Chemical and Cellular Antioxidant Activities of In Vitro Digesta of Tilapia Protein and Its Hydrolysates



Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Preparation of Protein Hydrolysates
2.3. Simulated In Vitro GI Digestion
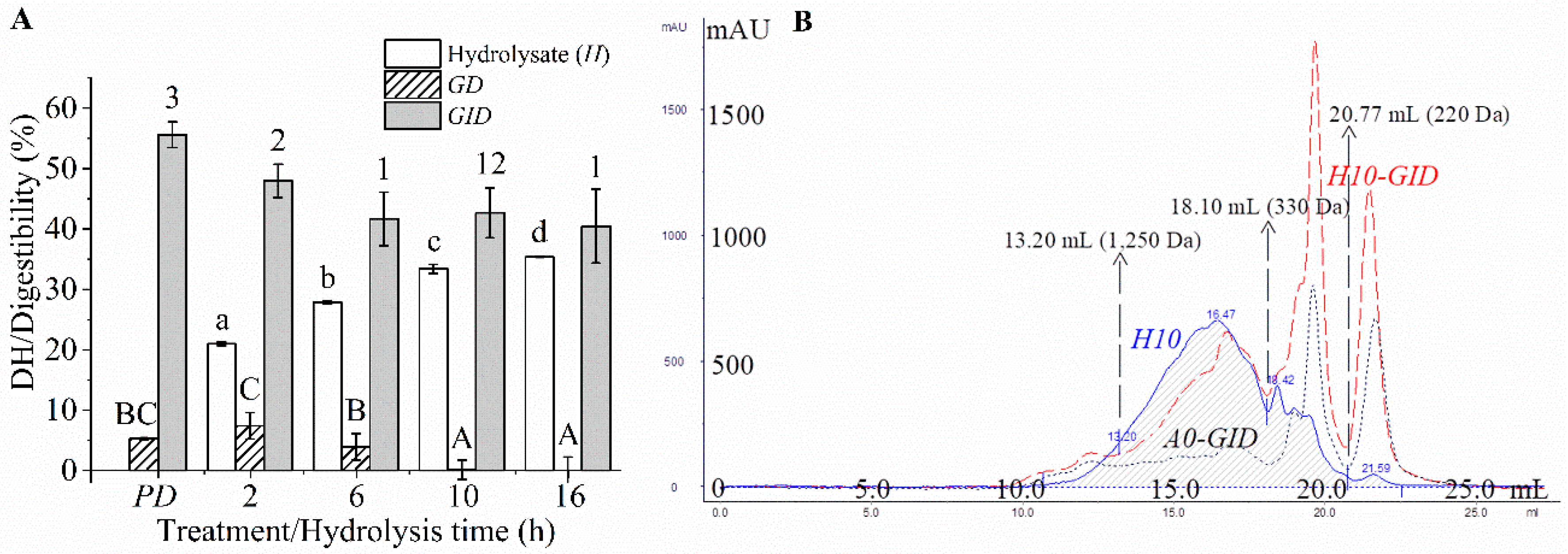
2.4. DH and Digestibility
2.5. Molecular Weight (MW) Distribution
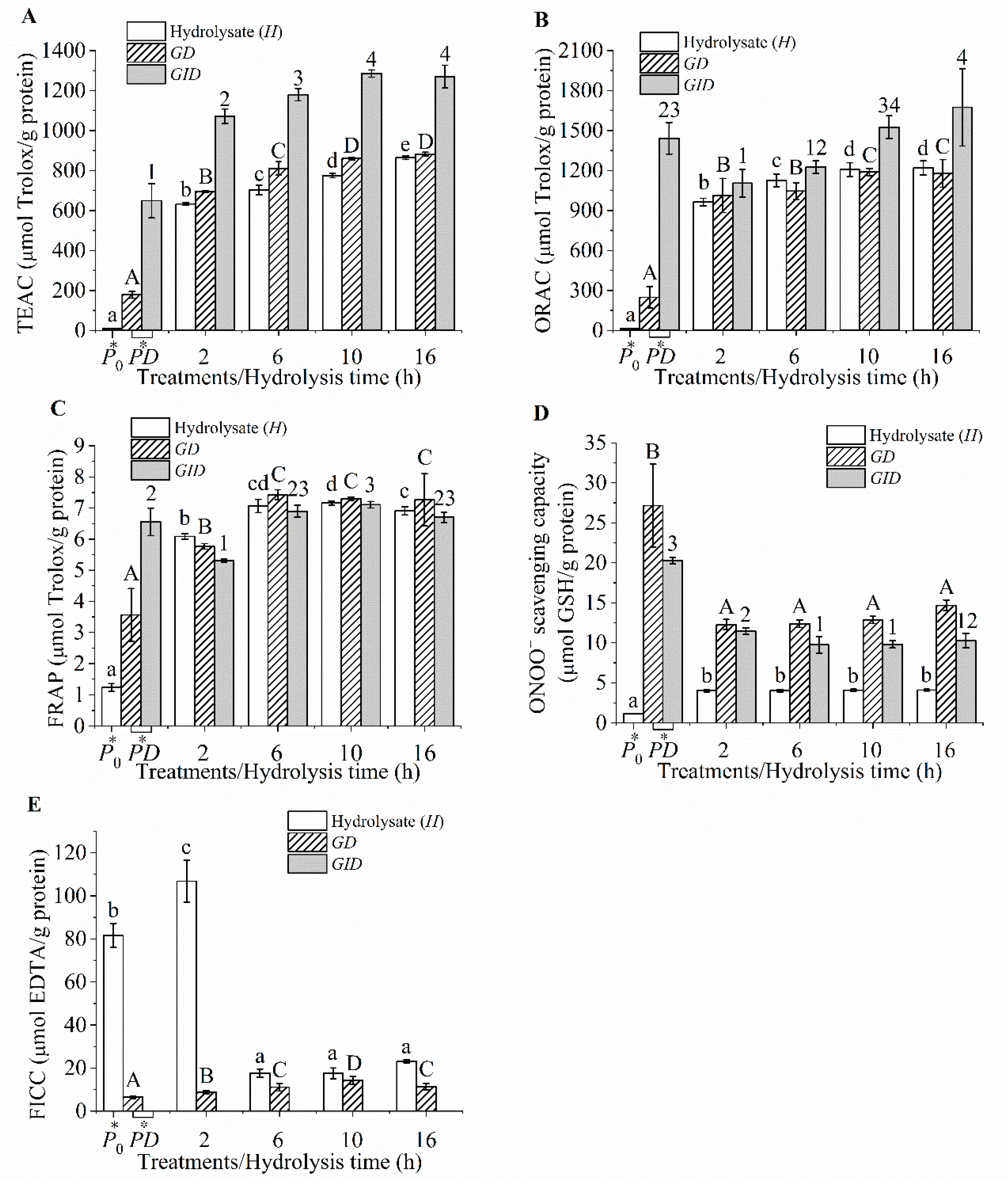
2.6. Chemical Antioxidant Activity
2.6.1. Trolox Equivalent Antioxidant Capacity (TEAC)
2.6.2. Ferric Reducing Antioxidant Power (FRAP)
2.6.3. Fe2+ Chelating Capacity (FICC)
2.6.4. Oxygen Radical Absorbance Capacity (ORAC)
2.6.5. Peroxynitrite () Scavenging
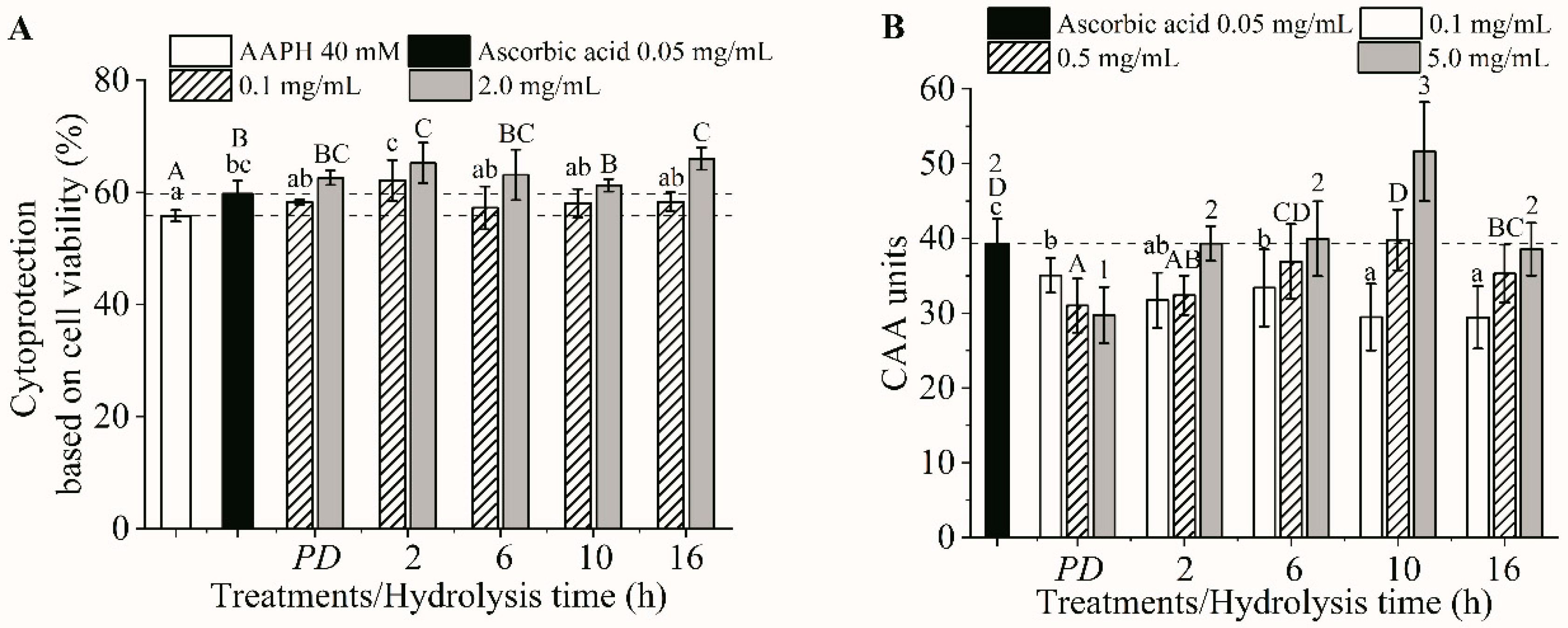
2.7. Cellular Antioxidant Activities
2.7.1. Cell Culture
2.7.2. Cytotoxicity and Cytoprotection
2.7.3. Intracellular ROS Scavenging Capacity
2.8. Statistical Analyses
3. Results and Discussion
3.1. Antioxidant Activities of the Enzymes Used in Hydrolysis and In Vitro GI Digestion
3.2. DH and MW Distribution
3.3. Chemical Antioxidant Activities
3.4. Cellular Antioxidant Activities
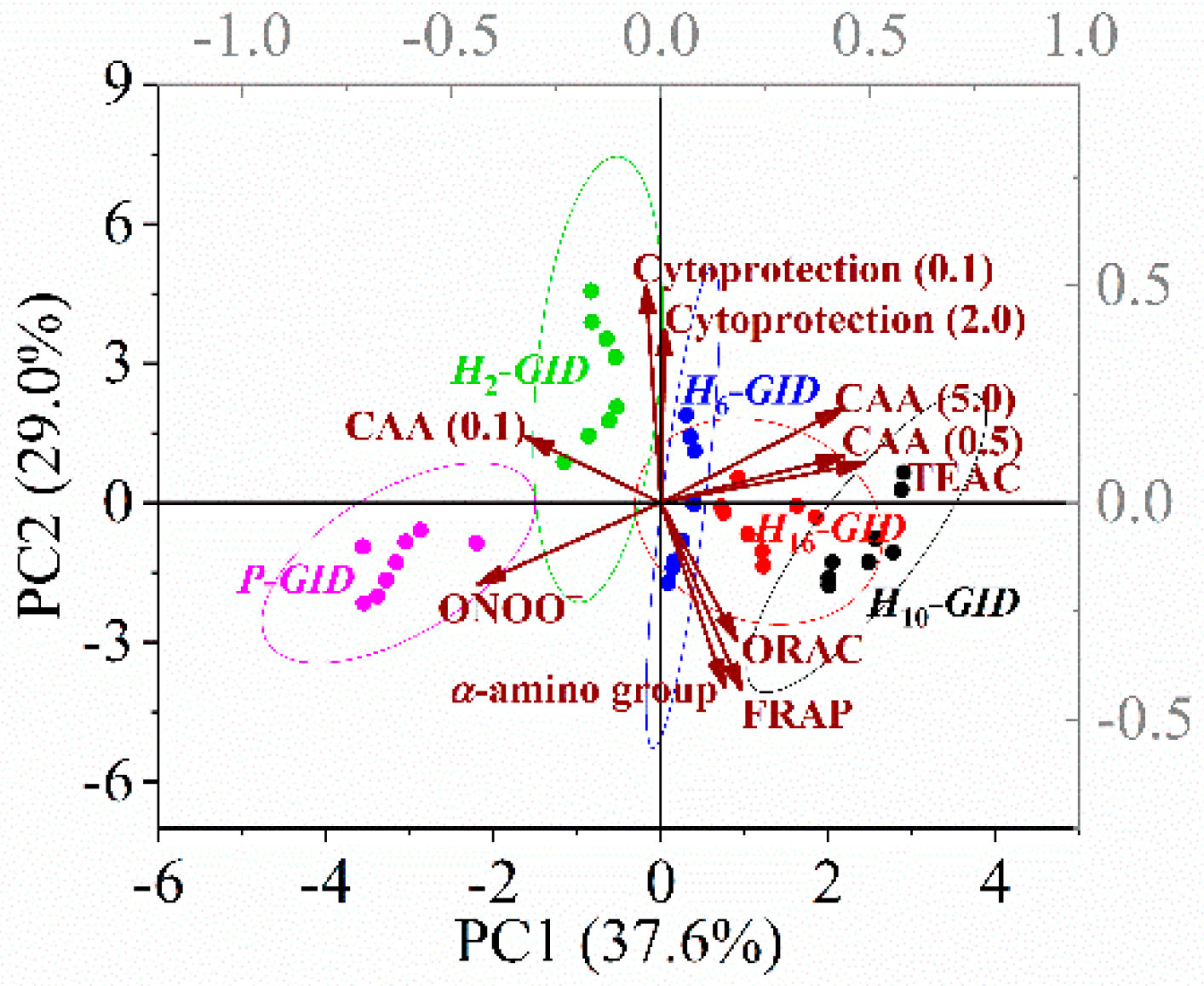
3.5. PCA
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wiriyaphan, C.; Xiao, H.; Decker, E.A.; Yongsawatdigul, J. Chemical and cellular antioxidative properties of threadfin bream (Nemipterus spp.) surimi byproduct hydrolysates fractionated by ultrafiltration. Food Chem. 2015, 167, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Saito, K.; Jin, D.-H.; Ogawa, T.; Muramoto, K.; Hatakeyama, E.; Yasuhara, T.; Nokihara, K. Antioxidative properties of tripeptide libraries prepared by the combinatorial chemistry. J. Agric. Food Chem. 2003, 51, 3668–3674. [Google Scholar] [CrossRef] [PubMed]
- Samaranayaka, A.G.P.; Kitts, D.D.; Li-Chan, E.C.Y. Antioxidative and angiotensin-I-converting enzyme inhibitory potential of a Pacific hake (Merluccius productus) fish protein hydrolysate subjected to simulated gastrointestinal digestion and Caco-2 cell permeation. J. Agric. Food Chem. 2010, 58, 1535–1542. [Google Scholar] [CrossRef] [PubMed]
- Nwachukwu, I.D.; Aluko, R.E. Structural and functional properties of food protein-derived antioxidant peptides. J. Food BioChem. 2019, 43, e12761. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Li, Q.; Hong, H.; Luo, Y. Prevention of protein oxidation and enhancement of gel properties of silver carp (Hypophthalmichthys molitrix) surimi by addition of protein hydrolysates derived from surimi processing by-products. Food Chem. 2020, 316, 126343. [Google Scholar] [CrossRef]
- Wang, J.; Lu, S.; Guo, X.; Li, R.; Huang, L. Effect of crude peptide extract from mutton ham on antioxidant properties and quality of mutton patties. J. Food Process. Preserv. 2019, e14436. [Google Scholar] [CrossRef]
- Samaranayaka, A.G.P.; Li-Chan, E.C.Y. Food-derived peptidic antioxidants: A review of their production, assessment, and potential applications. J. Funct. Foods 2011, 3, 229–254. [Google Scholar] [CrossRef]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020. [Google Scholar] [CrossRef]
- Hou, Y.; Wu, Z.; Dai, Z.; Wang, G.; Wu, G. Protein hydrolysates in animal nutrition: Industrial production, bioactive peptides, and functional significance. J. Anim. Sci. Biotechnol. 2017, 8, 24. [Google Scholar] [CrossRef] [Green Version]
- Zheng, K.; Liang, M.; Yao, H.; Wang, J.; Chang, Q. Effect of size-fractionated fish protein hydrolysate on growth and feed utilization of turbot (Scophthalmus maximus L.). Aquac. Res. 2013, 44, 895–902. [Google Scholar] [CrossRef]
- Zheng, K.; Liang, M.; Yao, H.; Wang, J.; Chang, Q. Effect of dietary fish protein hydrolysate on growth, feed utilization and IGF-I levels of Japanese flounder (Paralichthys olivaceus). Aquac. Nutr. 2012, 18, 297–303. [Google Scholar] [CrossRef]
- Jang, H.L.; Liceaga, A.M.; Yoon, K.Y. Purification, characterisation and stability of an antioxidant peptide derived from sandfish (Arctoscopus japonicus) protein hydrolysates. J. Funct. Foods 2016, 20, 433–442. [Google Scholar] [CrossRef]
- Chakrabarti, S.; Guha, S.; Majumder, K. Food-derived bioactive peptides in human health: Challenges and opportunities. Nutrients 2018, 10, 1738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ketnawa, S.; Martínez-Alvarez, O.; Benjakul, S.; Rawdkuen, S. Gelatin hydrolysates from farmed Giant catfish skin using alkaline proteases and its antioxidative function of simulated gastro-intestinal digestion. Food Chem. 2016, 192, 34–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ketnawa, S.; Wickramathilaka, M.; Liceaga, A.M. Changes on antioxidant activity of microwave-treated protein hydrolysates after simulated gastrointestinal digestion: Purification and identification. Food Chem. 2018, 254, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Ao, J.; Li, B. Stability and antioxidative activities of casein peptide fractions during simulated gastrointestinal digestion in vitro: Charge properties of peptides affect digestive stability. Food Res. Int. 2013, 52, 334–341. [Google Scholar] [CrossRef]
- Yarnpakdee, S.; Benjakul, S.; Kristinsson, H.G.; Bakken, H.E. Preventive effect of Nile tilapia hydrolysate against oxidative damage of HepG2 cells and DNA mediated by H2O2 and AAPH. J. Food Sci. Technol. 2015, 52, 6194–6205. [Google Scholar] [CrossRef] [Green Version]
- Raghavan, S.; Kristinsson, H.G.; Leeuwenburgh, C. Radical scavenging and reducing ability of tilapia (Oreochromis niloticus) protein hydrolysates. J. Agric. Food Chem. 2008, 56, 10359–10367. [Google Scholar] [CrossRef]
- Zhang, X.; Cao, D.; Sun, X.; Sun, S.; Xu, N. Preparation and identification of antioxidant peptides from protein hydrolysate of marine alga Gracilariopsis lemaneiformis. J. Appl. Phycol. 2019, 31, 2585–2596. [Google Scholar] [CrossRef]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019. [Google Scholar] [CrossRef]
- Mulet Cabero, A.; Egger, L.; Portmann, R.; Menard, O.; Marze, S.; Le Feunteun, S.; Sarkar, A.; Grundy, M.; Carrière, F.; Golding, M.; et al. A standardised semi-dynamic in vitro digestion method suitable for food—An international consensus. Food Funct. 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerra, A.; Etienne-Mesmin, L.; Livrelli, V.; Denis, S.; Blanquet-Diot, S.; Alric, M. Relevance and challenges in modeling human gastric and small intestinal digestion. Trends Biotechnol. 2012, 30, 591–600. [Google Scholar] [CrossRef] [PubMed]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food—An international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egger, L.; Schlegel, P.; Baumann, C.; Stoffers, H.; Guggisberg, D.; Brügger, C.; Dürr, D.; Stoll, P.; Vergères, G.; Portmann, R. Physiological comparability of the harmonized INFOGEST in vitro digestion method to in vivo pig digestion. Food Res. Int. 2017, 102, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Dave, L.A.; Hayes, M.; Mora, L.; Montoya, C.A.; Moughan, P.J.; Rutherfurd, S.M. Gastrointestinal endogenous protein-derived bioactive peptides: An in vitro study of their gut modulatory potential. Int. J. Mol. Sci. 2016, 17, 482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitcomb, D.C.; Lowe, M.E. Human pancreatic digestive enzymes. Digestive Dis. Sci. 2007, 52, 1–17. [Google Scholar] [CrossRef]
- López-Alarcón, C.; Denicola, A. Evaluating the antioxidant capacity of natural products: A review on chemical and cellular-based assays. Anal. Chim. Acta 2013, 763, 1–10. [Google Scholar] [CrossRef]
- Wolfe, K.L.; Liu, R.H. Cellular antioxidant activity (CAA) assay for assessing antioxidants, foods, and dietary supplements. J. Agric. Food Chem. 2007, 55, 8896–8907. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 17th ed.; Association of Official Chemists: Gaithersburg, MD, USA, 2000. [Google Scholar]
- Adler-Nissen, J. Determination of the degree of hydrolysis of food protein hydrolysates by trinitrobenzenesulfonic acid. J. Agric. Food Chem. 1979, 27, 1256–1262. [Google Scholar] [CrossRef]
- Benjakul, S.; Morrissey, M.T. Protein hydrolysates from Pacific whiting solid wastes. J. Agric. Food Chem. 1997, 45, 3423–3430. [Google Scholar] [CrossRef]
- Wu, G.; Mansy, S.S.; Wu, S.-p.; Surerus, K.K.; Foster, M.W.; Cowan, J.A. Characterization of an iron-sulfur cluster assembly protein (ISU1) from Schizosaccharomyces pombe. Biochemistry 2002, 41, 5024–5032. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. BioChem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decker, E.A.; Welch, B. Role of ferritin as a lipid oxidation catalyst in muscle food. J. Agric. Food Chem. 1990, 38, 674–677. [Google Scholar] [CrossRef]
- Dávalos, A.; Gómez-Cordovés, C.; Bartolomé, B. Extending applicability of the oxygen radical absorbance capacity (ORAC-fluorescein) assay. J. Agric. Food Chem. 2004, 52, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Kooy, N.W.; Royall, J.A.; Ischiropoulos, H.; Beckman, J.S. Peroxynitrite-mediated oxidation of dihydrorhodamine 123. Free Radic. Biol. Med. 1994, 16, 149–156. [Google Scholar] [CrossRef]
- Kellett, M.E.; Greenspan, P.; Pegg, R.B. Modification of the cellular antioxidant activity (CAA) assay to study phenolic antioxidants in a Caco-2 cell line. Food Chem. 2018, 244, 359–363. [Google Scholar] [CrossRef]
- Yarnpakdee, S.; Benjakul, S.; Kristinsson, H.G.; Kishimura, H. Antioxidant and sensory properties of protein hydrolysate derived from Nile tilapia (Oreochromis niloticus) by one-and two-step hydrolysis. J. Food Sci. Technol. 2015, 52, 3336–3349. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Li, B.; Wang, B.; Xie, N. Degradation and antioxidant activities of peptides and zinc–peptide complexes during in vitro gastrointestinal digestion. Food Chem. 2015, 173, 733–740. [Google Scholar] [CrossRef]
- Keil, B. Data treatment. In Specificity of Proteolysis; Keil, B., Ed.; Springer: Berlin, Germany, 1992; pp. 7–18. [Google Scholar]
- Ketnawa, S.; Benjakul, S.; Martínez-Alvarez, O.; Rawdkuen, S. Fish skin gelatin hydrolysates produced by visceral peptidase and bovine trypsin: Bioactivity and stability. Food Chem. 2017, 215, 383–390. [Google Scholar] [CrossRef]
- Apak, R.; Özyürek, M.; Güçlü, K.; Çapanoğlu, E. Antioxidant activity/capacity measurement. 2. Hydrogen atom transfer (HAT)-based, mixed-mode (electron transfer (ET)/HAT), and lipid peroxidation assays. J. Agric. Food Chem. 2016, 64, 1028–1045. [Google Scholar] [CrossRef] [PubMed]
- Aluko, R.E. Amino acids, peptides, and proteins as antioxidants for food preservation. In Handbook of Antioxidants for Food Preservation; Shahidi, F., Ed.; Woodhead Publishing: Sawston, Cambridge, UK, 2015; pp. 105–140. [Google Scholar]
- Balavoine, G.G.A.; Geletii, Y.V. Peroxynitrite scavenging by different antioxidants. Part I: Convenient assay. Nitric Oxide 1999, 3, 40–54. [Google Scholar] [CrossRef] [PubMed]
- Adeyeye, E.I. Amino acid composition of three species of Nigerian fish: Clarias anguillaris, Oreochromis niloticus and Cynoglossus senegalensis. Food Chem. 2009, 113, 43–46. [Google Scholar] [CrossRef]
- You, L.; Zhao, M.; Regenstein, J.M.; Ren, J. Changes in the antioxidant activity of loach (Misgurnus anguillicaudatus) protein hydrolysates during a simulated gastrointestinal digestion. Food Chem. 2010, 120, 810–816. [Google Scholar] [CrossRef]
- Mirzaei, M.; Aminlari, M.; Hosseini, E. Antioxidant, ACE-inhibitory and antimicrobial activities of Kluyveromyces marxianus protein hydrolysates and their peptide fractions. Funct. Foods Health Dis. 2016, 6, 425–439. [Google Scholar] [CrossRef]
- Du, Y.; Esfandi, R.; Willmore, W.G.; Tsopmo, A. Antioxidant activity of oat proteins derived peptides in stressed hepatic HepG2 cells. Antioxidants 2016, 5, 39. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | A0 | Pep-I | Pep-Pan-I | A0-GID | H10-GID# | |
|---|---|---|---|---|---|---|
| Parameters | ||||||
| α-Amino (mmol L-Leu) | Free | 0.02 ± 0.00 a | 0.01 ± 0.00 a | 1.05 ± 0.01 b | 1.02 ± 0.02 b | 2.69 ± 0.09 c |
| Total | – | – | – | 2.00 ± 0.15 a | 4.73 ± 0.12 b | |
| TEAC (μmol Trolox) | 12.18 ± 0.98 a | 39.02 ± 0.00 b | 389.54 ± 0.55 c | 487.47 ± 0.65 d | 1004.83 ± 7.35 e | |
| FRAP (μmol Trolox) | 0.11 ± 0.08 a | 0.03 ± 0.00 a | 1.94 ± 0.04 c | 1.78 ± 0.04 b | 4.64 ± 0.04 d | |
| FICC (μmol EDTA) | 5.99 ± 0.40 a | ND | 7.33 ± 0.68 ab | 11.59 ± 2.11 c | 8.25 ± 0.35 b | |
| ORAC (μmol Trolox) | 12.39 ± 1.09 a | 0.57 ± 0.03 a | 235.69 ± 58.40 b | 370.63 ± 10.35 c | 984.54 ± 34.18 d | |
| (μmol GSH) | ND | ND | 0.59 ± 0.05 a | 1.06 ± 0.16 b | 3.95 ± 0.19 c | |
| Sample | Percentage of Area under Chromatogram | |||
|---|---|---|---|---|
| >1250 Da | 1250–330 Da | 330–220 Da | <220 Da | |
| Protein hydrolysates | ||||
| H2 | 12.29 ± 1.31 c | 77.49 ± 1.67 b | 10.00 ± 0.36 a | 0.22 ± 0.12 a |
| H6 | 7.60 ± 0.68 b | 75.64 ± 1.88 ab | 15.59 ± 0.57 b | 1.17 ± 0.76 ab |
| H10 | 4.46 ± 0.30 a | 75.55 ± 0.92 ab | 18.73 ± 0.70 c | 1.25 ± 0.39 ab |
| H16 | 3.61 ± 0.54 a | 73.52 ± 0.34 a | 21.02 ± 0.75 d | 1.85 ± 0.15 b |
| Digesta of protein hydrolysates | ||||
| P-GID | 5.79 ± 0.30 A | 37.50 ± 0.27 BCD | 35.27 ± 0.37 BC | 21.44 ± 0.36 D |
| H2-GID | 6.13 ± 0.11 A | 37.74 ± 0.38 CD | 35.70 ± 0.32 BC | 20.43 ± 0.55 CD |
| H6-GID | 5.71 ± 0.21 A | 38.77 ± 0.61 D | 36.24 ± 0.16 BC | 19.28 ± 0.27 ABC |
| H10-GID | 5.76 ± 0.24 A | 37.59 ± 0.16 BCD | 36.61 ± 0.30 C | 20.04 ± 0.27 BC |
| H16-GID | 6.07 ± 0.45 A | 37.52 ± 0.26 BCD | 36.59 ± 0.46 C | 19.82 ± 0.24 BC |
| Enzyme blanks | ||||
| Pan0 | 16.47 ± 0.34 D | 36.57 ± 0.49 BC | 28.90 ± 0.96 A | 18.06 ± 0.69 A |
| A0-Pep0-Pan0 | 14.96 ± 1.08 C | 36.32 ± 0.11 B | 29.83 ± 1.40 A | 18.90 ± 0.42 AB |
| A0-GID | 8.99 ± 0.43 B | 26.87 ± 0.74 A | 34.57 ± 0.64 B | 29.56 ± 0.71 E |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Noisa, P.; Yongsawatdigul, J. Chemical and Cellular Antioxidant Activities of In Vitro Digesta of Tilapia Protein and Its Hydrolysates. Foods 2020, 9, 833. https://doi.org/10.3390/foods9060833
Zhang X, Noisa P, Yongsawatdigul J. Chemical and Cellular Antioxidant Activities of In Vitro Digesta of Tilapia Protein and Its Hydrolysates. Foods. 2020; 9(6):833. https://doi.org/10.3390/foods9060833
Chicago/Turabian StyleZhang, Xiaogang, Parinya Noisa, and Jirawat Yongsawatdigul. 2020. "Chemical and Cellular Antioxidant Activities of In Vitro Digesta of Tilapia Protein and Its Hydrolysates" Foods 9, no. 6: 833. https://doi.org/10.3390/foods9060833
APA StyleZhang, X., Noisa, P., & Yongsawatdigul, J. (2020). Chemical and Cellular Antioxidant Activities of In Vitro Digesta of Tilapia Protein and Its Hydrolysates. Foods, 9(6), 833. https://doi.org/10.3390/foods9060833