Potato Tuber Chemical Properties in Storage as Affected by Cultivar and Nitrogen Rate: Implications for Acrylamide Formation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Analysis
2.2. Statistical Analysis
3. Results and Discussion
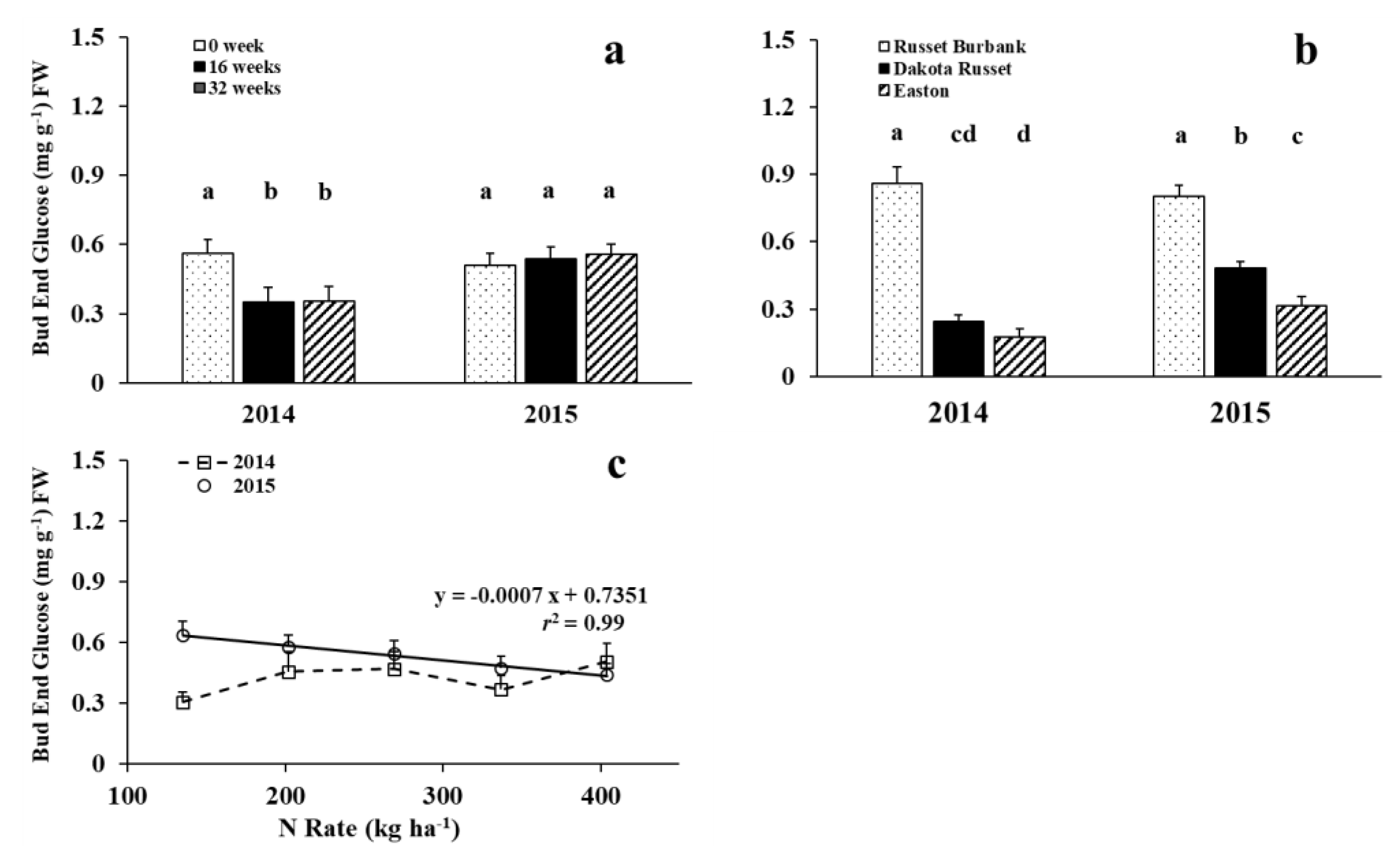
3.1. Glucose Concentrations
3.2. Concentrations of Acrylamide and Acrylamide Precursors
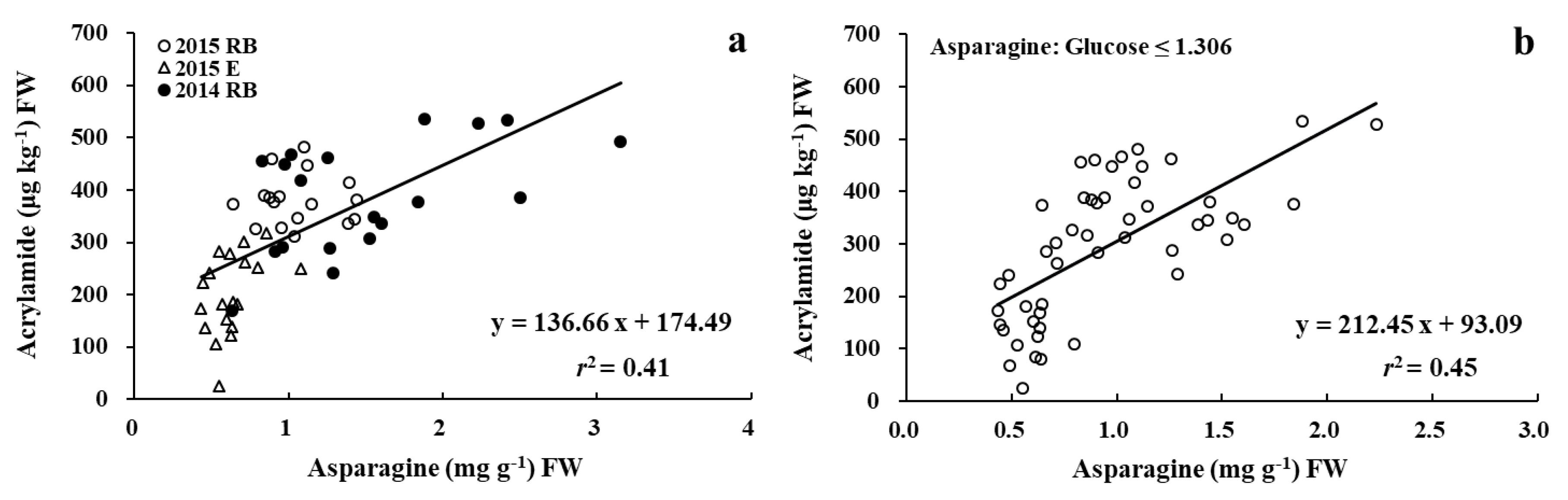
3.3. Relationships between Acrylamide, Glucose, and Asparagine
3.4. Relationships between Fry Color, Glucose, and Acrylamide at 16 Weeks of Storage
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mottram, D.S.; Wedzicha, B.L.; Dodson, A.T. Food chemistry: Acrylamide is formed in the Maillard reaction. Nature 2002, 419, 448–449. [Google Scholar] [CrossRef]
- Cummins, E.; Butlet, F.; Brunton, N.; Gormley, R. Factors affecting acrylamide formation in processed potato products—A simulation approach. In Proceedings of the 13th World Congress of Food Science and Technology, Nantes, France, 17–21 September 2006. [Google Scholar]
- Visvanathan, R.; Krishnakumar, T. Acrylamide in Food Products: A Review. J. Food Process. Technol. 2014, 5, 344. [Google Scholar] [CrossRef] [Green Version]
- Kooman, P.L.; Haverkort, A.J. Modelling development and growth of the potato crop influenced by temperature and daylength: LINTUL-POTATO. In Proceedings of the Potato Ecology and Modelling of Crops under Conditions Limiting Growth, Wageningen, The Netherlands, 17–19 May 1994; Volume 3, pp. 41–59. [Google Scholar]
- Amrein, T.M.; Bachmann, S.; Noti, A.; Biedermann, M.; Barbosa, M.F.; Biedermann-Brem, S.; Grob, K.; Keiser, A.; Realini, P.; Escher, F.; et al. Potential of acrylamide formation, sugars, and free asparagine in potatoes: A comparison of cultivars and farming systems. J. Agric. Food Chem. 2003, 51, 5556–5560. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Singh, B.P.; Kumar, P. An overview of the factors affecting sugar content of potatoes. Ann. Appl. Biol. 2004, 145, 247–256. [Google Scholar] [CrossRef]
- Rommens, C.M.; Yan, H.; Sword, K.; Richael, C.; Ye, J. Low-acrylamide French fries and potato chips. Plant Biotechnol. J. 2008, 6, 843–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, J.; Shakya, R.; Shrestha, P.; Rommens, C.M. Tuber-specific silencing of the acid invertase gene substantially lowers the acrylamide-forming potential of potato. J. Agric. Food Chem. 2010, 58, 12162–12167. [Google Scholar] [CrossRef]
- Bethke, P.C.; Bussan, A.J. Acrylamide in processed potato products. Am. J. Potato Res. 2013, 90, 403–424. [Google Scholar] [CrossRef]
- Muttucumaru, N.; Powers, S.J.; Elmore, J.S.; Mottram, D.S.; Halford, N.G. Effects of nitrogen and sulfur fertilization on free amino acids, sugars, and acrylamide-forming potential in potato. J. Agric. Food Chem. 2013, 61, 6734–6742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muttucumaru, N.; Powers, S.J.; Elmore, J.S.; Mottram, D.S.; Halford, N.G. Effects of water availability on free amino acids, sugars, and acrylamide-forming potential in potato. J. Agric. Food Chem. 2015, 63, 2566–2575. [Google Scholar] [CrossRef]
- Paul, V.; Ezekiel, R.; Pandey, R. Acrylamide in processed potato products: Progress made and present status. Acta Physiol. Plant. 2016, 38, 276. [Google Scholar] [CrossRef]
- Rosen, C.; Sun, N.; Olsen, N.; Thornton, M.; Pavek, M.; Knowles, L.; Knowles, N.R. Impact of agronomic and storage practices on acrylamide in processed potatoes. Am. J. Potato Res. 2018, 95, 319–327. [Google Scholar] [CrossRef]
- Powers, S.J.; Mottram, D.S.; Curtis, A.; Halford, N.G. Acrylamide concentrations in potato crisps in Europe from 2002 to 2011. Food Addit. Contam. A 2013, 30, 1493–1500. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Bethke, P.C.; Bussan, A.J.; Glynn, M.T.; Holm, D.G.; Navarro, F.M.; Novy, R.G.; Palta, J.P.; Pavek, M.J.; Porter, G.A.; et al. Acrylamide-forming potential and agronomic properties of elite us potato germplasm from the national fry processing trial. Crop Sci. 2016, 56, 30–39. [Google Scholar] [CrossRef]
- European Food Safety Authority. Acrylamide in Food is a Public Health Concern. Available online: http://www.efsa.europa.eu/en/press/news/150604 (accessed on 10 January 2020).
- Westermann, D.T.; James, D.W.; Tindall, T.A.; Hurst, R.L. Nitrogen and potassium fertilization of potatoes: Sugars and starch. Am. Potato J. 1994, 71, 433–453. [Google Scholar] [CrossRef]
- De Wilde, T.B.; De Meulenaer, B.; Mestdagh, F.; Govaert, Y.; Vandeburie, S.; Ooghe, W.; Fraselle, S.; Demeulemeester, K.; Van Peteghem, C.; Calus, A.; et al. Influence of fertilization on acrylamide formation during frying of potatoes harvested in 2003. J. Agric. Food Chem. 2006, 54, 404–408. [Google Scholar] [CrossRef]
- Argyropoulos, D.; Psallida, C.; Varzakas, T. The effect of nitrogen fertilisation and metabolic regulators SNRK1, GCN2 on the formation of acrylamide in two potato varieties (Spunta and Lady Rosetta) fried in corn oil. Curr. Res. Nutr. Food Sci. 2016, 4, 69–73. [Google Scholar] [CrossRef]
- Kolbe, H. Kartoffeldüngung Unter Differenzierten ökologischen Bedingungen. Ph.D. Thesis, Georg-August-Universität, Götingen, Germany, 1990. [Google Scholar]
- Morales, F.; Capuano, E.; Fogliano, V. Mitigation strategies to reduce acrylamide formation in fried potato products. Ann. NY Acad. Sci. 2008, 1126, 89–100. [Google Scholar] [CrossRef]
- Knowles, N.R.; Driskill, E.P.; Knowles, L.O. Sweetening responses of potato tubers of different maturity to conventional and non-conventional storage temperature regimes. Postharvest Biol. Technol. 2009, 52, 49–61. [Google Scholar] [CrossRef]
- Elmore, J.S.; Briddon, A.; Dodson, A.T.; Muttucumaru, N.; Halford, N.G.; Mottram, D.S. Acrylamide in potato crisps prepared from 20 UK-grown varieties: Effects of variety and tuber storage time. Food Chem. 2015, 182, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Muttucumaru, N.; Powers, S.J.; Elmore, J.S.; Dodson, A.; Briddon, A.; Mottram, D.S.; Halford, N.G. Acrylamide-forming potential of potatoes grown at different locations, and the ratio of free asparagine to reducing sugars at which free asparagine becomes a limiting factor for acrylamide formation. Food Chem. 2017, 220, 76–86. [Google Scholar] [CrossRef] [Green Version]
- Sun, N.; Rosen, C.; Thompson, A. Acrylamide formation in processed potates as affected by cultivar, nitrogen fertilization and storage time. Am. J. Potato Res. 2018, 95, 473–486. [Google Scholar] [CrossRef]
- De Wilde, T.; De Meulenaer, B.; Mestdagh, F.; Govaert, Y.; Vandeburie, S.; Ooghe, W.; Fraselle, S.; Demeulemeester, K.; van Peteghem, C.; Calus, A.; et al. Influence of storage practices on acrylamide formation during potato frying. J. Agric. Food Chem. 2005, 53, 6550–6557. [Google Scholar] [CrossRef] [PubMed]
- Ohara-Takada, A.; Matsuura-Endo, C.; Chuda, Y.; Ono, H.; Yada, H.; Yoshida, M.; Kobayashi, A.; Tsuda, S.; Takigawa, S.; Noda, T.; et al. Change in content of sugars and free amino acids in potato tubers under short-term storage at low temperature and the effect on acrylamide level after frying. Biosci. Biotechnol. Biochem. 2005, 69, 1232–1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavek, M.J.; Knowles, N.R. WSU Potato Cultivar Yield and Postharvest Quality Evaluations for 2015. Washington State University Special Report, 114. Available online: http://potatoes.wsu.edu/wp-content/uploads/2016/01/Potato-Cultivar-Yield-and-Postharvest-Quality-Evaluations-Research-Edition-2015.pdf. (accessed on 10 January 2020).
- Olsson, K.; Svensson, R.; Roslund, C.A. Tuber components affecting acrylamide formation and colour in fried potato: Variation by variety, year, storage temperature and storage time. J. Sci. Food Agric. 2004, 84, 447–458. [Google Scholar] [CrossRef]
- Matsuura-Endo, C.; Ohara-Takada, A.; Chuda, Y.; Ono, H.; Yada, H.; Yoshida, M.; Kobayashi, A.; Tsuda, S.; Takigawa, S.; Noda, T.; et al. Effects of storage temperature on the contents of sugars and free amino acids in tubers from different potato cultivars and acrylamide in chips. Biosci. Biotechnol. Biochem. 2006, 70, 1173–1180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halford, N.G.; Curtis, T.Y.; Muttucumaru, N.; Postles, J.; Elmore, J.S.; Mottram, D.S. The acrylamide problem: A plant and agronomic science issue. J. Exp. Bot. 2012, 63, 2841–2851. [Google Scholar] [CrossRef] [Green Version]
- Amrein, T.M.; Schönbächler, B.; Rohner, F.; Lukac, H.; Schneider, H.; Keiser, A.; Escher, F.; Amadò, R. Potential for acrylamide formation in potatoes: Data from the 2003 harvest. Eur. Food Res. Technol. 2004, 219, 572–578. [Google Scholar] [CrossRef]
- Halford, N.G.; Muttucumaru, N.; Powers, S.J.; Gillatt, P.N.; Hartley, L.; Elmore, J.S.; Mottram, D.S. Concentrations of free amino acids and sugars in nine potato varieties: Effects of storage and relationship with acrylamide formation. J. Agric. Food Chem. 2012, 60, 12044–12055. [Google Scholar] [CrossRef]
- Zhu, F.; Cai, Y.Z.; Ke, J.; Corke, H. Compositions of phenolic compounds, amino acids and reducing sugars in commercial potato varieties and their effects on acrylamide formation. J. Sci. Food Agric. 2010, 90, 2254–2262. [Google Scholar] [CrossRef]
- Shepherd, L.V.T.; Bradshaw, J.E.; Dale, M.F.B.; McNicol, J.W.; Pont, S.D.A.; Mottram, D.S.; Davies, H.V. Variation in acrylamide producing potential in potato: Segregation of the trait in a breeding population. Food Chem. 2010, 123, 568–573. [Google Scholar] [CrossRef]
- Muttucumaru, N.; Powers, S.J.; Elmore, J.S.; Briddon, A.; Mottram, D.S.; Halford, N.G. Evidence for the complex relationship between free amino acid and sugar concentrations and acrylamide-forming potential in potato. Ann. Appl. Biol. 2014, 164, 286–300. [Google Scholar] [CrossRef] [PubMed]
- Sun, N.; Wang, Y.; Gupta, S.K.; Rosen, C.J. Nitrogen fertility and cultivar effects on potato agronomic properties and acrylamide-forming potential. Agron. J. 2019, 3, 408–418. [Google Scholar] [CrossRef]
- Ryan, S.E. A Tutorial on the Piecewise Regression Approach Applied to Bedload Transport Data. USDA Forest Service—General Technical Report RMRS-GTR-189. Available online: https://www.fs.usda.gov/rds/archive/Product/RDS-2007-0004/ (accessed on 10 January 2020).
- Silva, E.M.; Simon, P.W. Genetic, physiological, and environmental factors affecting acrylamide concentration in fried potato products. In Chemistry and Safety of Acrylamide in Food; Friedman, M., Mottram, D., Eds.; Springer: Boston, MA, USA, 2005; Volume 561, pp. 371–386. [Google Scholar]
- Long, C.M.; Snapp, S.S.; Douches, D.S.; Chase, R.W. Tuber yield, storability, and quality of Michigan cultivars in response to nitrogen management and seed piece spacing. Am. J. Potato Res. 2004, 81, 347–357. [Google Scholar] [CrossRef]
- Zebarth, B.J.; Leclerc, Y.; Moreau, G.; Botha, E. Rate and timing of nitrogen fertilization of Russet Burbank potato: Yield and processing quality. Can. J. Plant Sci. 2004, 84, 855–863. [Google Scholar] [CrossRef]
- Gerendás, J.; Heuser, F.; Sattelmacher, B. Influence of nitrogen and potassium supply on contents of acrylamide precursors in potato tubers and on acrylamide accumulation in French fries. J. Plant Nutr. 2007, 30, 1499–1516. [Google Scholar] [CrossRef]
- Knowles, N.R.; Pavek, M.J.; Knowles, L.O. Developmental profiles, nitrogen use and postharvest quality of alpine and sage russet potatoes in the columbia basin. In Proceedings of the Annual Washington and Oregon Potato Conference, Kennwick, WA, USA, 27–30 January 2015; pp. 37–50. Available online: http://www.nwpotatoresearch.com/IPMStuff/PDFs/Proceedings2015.pdf. (accessed on 10 January 2020).
- Gause, K. Effect of Nitrogen and Potassium on Potato Yield, Quality and Acrylamide-Forming Potential. Master’s Thesis, University Maine, Orono, ME, USA, 2014. [Google Scholar]
- North Dakota State University Research Foundation. Available online: http://www.ndsuresearchfoundation.org/dakota_russet (accessed on 10 January 2020).
- Porter, G.; Alyokhin, A.; Lambert, D.; Halseth, D.; Yencho, G.; Jemison, J.; Freeman, J.; Perry, K.; Perkins, L.B.; Zotarelli, L.; et al. Potato Breeding and Variety Development for Improved Quality and Pest Resistance in the Eastern United States; Grant reports; University of Maine office of research and sponsored programs: Orono, ME, USA; Available online: http://digitalcommons.library.umaine.edu/orsp_reports/31/. (accessed on 10 January 2020).
- Shock, C.C.; Stieber, T.D.; Zalewski, J.C.; Eldredge, E.P.; Lewis, M.D. Potato tuber stem-end fry color determination. Am. J. Potato Res. 1994, 71, 77–88. [Google Scholar] [CrossRef]
- Driskill, E.P.; Knowles, L.O.; Knowles, N.R. Temperature-induced changes in potato processing quality during storage are modulated by tuber maturity. Am. J. Potato Res. 2007, 84, 367–383. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variance | Glucose During 32 Weeks of Storage | |
|---|---|---|
| Stem End | Bud End | |
| Main Effect | ||
| Cultivar (C) | <0.0001 | <0.0001 |
| N Rate (N) | 0.0718 | 0.4334 |
| Year (Y) | 0.1461 | 0.0018 |
| Storage Time (S) | <0.0001 | 0.0765 |
| Interactions | ||
| N ×Y | 0.3955 | 0.0142 |
| N × S | 0.3319 | 0.7266 |
| S × Y | <0.0001 | 0.0052 |
| C × Y | <0.0001 | 0.0049 |
| C × N | 0.0033 | 0.8609 |
| C × S | <0.0001 | 0.2078 |
| N × S × Y | 0.5359 | 0.2544 |
| C × N × Y | 0.4159 | 0.1309 |
| C × N × S | 0.0562 | 0.6043 |
| C × S × Y | 0.0295 | 0.2783 |
| C × N × S × Y | 0.4247 | 0.6849 |
| Source | Whole-Tuber | ||
|---|---|---|---|
| Glucose (16 Weeks) | Asparagine (After Vine Kill) | Acrylamide (16 Weeks) | |
| Main Effect | |||
| Cultivar (C) | <0.0001 | <0.0001 | <0.0001 |
| N Rate (N) | 0.3042 | 0.0002 | <0.0001 |
| Year (Y) | 0.3065 | <0.0001 | 0.0052 |
| Interactions | |||
| C × N | 0.1617 | 0.7709 | 0.4667 |
| C × Y | <0.0001 | 0.3661 | 0.0049 |
| N × Y | 0.4420 | 0.4715 | 0.8615 |
| C × N × Y | 0.7170 | 0.9011 | 0.4493 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, N.; Wang, Y.; Gupta, S.K.; Rosen, C.J. Potato Tuber Chemical Properties in Storage as Affected by Cultivar and Nitrogen Rate: Implications for Acrylamide Formation. Foods 2020, 9, 352. https://doi.org/10.3390/foods9030352
Sun N, Wang Y, Gupta SK, Rosen CJ. Potato Tuber Chemical Properties in Storage as Affected by Cultivar and Nitrogen Rate: Implications for Acrylamide Formation. Foods. 2020; 9(3):352. https://doi.org/10.3390/foods9030352
Chicago/Turabian StyleSun, Na, Yi Wang, Sanjay K. Gupta, and Carl J. Rosen. 2020. "Potato Tuber Chemical Properties in Storage as Affected by Cultivar and Nitrogen Rate: Implications for Acrylamide Formation" Foods 9, no. 3: 352. https://doi.org/10.3390/foods9030352
APA StyleSun, N., Wang, Y., Gupta, S. K., & Rosen, C. J. (2020). Potato Tuber Chemical Properties in Storage as Affected by Cultivar and Nitrogen Rate: Implications for Acrylamide Formation. Foods, 9(3), 352. https://doi.org/10.3390/foods9030352