. Validation of Drosophila melanogaster as an in vivo model for genotoxicity assessment using modified alkaline Comet assay. Mutagenesis 2005, 20, 285–290. [Google Scholar] [CrossRef] [PubMed]
- Martínez, A.; Ikken, I.; Cambero, M.I.; Marín, M.L.; Haza, A.I.; Casas, C.; Morales, P. Mutagenicity and cytotoxicity of fruits and vegetables evaluated by the Ames test and 3-(4,5-dimethylthiazol-2-yl)- 2,5-diphenyltetrazolium bromide (MTT) assay. Food Sci. Technol. Int. 1999, 5, 431–437. [Google Scholar] [CrossRef]
- Chughtai, S.R.; Dhmad, M.A.; Khalid, N.; Mohamed, A.S. Genotoxicity testing of some spices in diploid yeast. Pakisthan J. Bot. 1998, 30, 33–38. [Google Scholar]
- Shukla, Y.; Taneja, P. Antimutagenic effects of garlic extract on chromosomal aberrations. Cancer Lett. 2002, 176, 31–36. [Google Scholar] [CrossRef]
- Abraham, S.K.; Kesavan, P. Genotoxicity of garlic, turmeric and asafoetida in mice. Mutat. Res. 1984, 136, 85–88. [Google Scholar] [CrossRef]
- Sowjanya, B.L.; Devi, K.R.; Madhavi, D. Modulatory effects of garlic extract against the cyclophosphamide induced genotoxicity in human lymphocytes in vitro. J. Environ. Biol. 2009, 30, 663–666. [Google Scholar]
- Kulkarni, M.; Ascough, G.D.; Verschaeve, L.; Baeten, K.; Arruda, M.P.; Van Staden, J. Effect of smoke-water and a smoke-isolated butenolide on the growth and genotoxicity of commercial onion. Sci. Hort. 2010, 124, 434–439. [Google Scholar] [CrossRef]
- Guyonnet, D.; Belloir, C.; Suschetet, M.M.; Siess, M.H.; Le Bon, A.M. Antimutagenic activity of organosulfur compounds from Allium is associated with phase II enzyme induction. Mutat. Res. 2001, 495, 135–145. [Google Scholar] [CrossRef]
- Musk, S.; Clapham, P.; Johnson, I. Cytotoxicity and genotoxicity of diallyl sulfide and diallyl disulfide towards Chinese hamster ovary cells. Food Chem. Toxicol. 1997, 35, 379–385. [Google Scholar] [CrossRef]
- Ramel, C.; Alekperok, U.K.; Ames, B.N.; Kada, T.; Wattenberg, L.W. Inhibitors of mutagenesis and their relevance to carcinogenesis: Report by ICPEMC expert group on antimutagens and desmutagens. Mutat. Res. 1986, 168, 47–65. [Google Scholar] [CrossRef]
- Ito, Y.; Maeda, S.; Sugiyama, T. Suppression of 7, 12-dimethylbenz [a] anthracene-induced chromosome aberrations in rat bone marrow cells by vegetable juices. Mutat. Res. 1986, 172, 55–60. [Google Scholar] [CrossRef]
- Prakash, D.; Singh, B.N.; Upadhyay, G. Antioxidant and free radical scavenging activities of phenols from onion (Allium cepa). Food Chem. 2007, 102, 1389–1393. [Google Scholar] [CrossRef]
- Imai, J.; Ide, N.; Nagae, S.; Moriguchi, T.; Matsuura, H.; Ytakura, Y. Antioxidant and radical scavenging effects of aged garlic extract and its constituents. Planta Med. 1994, 60, 417–420. [Google Scholar] [CrossRef]
- Singh, S.; Abraham, S.; Kesavan, P. Radioprotection of mice following garlic pretreatment. Br. J. Cancer 1996, 27, S102–S104. [Google Scholar]
- Kim, J.M.; Jeon, G.I.; Park, E.J. Protective Effect of Garlic (Allium sativum L.) extracts prepared by different processing methods on DNA damage in human leukocytes. J. Korean Soc. Food Sci. Nutr. 2010, 39, 805–812. [Google Scholar] [CrossRef]
- Sengupta, A.; Ghosh, S.; Das, S. Administration of garlic and tomato can protect from carcinogen induced clastogenicity. Nutr. Res. 2002, 22, 859–866. [Google Scholar] [CrossRef]
- Le Bon, A.M.; Roy, C.; Dupont, C.; Suschetet, M. In vivo antigenotoxic effects of dietary allyl sulfides in the rat. Cancer Lett. 1997, 114, 131–134. [Google Scholar] [CrossRef]
- Anh, N.T.T.; Nishitani, M.; Harada, S.; Yamaguchi, M.; Kamei, K.A. Drosophila model for the screening of bioavailable NADPH oxidase inhibitors and antioxidants. Mol. Cell. Biochem. 2011, 352, 91–98. [Google Scholar] [CrossRef]
- Zhang, Z.; Han, S.; Wang, H.; Wang, T. Lutein extends the lifespan of Drosophila melanogaster. Arch. Gerontol. Geriatr. 2014, 58, 153–159. [Google Scholar] [CrossRef]
- Lei, M.M.; Xu, M.Y.; Zhang, Z.S.; Zhang, M.; Gao, Y.F. The analysis of saccharide in black garlic and its antioxidant activity. Adv. J. Food Sci. Technol. 2014, 6, 755–760. [Google Scholar] [CrossRef]
- Prowse, G.M.; Galloway, T.S.; Foggo, A. Insecticidal activity of garlic juice in two dipteran pests. Agric. For. Entomol. 2006, 8, 1–6. [Google Scholar] [CrossRef]
- Penner, R.; Fedorak, R.N.; Madsen, K.L. Probiotics and nutraceuticals: Non-medicinal treatments of gastrointestinal diseases. Curr. Opin. Pharmacol. 2005, 5, 596–603. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.S.; Ham, Y.A.; Choi, J.H.; Kim, J. Effects of allyl sulfur compounds and garlic extract on the expressions of Bcl-2, Bax, and p53 in non small cell lung cancer cell lines. Exp. Mol. Med. 2000, 32, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Druesne, N.; Pagniez, A.; Mayeur, C.; Thomas, M.; Cherbuy, C.; Duée, P.H.; Martel, P.; Chaumontet, C. Diallyl disulfide (DADS) increases histone acetylation and p21waf1/cip1 expression in human colon tumor cell lines. Carcinogenesis 2004, 25, 1227–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, K.B.; Yoo, S.J.; Ryu, D.G.; Yang, J.Y.; Rho, H.W.; Kim, J.S.; Park, J.W.; Kim, H.R.; Park, B.H. Induction of apoptosis by diallyl disulfide through activation of caspase-3 in human leukemia HL-60 cells. Biochem. Pharmacol. 2002, 63, 41–47. [Google Scholar] [CrossRef]
- Sundaram, S.G.; Milner, J.A. Diallyl disulfide induces apoptosis of human colon tumor cells. Carcinogenesis 1996, 17, 669–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chun, H.S.; Kim, H.J.; Choi, E.H. Modulation of cytochrome P4501-mediated bioactivation of benzo [a] pyrene by volatile allyl sulfides in human hepatoma cells. Biosci. Biotechnol. Biochem. 2001, 65, 2205–2212. [Google Scholar] [CrossRef]
- Iciek, M.; Kwiecien, I.; Wlodek, L. Biological properties of garlic and garlic-derived organosulfur compounds. Environ. Mol. Mutagen. 2009, 50, 247–265. [Google Scholar] [CrossRef]
- Cohen, G.M.; Sun, X.M.; Snowden, R.T.; Dinsdale, D.; Skilleter, D.N. Key morphological features of apoptosis may occur in the absence of internucleosomal DNA fragmentation. Biochem. J. 1992, 286, 331–334. [Google Scholar] [CrossRef]
- Balasenthil, S.; Rao, K.; Nagini, S. Garlic induces apoptosis during 7, 12-dimethylbenz [a] anthracene-induced hamster buccal pouch carcinogenesis. Oral Oncol. 2002, 38, 431–436. [Google Scholar] [CrossRef]
- Cardile, V.; Scifo, C.; Russo, A.; Falsaperla, M.; Morgia, G.; Motta, M.; Renis, M.; Imbriani, E.; Silvestre, G. Involvement of HSP70 in resveratrol-induced apoptosis of human prostate cancer. Anticancer Res. 2002, 23, 4921–4926. [Google Scholar]
- Zulkifli, S.; Luddin, N.; Thirumulu, K.P. Perspectives for the Comet assay in assessing genotoxicity—A review. Curr. Pharmacogenomics Person. Med. 2016, 14, 29–35. [Google Scholar] [CrossRef]
- Fairbairn, D.W.; O’Neill, K.L. Necrotic DNA degradation mimics apoptotic nucleosomal fragmentation comet tail length. In Vitro Cell. Dev. Biol. Anim. 1995, 31, 171–173. [Google Scholar] [CrossRef] [PubMed]
- Belloir, C.; Singh, V.; Daurat, C.; Siess, M.H.; Le Bon, A.M. Protective effects of garlic sulfur compounds against DNA damage induced by direct-and indirect-acting genotoxic agents in HepG2 cells. Food Chem. Toxicol. 2006, 44, 827–834. [Google Scholar] [CrossRef]
- Roman-Gomez, J.; Jimenez-Velasco, A.; Agirre, X.; Castillejo, J.A.; Navarro, G.; San Jose-Eneriz, E.; Garate, L.; Cordeu, L.; Cervantes, F.; Prosper, F. Repetitive DNA hypomethylation in the advanced phase of chronic myeloid leukemia. Leuk. Res. 2008, 32, 487–490. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clones Per Wings (Number of Spots) (1) | ||||||
|---|---|---|---|---|---|---|
| Compounds | N | Small Spots (1–2 cells) | Large Spots (˃2 cells) | Twin Spots | Total Spots | Mann-Whitney Test (3) |
| Controls | ||||||
| H2O | 40 | 0.10 (4) (2) | 0 | 0 | 0.10 (4) | |
| H2O2 (0.12M) | 40 | 0.30 (12) | 0.05 (2) | 0.02 (1) | 0.37 (15) + | Ω |
| Onion (mg/mL) | ||||||
| 0.625 | 40 | 0.17 (7) | 0 | 0 | 0.17 (7) i | Δ |
| 5 | 38 | 0.08 (3) | 0.03 (1) | 0 | 0.10 (4) i | Δ |
| 0.625 + H2O2 | 40 | 0.20 (8) | 0 | 0.05 (2) | 0.25 (10) λ | Ω |
| 5 + H2O2 | 38 | 0.08 (3) | 0 | 0.03 (1) | 0.11 (4) β | |
| Garlic (mg/mL) | ||||||
| 0.625 | 40 | 0.07 (3) | 0 | 0 | 0.07 (3) i | Δ |
| 5 | 40 | 0.05 (2) | 0 | 0 | 0.05 (2) i | Δ |
| 0.625 + H2O2 | 40 | 0.27 (11) | 0 | 0.02 (1) | 0.30 (12) λ | Δ |
| 5 + H2O2 | 40 | 0.15 (6) | 0 | 0 | 0.15 (6) β | |
| DPDS (mM) | ||||||
| 4 | 40 | 0.22 (9) | 0.02 (1) | 0.02 (1) | 0.27 (11) i | Δ |
| 33 | 40 | 0.07 (3) | 0.07 (3) | 0 | 0.15 (6) i | Δ |
| 4 + H2O2 | 40 | 0.20 (8) | 0.05 (2) | 0 | 0.25 (10) λ | Ω |
| 33 + H2O2 | 40 | 0.17 (7) | 0.02 (1) | 0 | 0.20 (8) λ | Ω |
| DADS (mM) | ||||||
| 4 | 40 | 0.15 (6) | 0 | 0 | 0.15 (6) i | Δ |
| 34 | 26 | 0.04 (1) | 0 | 0 | 0.04 (1) i | Δ |
| 4 + H2O2 | 40 | 0.20 (8) | 0.02 (1) | 0 | 0.22 (9) λ | Ω |
| 34 + H2O2 | 40 | 0.02 (1) | 0 | 0 | 0.02 (1) β | |
| Mean Lifespan (Days) | Mean Lifespan Difference (%) a | Health-Span (75th Percentile) (Days) | Health-Span Difference (%) a | |
|---|---|---|---|---|
| Onion (mg/mL) | ||||
| Control | 92.24 ± 3.58 | 0 | 76.00 ± 12.63 | 0 |
| 0.625 | 95.77 ± 3.45 | 4 | 83.00 ± 5.04 | 9 |
| 1.25 | 92.83 ± 3.36 | 1 | 83.00 ± 5.08 | 9 |
| 2.5 | 81.92 ± 4.98 | −11 | 65.00 ± 13.34 | −11 |
| Garlic (mg/mL) | ||||
| Control | 81.25 ± 4.57 | 0 | 51.14 ± 4.31 | 0 |
| 0.625 | 79.51 ± 3.30 | −2 | 58.83 ± 1.76 | 15 |
| 1.25 | 76.21 ± 4.46 | −6 | 47.29 ± 3.73 | −7 |
| 2.5 | 77.68 ± 3.35 | −4 | 53.57 ± 3.48 | 5 |
| DPDS (mM) | ||||
| Control | 91.63 ± 2.06 | 0 | 77.37 ± 1.05 | 0 |
| 4 | 89.58 ± 2.39 | −2 | 73.00 ± 2.80 | −6 |
| 8 | 84.23 ± 2.38* | −8 | 67.12 ± 2.69 ** | −13 |
| 16 | 82.25 ± 3.23* | −10 | 64.20 ± 6.63 * | −17 |
| DADS (mM) | ||||
| Control | 88.00 ± 2.26 | 0 | 73.11 ± 2.40 | 0 |
| 4 | 84.64 ± 3.01 | −4 | 68.00 ± 3.34 | −7 |
| 8 | 88.83 ± 2.36 | 1 | 75.22 ± 2.50 | 3 |
| 17 | 86.21 ± 3.27 | −2 | 62.87 ± 3.96 * | −14 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández-Bedmar, Z.; Demyda-Peyrás, S.; Merinas-Amo, T.; del Río-Celestino, M. Nutraceutic Potential of Two Allium Species and Their Distinctive Organosulfur Compounds: A Multi-Assay Evaluation. Foods 2019, 8, 222. https://doi.org/10.3390/foods8060222
Fernández-Bedmar Z, Demyda-Peyrás S, Merinas-Amo T, del Río-Celestino M. Nutraceutic Potential of Two Allium Species and Their Distinctive Organosulfur Compounds: A Multi-Assay Evaluation. Foods. 2019; 8(6):222. https://doi.org/10.3390/foods8060222
Chicago/Turabian StyleFernández-Bedmar, Zahira, Sebastián Demyda-Peyrás, Tania Merinas-Amo, and Mercedes del Río-Celestino. 2019. "Nutraceutic Potential of Two Allium Species and Their Distinctive Organosulfur Compounds: A Multi-Assay Evaluation" Foods 8, no. 6: 222. https://doi.org/10.3390/foods8060222
APA StyleFernández-Bedmar, Z., Demyda-Peyrás, S., Merinas-Amo, T., & del Río-Celestino, M. (2019). Nutraceutic Potential of Two Allium Species and Their Distinctive Organosulfur Compounds: A Multi-Assay Evaluation. Foods, 8(6), 222. https://doi.org/10.3390/foods8060222