Metabolomics Analysis of the Deterioration Mechanism and Storage Time Limit of Tender Coconut Water during Storage



Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Sample Collection
2.3. Determination of Physical and Chemical Indexes
2.4. Preparing Samples for UPLC-MS/MS Analysis
2.5. Metabolomics Analysis Based on UPLC-MS/MS
2.6. Data Handling and Multivariate Statistical Analysis
3. Results
3.1. Physical and Chemical Indicators of Change
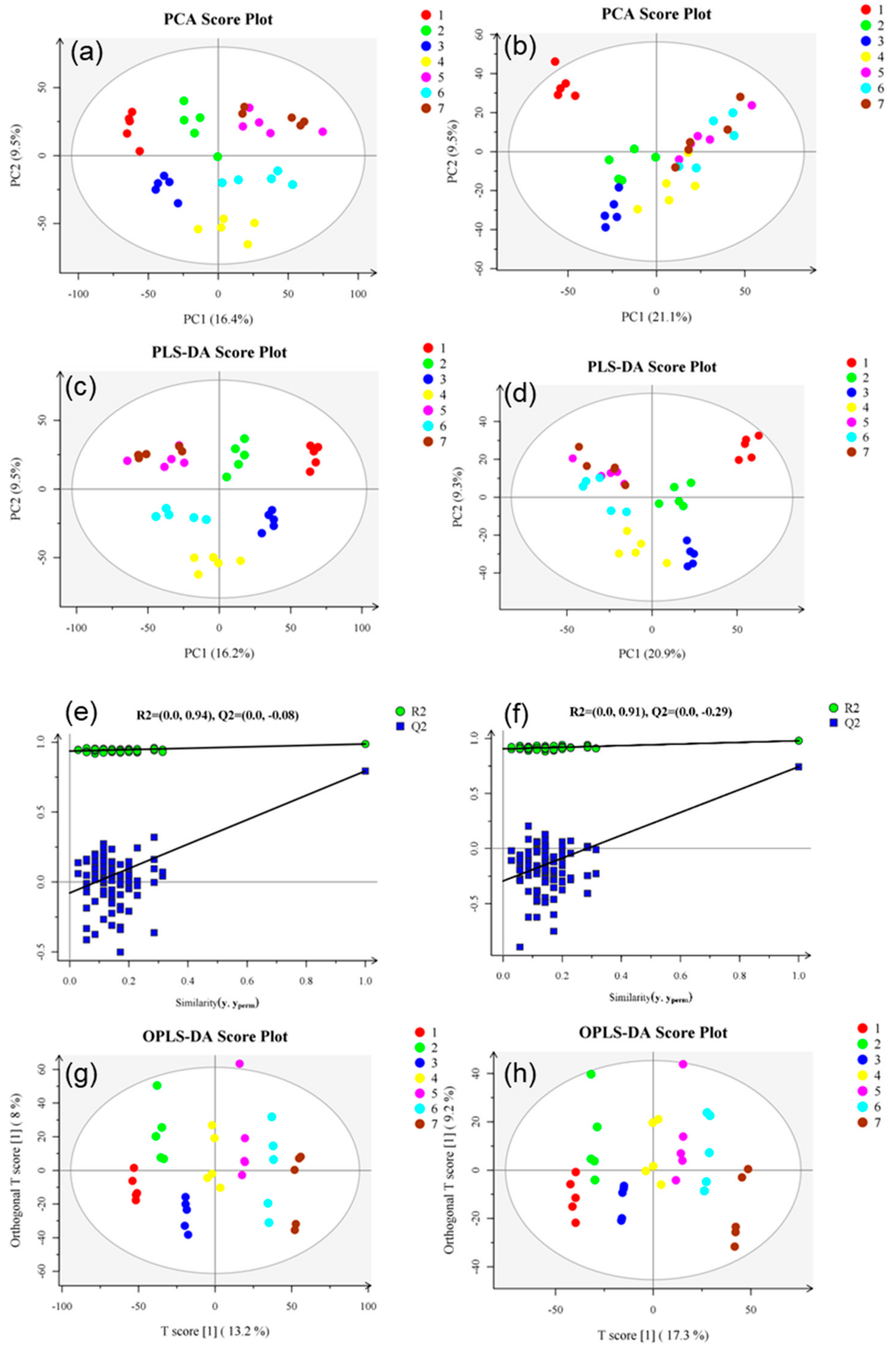
3.2. Metabolic Profile Analysis of Tender Coconut Water during Storage
3.3. Metabolomics Analysis of Tender Coconut Water
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name 1 | m/z 2 | rt 3 | ppm 4 | Exact Mass 5 | Formula | KEGG 6 | Posneg 7 |
|---|---|---|---|---|---|---|---|
| Piperidine | 86.097103 | 130.617 | 8 | 85.1475 | C5H11N | C01746 | pos |
| Pyruvate | 87.007167 | 95.2173 | 19 | 88.06206 | C3H4O3 | C00022 | neg |
| D-Glyceraldehyde | 89.02315 | 23.7384 | 8 | 90.07794 | C3H6O3 | C00577 | neg |
| Acetoin | 89.060419 | 75.4608 | 9 | 88.10512 | C4H8O2 | C00466 | pos |
| Putrescine | 89.107731 | 75.8816 | 10 | 88.1515 | C4H12N2 | C00134 | pos |
| 3-Methyl Pyruvic Acid | 101.02312 | 94.7351 | 13 | 102.0886 | C4H6O3 | C00109 | neg |
| Gamma-Aminobutyric Acid | 104.07055 | 120.134 | 1 | 103.1198 | C4H9NO2 | C00334 | pos |
| Choline | 104.10692 | 90.8299 | 2 | 104.1708 | C5H14NO | C00114 | pos |
| l-Serine | 106.05005 | 87.5235 | 2 | 105.09262 | C3H7NO3 | C00065 | pos |
| Diethanolamine | 106.08649 | 87.4857 | 2 | 105.13568 | C4H11NO2 | C06772 | pos |
| Benzaldehyde | 107.0494 | 353.073 | 3 | 106.1219 | C7H6O | C00193 | pos |
| Uracil | 111.01883 | 122.908 | 11 | 112.08684 | C4H4N2O2 | C00106 | neg |
| l-Proline | 116.07017 | 101.442 | 0 | 115.1305 | C5H9NO2 | C00148 | pos |
| l-Valine | 116.0707 | 112.712 | 9 | 117.14638 | C5H11NO2 | C00183 | neg |
| Glycine Betaine | 118.08633 | 94.5577 | 2 | 117.14638 | C5H11NO2 | C00719 | pos |
| l-Erythrulose | 119.03418 | 94.3132 | 7 | 120.10392 | C4H8O4 | C02045 | neg |
| l-Threonine | 120.0656 | 92.1067 | 2 | 119.1192 | C4H9NO3 | C00188 | pos |
| 4-Hydroxybenzaldehyde | 121.02785 | 536.412 | 14 | 122.12134 | C7H6O2 | C00633 | neg |
| Nicotinic acid | 122.02354 | 121.987 | 4 | 123.10944 | C6H5NO2 | C00253 | neg |
| Nicotinamide | 123.05547 | 116.543 | 0 | 122.12472 | C6H6N2O | C00153 | pos |
| Imidazoleacetic Acid | 127.05018 | 101.293 | 0 | 126.114 | C5H6N2O2 | C02835 | pos |
| 4-Hydroxy-l-Proline | 129.97455 | 135.595 | 16 | 131.1299 | C5H9NO3 | C01015 | neg |
| 5-Oxo-l-Proline | 130.04944 | 195.121 | 2 | 129.114 | C5H7NO3 | C01879 | pos |
| l-Pipecolic Acid | 130.086 | 80.1096 | 1 | 129.157 | C6H11NO2 | C00408 | pos |
| l-Leucine | 130.08668 | 196.186 | 14 | 131.17296 | C6H13NO2 | C00123 | neg |
| Glutaric Acid | 131.03365 | 78.4799 | 13 | 132.11462 | C5H8O4 | C00489 | neg |
| l-Asparagine | 131.04501 | 88.3785 | 9 | 132.118 | C4H8N2O3 | C00152 | neg |
| Agmatine | 131.12911 | 79.1293 | 0 | 130.19162 | C5H14N4 | C00179 | pos |
| l-Isoleucine | 132.10067 | 271.563 | 7 | 131.17296 | C6H13NO2 | C00407 | pos |
| Adenine | 134.04598 | 219.269 | 9 | 135.1269 | C5H5N5 | C00147 | neg |
| Threonate | 135.02871 | 84.5419 | 9 | 136.10332 | C4H8O5 | C01620 | neg |
| p-Salicylic acid | 137.02327 | 115.089 | 8 | 138.12074 | C7H6O3 | C00156 | neg |
| Salicylate | 139.11118 | 706.72 | 4 | 138.122 | C7H6O3 | C00805 | pos |
| 4-Guanidinobutanoic Acid | 146.09205 | 121.961 | 0 | 145.1597 | C5H11N3O2 | C01035 | pos |
| l-Glutamine | 147.07625 | 90.527 | 2 | 146.14458 | C5H10N2O3 | C00064 | pos |
| l-Lysine | 147.11259 | 80.1761 | 1 | 146.18764 | C6H14N2O2 | C00047 | pos |
| O-Acetyl-L-Serine | 148.06013 | 93.1268 | 6 | 147.1293 | C5H9NO4 | C00979 | pos |
| l-Glutamic Acid | 148.06014 | 102.515 | 3 | 147.1293 | C5H9NO4 | C00025 | pos |
| l-Methionine | 150.05874 | 144.366 | 0 | 149.21238 | C5H11NO2S | C00073 | pos |
| 3-Methyladenine | 150.07725 | 120 | 6 | 149.15348 | C6H7N5 | C00913 | pos |
| 9H-Xanthine | 151.02495 | 204.337 | 9 | 152.11102 | C5H4N4O2 | C00385 | neg |
| 3,4-Dimethylbenzoic acid | 151.07466 | 356.095 | 5 | 150.1745 | C9H10O2 | pos | |
| Guanine | 152.05492 | 121.98 | 12 | 151.126 | C5H5N5O | C00242 | pos |
| l-Histidine | 156.07667 | 86.0491 | 2 | 155.15468 | C6H9N3O2 | C00135 | pos |
| 1-Benzylimidazole | 159.09162 | 405.939 | 1 | 158.084398 | C10H10N2 | pos | |
| (R)-2-Hydroxycaprylic Acid | 159.10166 | 390.322 | 7 | 160.2108 | C8H16O3 | neg | |
| l-Phenylalanine | 164.07031 | 330.364 | 12 | 165.18918 | C9H11NO2 | C00079 | neg |
| Capric Acid | 171.13816 | 806.583 | 5 | 172.265 | C10H20O2 | C01571 | neg |
| Dehydroascorbic Acid | 173.00812 | 112.035 | 6 | 174.10824 | C6H6O6 | C05422 | neg |
| Shikimic Acid | 173.04464 | 84.3461 | 5 | 174.1513 | C7H10O5 | C00493 | neg |
| l-Arginine | 173.10357 | 96.2243 | 9 | 174.201 | C6H14N4O2 | C00062 | neg |
| Muscarine | 174.14873 | 335.354 | 1 | 173.141579 | C9H19NO2 | pos | |
| Citrulline | 176.1028 | 94.1013 | 1 | 175.18584 | C6H13N3O3 | C00327 | pos |
| l-Tyrosine | 180.06494 | 122.902 | 9 | 181.18858 | C9H11NO3 | C00082 | neg |
| Keto-D-Fructose | 180.08643 | 96.8 | 7 | 180.15588 | C6H12O6 | C10906 | pos |
| 2-Methylthio-1,3-Benzothiazole | 182.00909 | 815.127 | 1 | 181.278 | C8H7NS2 | C10910 | pos |
| Triethyl Phosphate | 183.07879 | 612.399 | 4 | 182.15466 | C6H15O4P | pos | |
| D-Glucitol | 183.08768 | 93.504 | 8 | 182.17176 | C6H14O6 | C00794 | pos |
| Nonanedioic Acid | 187.09744 | 302.361 | 1 | 188.22094 | C9H16O4 | C08261 | neg |
| 3-Hydroxycapric Acid | 187.13294 | 586.019 | 6 | 188.264 | C10H20O3 | neg | |
| Deethylatrazine | 188.07031 | 405.939 | 3 | 187.0625 | C6H10ClN5 | C06559 | pos |
| Quinic Acid | 191.05511 | 84.3797 | 5 | 192.16658 | C7H12O6 | C00296 | neg |
| N,N-Diethyl-M-Toluamide | 192.13757 | 755.649 | 4 | 191.2695 | C12H17NO | C10935 | pos |
| Butylparaben | 193.08626 | 761.377 | 4 | 194.227 | C11H14O3 | D01420 | neg |
| l-Leucyl-l-Alanine | 201.1237 | 125.201 | 4 | 202.2508 | C9H18N2O3 | neg | |
| Alanyl-DL-Leucine | 203.13918 | 182.305 | 1 | 202.131742 | C9H18N2O3 | pos | |
| ADMA | 203.15006 | 101.974 | 1 | 202.25428 | C8H18N4O2 | C03626 | pos |
| Indolelactic Acid | 206.08104 | 486.255 | 1 | 205.073893 | C11H11NO3 | C02043 | pos |
| 7-Oxo-11-Dodecenoic Acid | 211.13328 | 598.679 | 3 | 212.141237 | C12H20O3 | neg | |
| 8-Chlorotheophylline | 213.01457 | 486.288 | 18 | 214.025751 | C7H7ClN4O2 | neg | |
| Diphenylurea | 213.10194 | 713.78 | 1 | 212.248 | C13H12N2O | pos | |
| Octhilinone | 214.12574 | 808.599 | 1 | 213.34 | C11H19NOS | C18752 | pos |
| Tetradecylamine | 214.25247 | 777.08 | 2 | 213.245649 | C14H31N | pos | |
| 12-Hydroxydodecanoic Acid | 215.16486 | 616.949 | 2 | 216.3172 | C12H24O3 | C08317 | neg |
| Cymiazole | 219.09484 | 780.674 | 1 | 218.087769 | C12H14N2S | pos | |
| (R)-Pantothenic Acid | 220.11782 | 366.856 | 1 | 219.23502 | C9H17NO5 | C00864 | pos |
| Benzanthrone | 231.08369 | 121.982 | 14 | 230.073165 | C17H10O | pos | |
| Confertifoline | 233.15282 | 691.733 | 8 | 234.335 | C15H22O2 | C09376 | neg |
| Dropropizine | 237.15702 | 504.911 | 12 | 236.311 | C13H20N2O2 | pos | |
| l-Cystine | 239.12849 | 758.968 | 3 | 240.30256 | C6H12N2O4S2 | C00491 | neg |
| Uridine | 243.06232 | 123.067 | 0 | 244.20146 | C9H12N2O6 | C00299 | neg |
| Cytidine | 244.09223 | 121.485 | 1 | 243.21674 | C9H13N3O5 | C00475 | pos |
| N,N-Diisopropyl-3-Nitrobenzamide | 251.13594 | 144.97 | 12 | 250.131743 | C13H18N2O3 | pos | |
| Glycerophosphocholine | 258.10912 | 92.1093 | 4 | 257.2213 | C8H20NO6P | C00670 | pos |
| Gamma-Glu-Leu | 261.14469 | 317.015 | 1 | 260.2869 | C11H20N2O5 | pos | |
| l-Phenylalanyl-l-Proline | 263.13858 | 444.483 | 2 | 262.3043 | C14H18N2O3 | pos | |
| 12-oxo-2,3-Dinor-10,15-Phytodienoic Acid | 263.16518 | 831.168 | 0 | 264.36 | C16H24O3 | neg | |
| Adenosine | 268.10341 | 309.894 | 0 | 267.24152 | C10H13N5O4 | C00212 | pos |
| 16-Hydroxy Hexadecanoic Acid | 271.22753 | 833.483 | 1 | 272.4235 | C16H32O3 | C18218 | neg |
| Triethylcitrate | 277.12763 | 716.368 | 2 | 276.283 | C12H20O7 | D06228 | pos |
| Dibutyl Phthalate | 279.16034 | 797.891 | 4 | 278.3435 | C16H22O4 | C14214 | pos |
| Guanosine | 284.0984 | 316.055 | 4 | 283.24092 | C10H13N5O5 | C00387 | pos |
| Epicatechin | 289.0712 | 462.296 | 2 | 290.2681 | C15H14O6 | C09727 | neg |
| Catechin | 291.08533 | 462.105 | 5 | 290.2681 | C15H14O6 | C06562 | pos |
| Terbinafine | 292.21163 | 650.276 | 19 | 291.4299 | C21H25N | C08079 | pos |
| 5-S-Methyl-5-Thioadenosine | 298.09601 | 392.891 | 0 | 297.3347 | C11H15N5O3S | C00170 | pos |
| TMS | 301.14395 | 420.822 | 2 | 300.136159 | C18H20O4 | pos | |
| Dicyclomine | 310.27177 | 755.879 | 8 | 309.4867 | C19H35NO2 | C06951 | pos |
| Triptophenolide | 311.16832 | 790.124 | 10 | 312.172545 | C20H24O3 | neg | |
| 13(S)-HpODE | 311.22274 | 743.286 | 0 | 312.4443 | C18H32O4 | C04717 | neg |
| 9(S)-HpODE | 311.22294 | 775.306 | 0 | 312.4443 | C18H32O4 | C14827 | neg |
| 9,10-DiHOME | 313.23891 | 724.13 | 2 | 314.4602 | C18H34O4 | C14828 | neg |
| Acitretin | 325.18414 | 810.001 | 10 | 326.42934 | C21H26O3 | D02754 | neg |
| Quinine | 325.191 | 652.018 | 0 | 324.4168 | C20H24N2O2 | C06526 | pos |
| Yohimbic Acid | 339.1648 | 831.469 | 19 | 340.41624 | C20H24N2O3 | neg | |
| Vinpocetine | 351.21345 | 714.679 | 19 | 350.455 | C22H26N2O2 | pos | |
| Estradiol Valerate | 355.22845 | 599.118 | 2 | 356.499 | C23H32O3 | C12859 | neg |
| Sinapaldehyde Glucoside | 369.11997 | 96.3153 | 2 | 370.126376 | C17H22O9 | neg | |
| Methyl Arachidonyl Fluorophosphonate | 371.24766 | 763.893 | 9 | 370.243695 | C21H36FO2P | pos | |
| Gentian Violet | 372.24226 | 790.875 | 3 | 371.236135 | C25H30N3 | pos | |
| Tamsulosin | 409.18158 | 464.483 | 6 | 408.512 | C20H28N2O5S | C07124 | pos |
| Procyanidin B2 | 579.1489 | 407.114 | 1 | 578.5202 | C30H26O12 | C17639 | pos |
| Name 1 | 1 vs. 2_VIP 2 | Fold Change_2/1 | Log2(FC_2/1) 3 | p-Value 4 | FDR 5 |
|---|---|---|---|---|---|
| Vinpocetine | 1.869946 | 0.3202 | −1.6429 | 0.0000013 | 0.000254 |
| Glycerophosphocholine | 1.856177 | 42.044 | 5.3938 | 0.0000037 | 0.000517 |
| Choline | 1.855588 | 6.291 | 2.6533 | 0.0000039 | 0.000531 |
| l-Threonine | 1.813788 | 1.6411 | 0.71469 | 0.0000320 | 0.002047 |
| ADMA | 1.808401 | 4.7429 | 2.2458 | 0.0000392 | 0.002287 |
| l-Serine | 1.790296 | 3.2158 | 1.6852 | 0.0000727 | 0.003252 |
| Keto-D-Fructose | 1.77673 | 0.20739 | −2.2696 | 0.0001089 | 0.00426 |
| l-Histidine | 1.740644 | 2.6231 | 1.3913 | 0.0002689 | 0.007746 |
| l-Lysine | 1.739258 | 2.5496 | 1.3503 | 0.0002773 | 0.007874 |
| l-Methionine | 1.734536 | 4.0561 | 2.0201 | 0.0003074 | 0.00837 |
| Nicotinamide | 1.727199 | 5.4063 | 2.4346 | 0.0003589 | 0.009191 |
| Salicylate | 1.721537 | 1.1119 | 0.15298 | 0.0004026 | 0.010029 |
| l-Pipecolic acid | 1.648997 | 2.1215 | 1.0851 | 0.0013724 | 0.020324 |
| Agmatine | 1.629025 | 0.097098 | −3.3644 | 0.0018093 | 0.02423 |
| Guanine | 1.62376 | 2.0952 | 1.0671 | 0.0019395 | 0.025193 |
| Indolelactic acid | 1.621818 | 4.6738 | 2.2246 | 0.0019892 | 0.025604 |
| N,N-Diisopropyl-3-Nitrobenzamide | 1.614857 | 2.2462 | 1.1675 | 0.0021749 | 0.027391 |
| Dropropizine | 1.576419 | 0.89283 | −0.16354 | 0.0034327 | 0.035944 |
| 3-Methyladenine | 1.569726 | 2.1357 | 1.0947 | 0.0036958 | 0.037589 |
| l-Phenylalanyl-L-Proline | 1.54751 | 2.9302 | 1.551 | 0.0046725 | 0.04372 |
| O-Acetyl-l-Serine | 1.534309 | 2.9405 | 1.5561 | 0.0053332 | 0.048166 |
| Glycine Betaine | 1.526096 | 4.1876 | 2.0661 | 0.0057765 | 0.050485 |
| Putrescine | 1.49701 | 0.56468 | −0.8245 | 0.0075577 | 0.058673 |
| Adenosine | 1.492166 | 7.3509 | 2.8779 | 0.0078881 | 0.060157 |
| Citrulline | 1.484936 | 1.6943 | 0.76068 | 0.0084002 | 0.062645 |
| Imidazoleacetic acid | 1.414003 | 1.6133 | 0.69005 | 0.0147345 | 0.087407 |
| 4-Guanidinobutanoic acid | 1.390837 | 12.812 | 3.6794 | 0.0173733 | 0.096289 |
| 5-S-Methyl-5-Thioadenosine | 1.368643 | 4.2794 | 2.0974 | 0.0201933 | 0.105666 |
| Dibutyl Phthalate | 1.363868 | 3.1017 | 1.6331 | 0.0208389 | 0.107572 |
| Muscarine | 1.316996 | 3.677 | 1.8785 | 0.0279490 | 0.12836 |
| Piperidine | 1.275156 | 1.9234 | 0.94366 | 0.0355599 | 0.148035 |
| Tamsulosin | 1.271617 | 4.6915 | 2.23 | 0.0362616 | 0.149595 |
| (R)-Pantothenic acid | 1.224623 | 1.732 | 0.7924 | 0.0464829 | 0.171966 |
| 3,4-Dimethylbenzoic acid | 1.222463 | 0.18966 | −2.3985 | 0.0469943 | 0.173234 |
| Quinic acid | 1.780892 | 4.5158 | 2.175 | 0.0000016 | 0.000236 |
| l-Erythrulose | 1.739383 | 0.32058 | −1.6412 | 0.0000213 | 0.001223 |
| Pyruvate | 1.736553 | 0.354 | −1.4982 | 0.0000241 | 0.0013 |
| 3-Methyl Pyruvic acid | 1.728988 | 0.32725 | −1.6115 | 0.0000332 | 0.001502 |
| Threonate | 1.712791 | 3.4506 | 1.7868 | 0.0000607 | 0.002173 |
| Sinapaldehyde Glucoside | 1.701181 | 3.2492 | 1.7001 | 0.0000889 | 0.002691 |
| 8-Chlorotheophylline | 1.700766 | 0.39072 | −1.3558 | 0.0000901 | 0.0027 |
| l-Asparagine | 1.657314 | 5.2423 | 2.3902 | 0.0002870 | 0.006038 |
| Adenine | 1.585462 | 1.975 | 0.98184 | 0.0011264 | 0.015959 |
| 9(S)-HpODE | 1.504033 | 2.4481 | 1.2917 | 0.0034110 | 0.032279 |
| 9H-Xanthine | 1.489229 | 0.42715 | −1.2272 | 0.0040398 | 0.03549 |
| Glutaric acid | 1.472775 | 3.4595 | 1.7906 | 0.0048316 | 0.039341 |
| D-Glyceraldehyde | 1.457553 | 0.20576 | −2.281 | 0.0056575 | 0.042594 |
| Yohimbic acid | 1.447777 | 37.215 | 5.2178 | 0.0062382 | 0.045236 |
| l-Arginine | 1.428014 | 3.3292 | 1.7352 | 0.0075394 | 0.050283 |
| l-Leucine | 1.378902 | 2.4788 | 1.3096 | 0.0115875 | 0.063966 |
| 12-oxo-2,3-Dinor-10,15-Phytodienoic acid | 1.360253 | 2.2232 | 1.1527 | 0.0134607 | 0.070293 |
| Confertifoline | 1.346373 | 0.28192 | −1.8266 | 0.0149854 | 0.075265 |
| Uridine | 1.327309 | 3.6455 | 1.8661 | 0.0172704 | 0.082697 |
| Nonanedioic acid | 1.311257 | 9.5381 | 3.2537 | 0.0193733 | 0.088133 |
| 4-Hydroxy-l-Proline | 1.256131 | 0.52658 | −0.92528 | 0.0279380 | 0.109387 |
| 4-Hydroxybenzaldehyde | 1.161319 | 2.5676 | 1.3604 | 0.0481468 | 0.151292 |
References
- Xiao, Y.; Xu, P.W.; Fan, H.K.; Baudouin, L.; Xia, W.; Bocs, S.; Xu, J.Y.; Li, Q.; Guo, A.P.; Zhou, L.X.; et al. The genome draft of coconut (Cocos nucifera). GigaScience 2017, 6, 11. [Google Scholar] [CrossRef]
- Yong, J.W.H.; Ge, L.Y.; Ng, Y.F.; Tan, S.N. The Chemical Composition and Biological Properties of Coconut (Cocos nucifera L.) Water. Molecules 2009, 14, 5144–5164. [Google Scholar] [CrossRef] [Green Version]
- Kuberski, T.; Roberts, A.; Linehan, B.; Bryden, R.N.; Teburae, M. Coconut water as a rehydration fluid. N. Z. Med. J. 1979, 90, 98–100. [Google Scholar] [CrossRef]
- DebMandal, M.; Mandal, S. Coconut (Cocos nucifera L.:Arecaceae):In health promotion and disease prevention. Asian Pac. J. Trop. Med. 2011, 4, 241–247. [Google Scholar] [CrossRef] [Green Version]
- Alleyne, T.; Roache, S.; Thomas, C.; Shirley, A. The control of hypertension by use of coconut water and mauby: Two tropical food drinks. West Indian Med. J. 2005, 54, 3–8. [Google Scholar] [CrossRef]
- Patel, R.; Asplin, J.; Granja, I.; Jiang, P.; Osann, K.; Landman, J.; Clayman, R. PD17-03 CAN COCONUT WATER CONSUMPTION POTENTIALLY PREVENT KIDNEY STONES? J. Urol. 2018. [Google Scholar] [CrossRef] [Green Version]
- Anurag, P.; Rajamohan, T. Cardioprotective effect of tender coconut water in experimental myocardial infarction. Plant Foods Hum. Nutr. 2003, 58, 1–12. [Google Scholar] [CrossRef]
- Campbellfalck, D.; Thomas, T.; Falck, T.M.; Tutuo, N.; Clem, K. The intravenous use of coconut water. Am. J. Emerg. Med. 2000, 18, 108–111. [Google Scholar] [CrossRef]
- Overbeek, J.V.; Conklin, M.E.; Blakeslee, A.F. Factors in Coconut Milk Essential for Growth and Development of Very Young Datura Embryos. Science 1941, 94, 350–351. [Google Scholar] [CrossRef]
- Verdeil, J.L.; Hocher, V. Digestion and absorption of food in plants: A plant stomach. Trends Plant Sci. 2002, 7, 280–281. [Google Scholar] [CrossRef]
- Ang, S.L.P.; Yong, J.W.H. A protocol for in vitro germination and sustainable growth of two tropical mistletoes. Plant Cell Tissue Organ Cult. 2005, 80, 221–228. [Google Scholar] [CrossRef]
- Schmauder, H.P.; George, E.F.; Sherrington, P.D. Plant Propagation by Tissue Culture—Handbook and Directory of Commercial Laboratories. VIII + 709 S., 12 Abb., 40 Tab. Eversley, Basingstoke 1984. Exegetics Limited. £ 65.00. ISBN: 0-9509325-0-7. J. Basic Microbiol. 2010, 25, 475. [Google Scholar] [CrossRef]
- Haseena, M.; Bai, K.V.K.; Padmanabhan, S. Post-harvest quality and shelf-life of tender coconut. J. Food Sci. Technol. 2010, 47, 686–689. [Google Scholar] [CrossRef] [Green Version]
- Jirapong, C.; Wongs-Aree, C.; Noichinda, S.; Uthairatanakij, A.; Kanlayanarat, S. Assessment of volatile and non-volatile organic compounds in the liquid endosperm of young ‘Nam Hom’ coconut (Cocos nucifera L.) at two stages of maturity. J. Pomol. Hortic. Sci. 2015, 90, 477–482. [Google Scholar] [CrossRef]
- Lin, F.M.; Wilkens, W.F. Volatile flavor components of coconut meat. J. Food Sci. 2010, 35, 538–539. [Google Scholar] [CrossRef]
- Jirapong, C.; Changprasert, S.; Kanlayanarat, S.; Uthairatanakij, A.; Bodhipadma, K.; Noichinda, S.; Wongs-Aree, C. Characterization of the liquid endosperm attributes in young coconut fruit during storage. Int. Food Res. J. 2018, 25, 2650–2656. Available online: http://www.ifrj.upm.edu.my/ifrj-2018-25-issue-6.html (accessed on 14 August 2019).
- Kang, J.; Zhu, L.; Lu, J.; Zhang, X. Application of metabolomics in autoimmune diseases: Insight into biomarkers and pathology. J. Neuroimmunol. 2015, 279, 25–32. [Google Scholar] [CrossRef]
- Fiehn, O. Metabolomics–the link between genotypes and phenotypes. Plant Mol. Biol. 2002, 48, 155–171. [Google Scholar] [CrossRef]
- Jones, O.A.H.; Cheung, V.L. An introduction to metabolomics and its potential application in veterinary science. Comp. Med. 2007, 57, 436–442. [Google Scholar] [CrossRef]
- Fiehn, O.; Kopka, J.; Trethewey, R.N.; Willmitzer, L. Identification of uncommon plant metabolites based on calculation of elemental compositions using gas chromatography and quadrupole mass spectrometry. Anal. Chem. 2000, 72, 3573–3580. [Google Scholar] [CrossRef]
- Fiehn, O.; Kopka, J.; Dörmann, P.; Altmann, T.; Trethewey, R.N.; Willmitzer, L. Metabolite profiling for plant functional genomics. Nat. Biotechnol. 2000, 18, 1157–1161. [Google Scholar] [CrossRef]
- Miller, S.B.; Heuberger, A.L.; Broeckling, C.D.; Jahn, C.E. Non-Targeted Metabolomics Reveals Sorghum Rhizosphere-Associated Exudates are Influenced by the Belowground Interaction of Substrate and Sorghum Genotype. Int. J. Mol. Sci. 2019, 20, 431. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Beger, R.D.; Schnackenberg, L.K. Metabolomics as a tool for personalizing medicine: 2012 update. Pers. Med. 2013, 10, 149–161. [Google Scholar] [CrossRef]
- Field, K.J.; Lake, J.A. Environmental metabolomics links genotype to phenotype and predicts genotype abundance in wild plant populations. Physiol. Plant. 2011, 142, 352–360. [Google Scholar] [CrossRef]
- Longnecker, K.; Futrelle, J.; Coburn, E.; Soule, M.C.K.; Kujawinski, E.B. Environmental metabolomics: Databases and tools for data analysis. Mar. Chem. 2015, 177, 366–373. [Google Scholar] [CrossRef]
- Bundy, J.G.; Davey, M.P.; Viant, M.R. Environmental metabolomics: A critical review and future perspectives. Metabolomics 2009, 5, 3. [Google Scholar] [CrossRef]
- Dunn, W.B.; David, B.; Paul, B.; Eva, Z.; Sue, F.M.; Nadine, A.; Marie, B.; Knowles, J.D.; Antony, H.; Haselden, J.N. Procedures for large-scale metabolic profiling of serum and plasma using gas chromatography and liquid chromatography coupled to mass spectrometry. Nat. Protoc. 2011, 6, 1060–1083. [Google Scholar] [CrossRef]
- Eva, Z.; Dunn, W.B.; David, B.; Sue, F.M.; Carroll, K.M.; Paul, B.; Steve, O.H.; Knowles, J.D.; Antony, H.; Wilson, I.D. Development of a robust and repeatable UPLC-MS method for the long-term metabolomic study of human serum. Anal. Chem. 2009, 81, 1357–1364. [Google Scholar] [CrossRef]
- Timothy, S.; Hilary, M.; Robert, P.; Wilson, A.J.; Wilson, I.D. A pragmatic and readily implemented quality control strategy for HPLC-MS and GC-MS-based metabonomic analysis. Analyst 2006, 131, 1075–1078. [Google Scholar] [CrossRef]
- Smith, C.A.; Want, E.J.; Grace, O.M.; Ruben, A.; Gary, S. XCMS: Processing Mass Spectrometry Data for Metabolite Profiling Using Nonlinear Peak Alignment, Matching, and Identification. Anal. Chem. 2006, 78, 779–787. [Google Scholar] [CrossRef]
- Tautenhahn, R.; Patti, G.J.; Rinehart, D.; Siuzdak, G. XCMS Online: A Web-Based Platform to Process Untargeted Metabolomic Data. Anal. Chem. 2012, 84, 5035–5039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thévenot, E.A.; Aurélie, R.; Ying, X.; Eric, E.; Christophe, J. Analysis of the Human Adult Urinary Metabolome Variations with Age, Body Mass Index, and Gender by Implementing a Comprehensive Workflow for Univariate and OPLS Statistical Analyses. J. Proteome Res. 2015, 14, 3322–3335. [Google Scholar] [CrossRef] [PubMed]
- Famiani, F.; Cultrera, N.A.; Casulli, V.; Proietti, P.; Standardi, A.; Chen, Z.H.; Leegood, R.C.; Walker, R.P. Phosphoenolpyruvate carboxykinase and its potential role in the catabolism of organic acids in the flesh of soft fruit during ripening. J. Exp. Bot. 2005, 56, 2959–2969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trygg, J.; Wold, S. Orthogonal projections to latent structures (O-PLS). J. Chemom. 2002, 16, 119–128. [Google Scholar] [CrossRef]
- Wang, J.B.; Pu, S.B.; Sun, Y.; Li, Z.F.; Niu, M.; Yan, X.Z.; Zhao, Y.L.; Wang, L.F.; Qin, X.M.; Ma, Z.J. Metabolomic Profiling of Autoimmune Hepatitis: The Diagnostic Utility of Nuclear Magnetic Resonance Spectroscopy. J. Proteome Res. 2014, 13, 3792–3801. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Xi, C.; Xie, C.; Sun, L.; Ding, J.; Cai, H.; Singh, P.K. Longitudinal Metabolomics Profiling of Parkinson’s Disease-Related α-Synuclein A53T Transgenic Mice. PLoS ONE 2015, 10, e0136612. [Google Scholar] [CrossRef]
- Z-Score. 2009. Available online: http://www.doc88.com/p-3377640156606.html (accessed on 12 August 2019). [CrossRef]
- Gary, D.; Tom, S.; Verrall, S.R.; Sean, C.; McNicol, J.W.; Gavin, R.; Shepherd, L.V.T.; Davies, H.V.; Derek, S. Phytochemical diversity in tubers of potato cultivars and landraces using a GC-MS metabolomics approach. J. Agric. Food Chem. 2008, 56, 10280–10291. [Google Scholar] [CrossRef]
- Rao, G.; Sui, J.; Zhang, J. Metabolomics reveals significant variations in metabolites and correlations regarding the maturation of walnuts (Juglans regia L.). Biol. Open 2016, 5, 829–836. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.; Wishart, D.S. MetPA: A web-based metabolomics tool for pathway analysis and visualization. Bioinformatics 2010, 26, 2342–2344. [Google Scholar] [CrossRef] [Green Version]
- Sekowska, A.; Danchin, A. The methionine salvage pathway in Bacillus subtilis. BMC Microbiol. 2002, 2, 8. [Google Scholar] [CrossRef] [PubMed]
- Sekowska, A.; Dénervaud, V.; Ashida, H.; Michoud, K.; Haas, D.; Yokota, A.; Danchin, A. Bacterial variations on the methionine salvage pathway. BMC Microbiol. 2004, 4, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogata, H.; Goto, S.; Sato, K.; Fujibuchi, W.; Bono, H.; Kanehisa, M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 27, 29–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurihara, S.; Oda, S.; Kumagai, H.; Suzuki, H. γ-Glutamyl-γ-aminobutyrate hydrolase in the putrescine utilization pathway of Escherichia coli K-12. FEMS Microbiol. Lett. 2006, 256, 318–323. [Google Scholar] [CrossRef]
- Kurihara, S.; Oda, S.; Kato, K.; Kim, H.G.; Koyanagi, T.; Kumagai, H.; Suzuki, H. A Novel Putrescine Utilization Pathway Involves γ-Glutamylated Intermediates of Escherichia coli K-12. J. Biol. Chem. 2005, 280, 4602–4608. [Google Scholar] [CrossRef] [Green Version]
- Kelly, S.; Jing, Y.; Hohn, M.J.; Brian, J.; Devine, K.M.; Dieter, S.L. From one amino acid to another: tRNA-dependent amino acid biosynthesis. Nucleic Acids Res. 2008, 36, 1813–1825. [Google Scholar] [CrossRef] [Green Version]
- Carla, P.; Alexandre, A.; Amélie, B.; Winbush, S.A.M.; McCloskey, J.A.; Crain, P.F.; Wood, J.L.; Dieter, S.L. An aminoacyl-tRNA synthetase that specifically activates pyrrolysine. Proc. Natl. Acad. Sci. USA 2004, 101, 12450–12454. [Google Scholar] [CrossRef] [Green Version]
- Hai, X.; Yuzhen, Z.; Xiaokui, G.; Shuangxi, R.; Staempfli, A.A.; Juishen, C.; Weihong, J.; Guoping, Z. Isoleucine biosynthesis in Leptospira interrogans serotype lai strain 56601 proceeds via a threonine-independent pathway. J. Bacteriol. 2004, 186, 5400–5409. [Google Scholar] [CrossRef] [Green Version]









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Chen, W.; Chen, H.; Zhong, Q.; Yun, Y.; Chen, W. Metabolomics Analysis of the Deterioration Mechanism and Storage Time Limit of Tender Coconut Water during Storage. Foods 2020, 9, 46. https://doi.org/10.3390/foods9010046
Zhang Y, Chen W, Chen H, Zhong Q, Yun Y, Chen W. Metabolomics Analysis of the Deterioration Mechanism and Storage Time Limit of Tender Coconut Water during Storage. Foods. 2020; 9(1):46. https://doi.org/10.3390/foods9010046
Chicago/Turabian StyleZhang, Yunwu, Wenxue Chen, Haiming Chen, Qiuping Zhong, Yonghuan Yun, and Weijun Chen. 2020. "Metabolomics Analysis of the Deterioration Mechanism and Storage Time Limit of Tender Coconut Water during Storage" Foods 9, no. 1: 46. https://doi.org/10.3390/foods9010046
APA StyleZhang, Y., Chen, W., Chen, H., Zhong, Q., Yun, Y., & Chen, W. (2020). Metabolomics Analysis of the Deterioration Mechanism and Storage Time Limit of Tender Coconut Water during Storage. Foods, 9(1), 46. https://doi.org/10.3390/foods9010046