
Toxicity of Orthodontic Brackets Examined by Single Cell Tracking

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.1.1. Orthodontic Brackets and Bracket Sample Coding
2.1.2. Cell Culture Materials and Reagents
2.2. Preparation of Bracket-Conditioned Media
2.3. Metal Analysis of Conditioned Media
2.4. Cell Culture
2.5. Single Cell Tracking
2.6. Statistical Analysis
3. Results
3.1. Metal Ions Were Released from Brackets
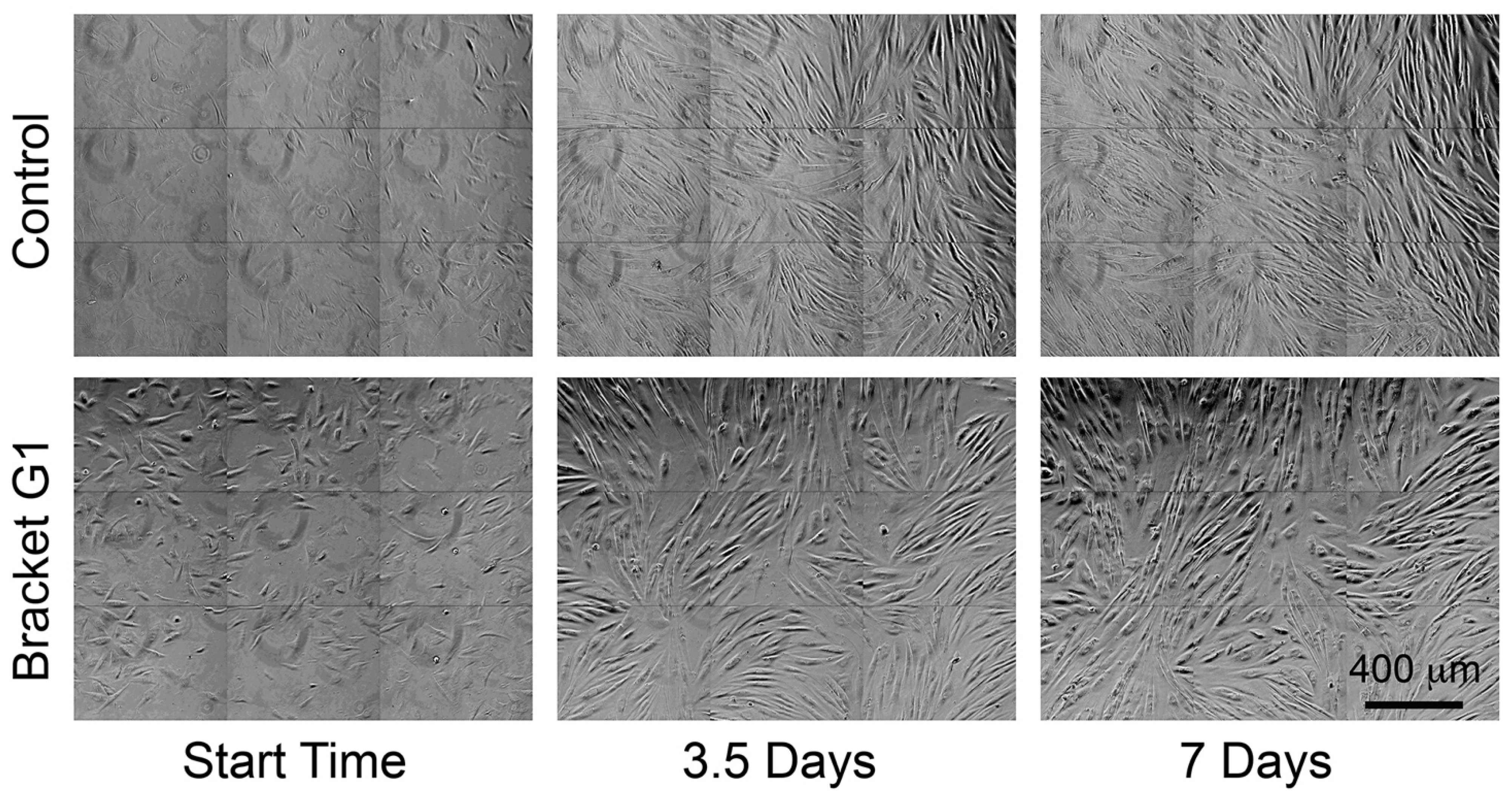
3.2. Conditioned Media from Some Brackets Reduced Fibroblast Proliferation
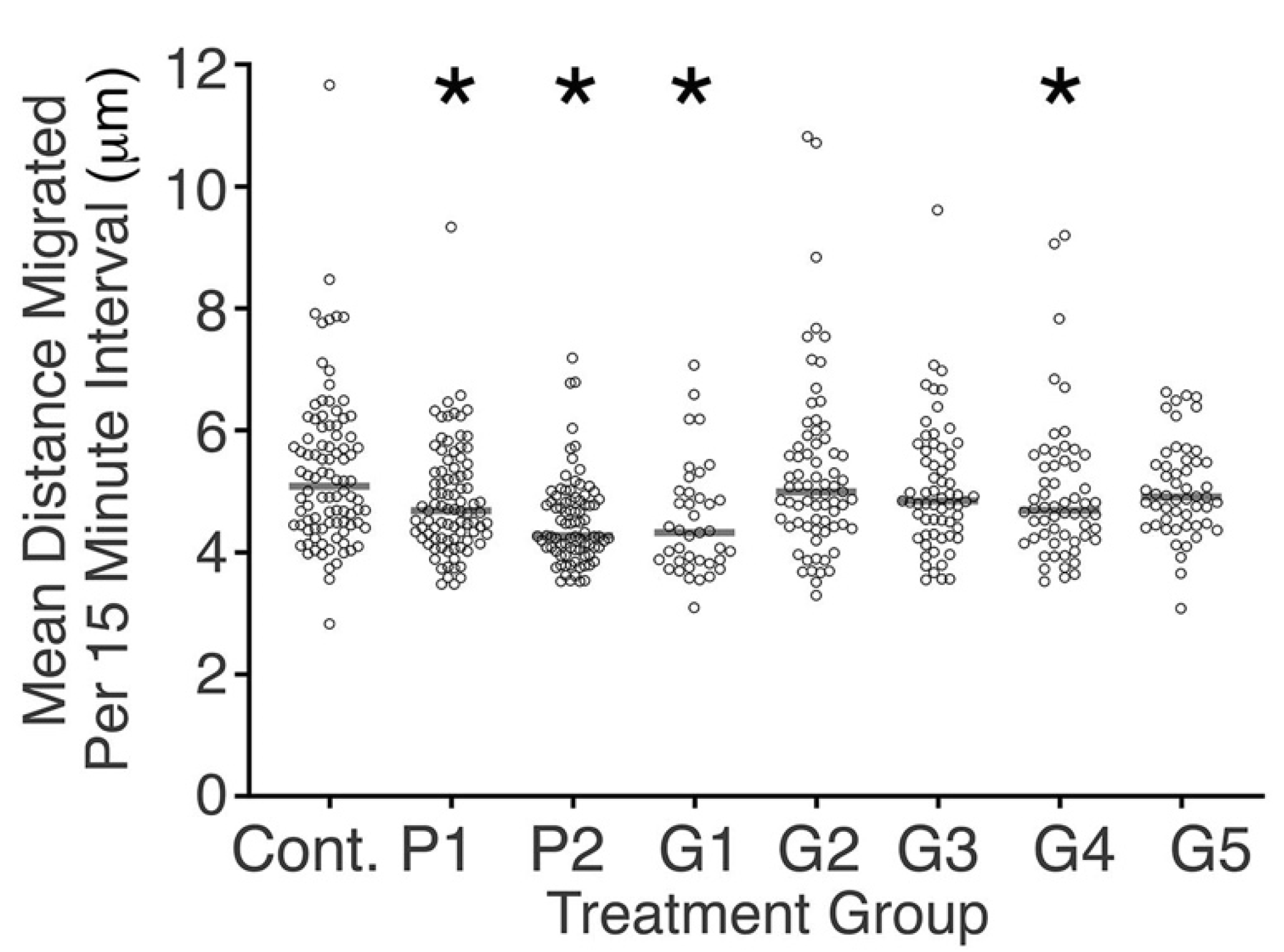
3.3. Some Bracket-Conditioned Media Reduced Fibroblast Migration
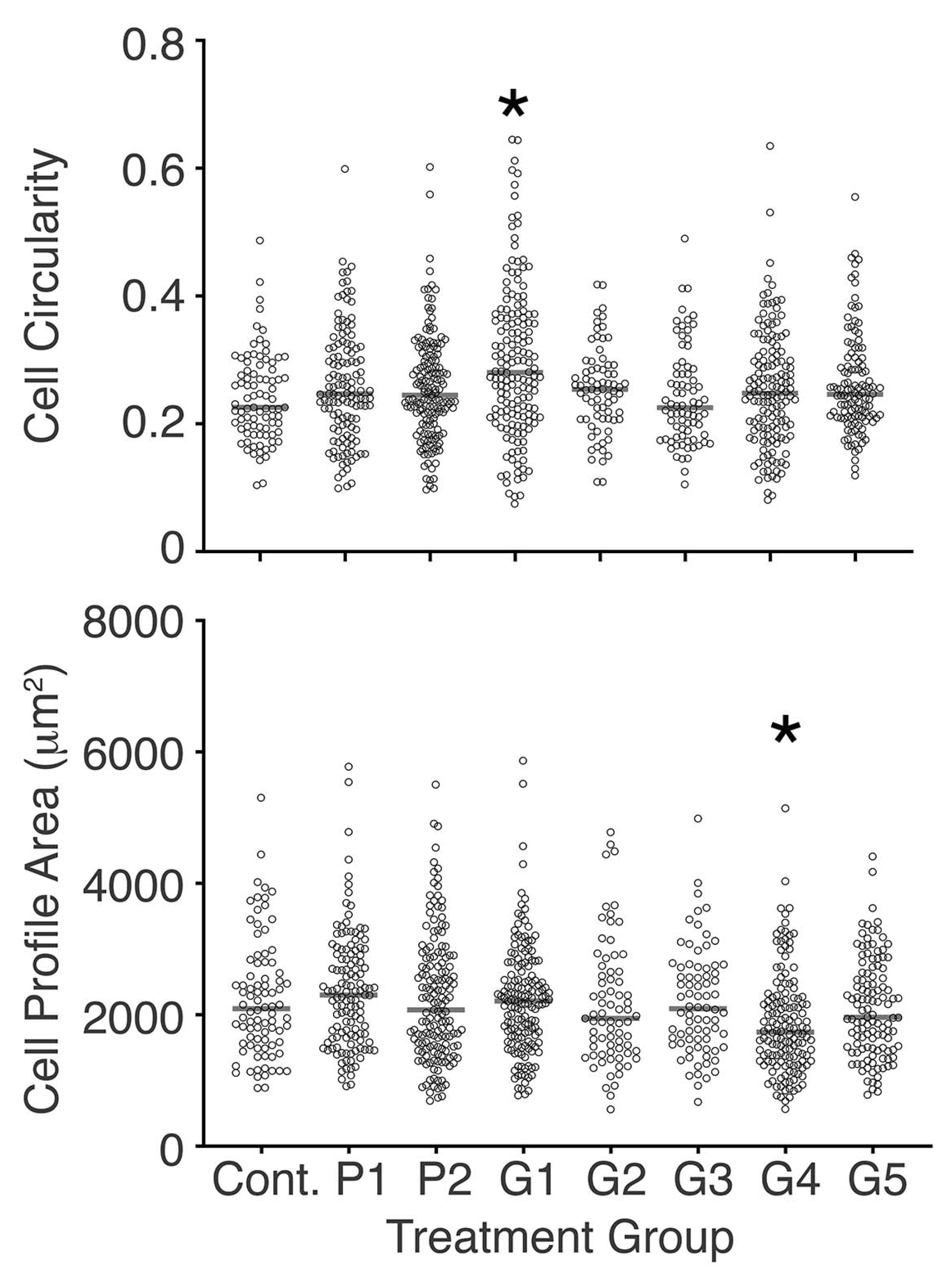
3.4. Bracket-Conditioned Media Had Little Effect on Cell Circularity and Cell Profile Area
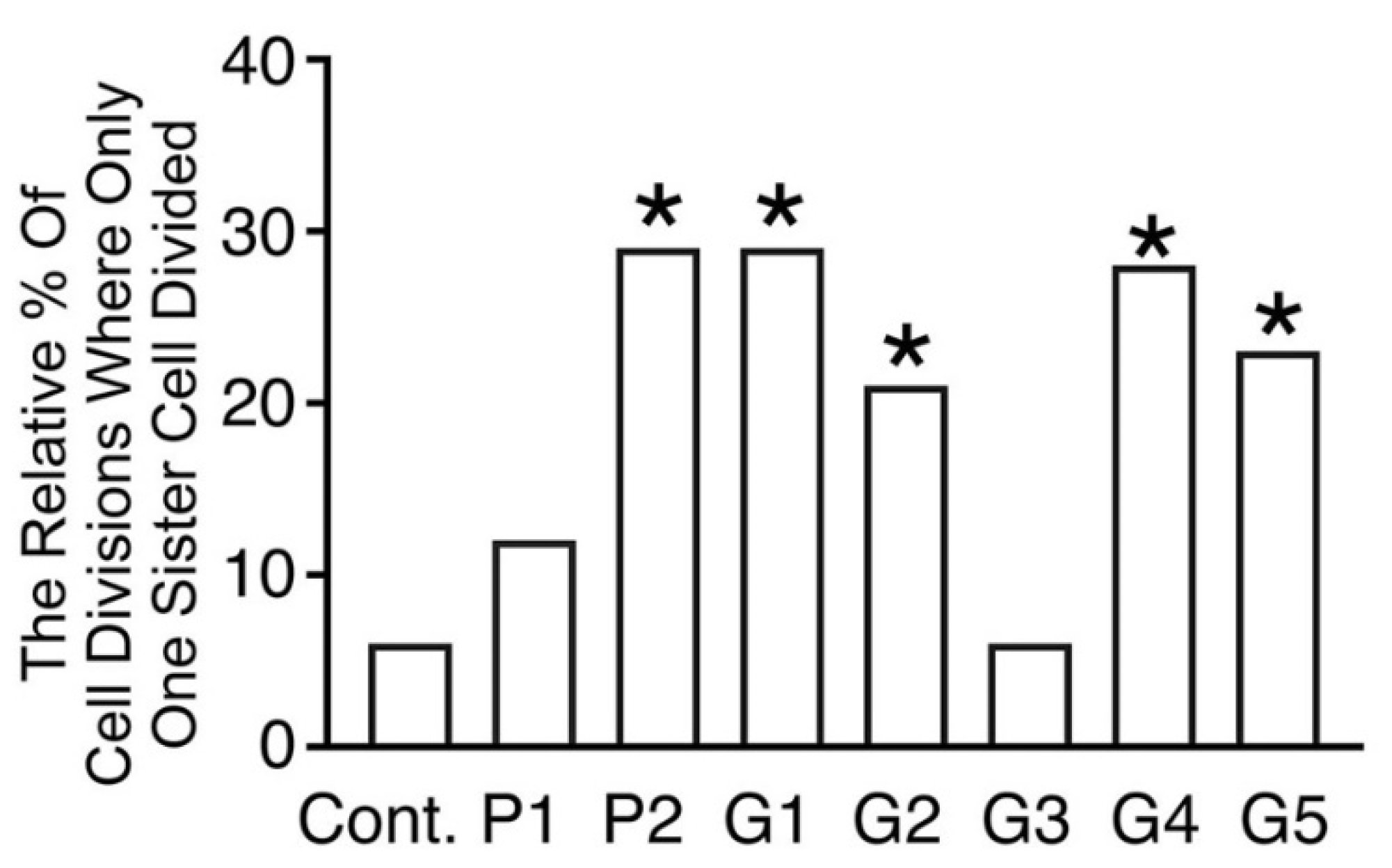
3.5. Loss of Paired Sister Cell Division in Bracket-Conditioned Medium
3.6. Lack of Clear Association between Discrete Metal Contaminants and Cellular Responses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dwivedi, A.; Tikku, T.; Khanna, R.; Maurya, R.P.; Verma, G.; Murthy, R. Release of nickel and chromium ions in the saliva of patients with fixed orthodontic appliance: An in-vivo study. Natl. J. Maxillofac. Surg. 2015, 6, 62. [Google Scholar] [CrossRef] [PubMed]
- Martín-Cameán, A.; Jos, Á.; Mellado-García, P.; Iglesias-Linares, A.; Solano, E.; Cameán, A.M. In vitro and in vivo evidence of the cytotoxic and genotoxic effects of metal ions released by orthodontic appliances: A review. Environ. Toxicol. Pharmacol. 2015, 40, 86–113. [Google Scholar] [CrossRef] [PubMed]
- Ağaoğlu, G.; Arun, T.; İzgü, B.; Yarat, A. Nickel and chromium levels in the saliva and serum of patients with fixed orthodontic appliances. Angle Orthod. 2001, 71, 375–379. [Google Scholar] [PubMed]
- Martín-Cameán, A.; Jos, A.; Cameán, A.M.; Solano, E.; Iglesias-Linares, A. Genotoxic and cytotoxic effects and gene expression changes induced by fixed orthodontic appliances in oral mucosa cells of patients: A systematic review. Toxicol. Mech. Methods 2015, 25, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Martín-Cameán, A.; Molina-Villalba, I.; Jos, A.; Iglesias-Linares, A.; Solano, E.; Cameán, A.M.; Gil, F. Biomonitorization of chromium, copper, iron, manganese and nickel in scalp hair from orthodontic patients by atomic absorption spectrometry. Environ. Toxicol. Pharmacol. 2014, 37, 759–771. [Google Scholar] [CrossRef]
- Menezes, L.M.; Quintão, C.A.; Bolognese, A.M. Urinary excretion levels of nickel in orthodontic patients. Am. J. Orthod. Dentofac. Orthop. 2007, 131, 635–638. [Google Scholar] [CrossRef]
- Amini, F.; Mollaei, M.; Harandi, S.; Rakhshan, V. Effects of fixed orthodontic treatment on hair nickel and chromium levels: A 6-month prospective preliminary study. Biol. Trace Elem. Res. 2015, 164, 12–17. [Google Scholar] [CrossRef]
- Kapadia, J.M.; Agarwal, A.R.; Mishra, S.; Joneja, P.; Yusuf, A.S.; Choudhary, D.S. Cytotoxic and Genotoxic effect on the Buccal Mucosa Cells of Patients Undergoing Fixed Orthodontic Treatment. J. Contemp. Dent. Pract. 2018, 19, 1358–1362. [Google Scholar] [CrossRef]
- Fors, R.; Persson, M. Nickel in dental plaque and saliva in patients with and without orthodontic appliances. Eur. J. Orthod. 2006, 28, 292–297. [Google Scholar] [CrossRef]
- Lages, R.B.; Bridi, E.C.; Pérez, C.A.; Basting, R.T. Salivary levels of nickel, chromium, iron, and copper in patients treated with metal or esthetic fixed orthodontic appliances: A retrospective cohort study. J. Trace Elem. Med. Biol. 2017, 40, 67–71. [Google Scholar] [CrossRef]
- Petoumenou, E.; Arndt, M.; Keilig, L.; Reimann, S.; Hoederath, H.; Eliades, T.; Jäger, A.; Bourauel, C. Nickel concentration in the saliva of patients with nickel-titanium orthodontic appliances. Am. J. Orthod. Dentofac. Orthop. 2009, 135, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Bussan, D.D.; Snaychuk, L.; Bartzas, G.; Douvris, C. Quantification of trace elements in surgical and KN95 face masks widely used during the SARS-COVID-19 pandemic. Sci. Total Environ. 2022, 814, 151924. [Google Scholar] [CrossRef] [PubMed]
- Burke, K. Dentists shown how to bypass rules. In The Sydney Morning Herald; Fairfax Media: Sydney, Australia, 2009. [Google Scholar]
- Wishney, M.; Savage, T.; Proschogo, N.; Petocz, P.; Darendeliler, M. The metal content of generic orthodontic brackets compared with proprietary brackets. Australas. Orthod. J. 2018, 34, 163. [Google Scholar] [CrossRef]
- Wilschefski, S.C.; Baxter, M.R. Inductively Coupled Plasma Mass Spectrometry: Introduction to Analytical Aspects. Clin. Biochem. Rev. 2019, 40, 115–133. [Google Scholar] [CrossRef] [PubMed]
- Romano, F.; Castiblanco, A.; Spadotto, F.; Di Scipio, F.; Malandrino, M.; Berta, G.N.; Aimetti, M. ICP-Mass-Spectrometry Ionic Profile of Whole Saliva in Patients with Untreated and Treated Periodontitis. Biomedicines 2020, 8, 354. [Google Scholar] [CrossRef]
- Amr, M.A.; Helal, A.F.I. Analysis of trace elements in teeth by ICP-MS: Implications for caries. J. Phys. Sci. 2010, 21, 1–12. [Google Scholar]
- Chang, H.H.; Hemberg, M.; Barahona, M.; Ingber, D.E.; Huang, S. Transcriptome-wide noise controls lineage choice in mammalian progenitor cells. Nature 2008, 453, 544. [Google Scholar] [CrossRef] [PubMed]
- Meijering, E.; Dzyubachyk, O.; Smal, I.; van Cappellen, W.A. Tracking in cell and developmental biology. Semin. Cell Dev. Biol. 2009, 20, 894–902. [Google Scholar] [CrossRef]
- Kumar, V.; Abbas, A.K.; Fausto, N.; Robbins, S.L. Robbins and Cotran Pathologic Basis of Disease; Elsevier Saunders: Philadelphia, PA, USA, 2005; p. xv, 1525. [Google Scholar]
- Hoppe, P.S.; Coutu, D.L.; Schroeder, T. Single-cell technologies sharpen up mammalian stem cell research. Nat. Cell Biol. 2014, 16, 919. [Google Scholar] [CrossRef]
- Cornwell, J.; Li, J.; Mahadevan, S.; Draper, J.; Joun, G.; Zoellner, H.; Asli, N.; Harvey, R.; Nordon, R. TrackPad: Software for semi-automated single-cell tracking and lineage annotation. SoftwareX 2020, 11, 100440. [Google Scholar] [CrossRef]
- Mahadevan, S.; Cornwell, J.A.; Chami, B.; Kelly, E.; Zoellner, H. Cell-Projection Pumping of Fibroblast Contents into Osteosarcoma SAOS-2 Cells Correlates with Increased SAOS-2 Proliferation and Migration, as well as Altered Morphology. Biomolecules 2021, 11, 1875. [Google Scholar] [CrossRef]
- Chakrabarti, S.; Paek, A.L.; Reyes, J.; Lasick, K.A.; Lahav, G.; Michor, F. Hidden heterogeneity and circadian-controlled cell fate inferred from single cell lineages. Nat. Commun. 2018, 9, 5372. [Google Scholar] [CrossRef]
- Cornwell, J.A.; Hallett, R.M.; der Mauer, S.A.; Motazedian, A.; Schroeder, T.; Draper, J.; Harvey, R.P.; Nordon, R. Quantifying intrinsic and extrinsic control of single-cell fates in cancer and stem/progenitor cell pedigrees with competing risks analysis. Sci. Rep. 2016, 6, 27100. [Google Scholar] [CrossRef]
- Emmanuel, C.; Huynh, M.; Matthews, J.; Kelly, E.; Zoellner, H. TNF-alpha and TGF-beta synergistically stimulate elongation of human endothelial cells without transdifferentiation to smooth muscle cell phenotype. Cytokine 2013, 61, 38–40. [Google Scholar] [CrossRef]
- David, M.S.; Huynh, M.D.; Kelly, E.; Rizos, H.; Coleman, H.; Rogers, G.; Zoellner, H. Membrane and cytoplasmic marker exchange between malignant neoplastic cells and fibroblasts via intermittent contact: Increased tumour cell diversity independent of genetic change. J. Pathol. 2012, 228, 495–505. [Google Scholar] [CrossRef]
- Korsnes, M.S.; Korsnes, R. Single-Cell Tracking of A549 Lung Cancer Cells Exposed to a Marine Toxin Reveals Correlations in Pedigree Tree Profiles. Front. Oncol. 2018, 8, 260. [Google Scholar] [CrossRef]
- Marcus, J.M.; Burke, R.T.; DeSisto, J.A.; Landesman, Y.; Orth, J.D. Longitudinal tracking of single live cancer cells to understand cell cycle effects of the nuclear export inhibitor, selinexor. Sci. Rep. 2015, 5, 14391. [Google Scholar] [CrossRef]
- Orth, J.D.; Tang, Y.; Shi, J.; Loy, C.T.; Amendt, C.; Wilm, C.; Zenke, F.T.; Mitchison, T.J. Quantitative live imaging of cancer and normal cells treated with Kinesin-5 inhibitors indicates significant differences in phenotypic responses and cell fate. Mol. Cancer Ther. 2008, 7, 3480–3489. [Google Scholar] [CrossRef]
- Gascoigne, K.E.; Taylor, S.S. Cancer cells display profound intra- and interline variation following prolonged exposure to antimitotic drugs. Cancer Cell 2008, 14, 111–122. [Google Scholar] [CrossRef]
- Hallett, R.M.; Huang, C.; Motazedian, A.; Auf der Mauer, S.; Pond, G.R.; Hassell, J.A.; Nordon, R.E.; Draper, J.S. Treatment-induced cell cycle kinetics dictate tumor response to chemotherapy. Oncotarget 2015, 6, 7040–7052. [Google Scholar] [CrossRef]
- Rancourt, A.; Sato, S.; Satoh, M.S. Dose-dependent spatiotemporal responses of mammalian cells to an alkylating agent. PLoS ONE 2019, 14, e0214512. [Google Scholar] [CrossRef]
- Mockers, O.; Deroze, D.; Camps, J. Cytotoxicity of orthodontic bands, brackets and archwires in vitro. Dent. Mater. 2002, 18, 311–317. [Google Scholar] [CrossRef]
- Ortiz, A.J.; Fernández, E.; Vicente, A.; Calvo, J.L.; Ortiz, C. Metallic ions released from stainless steel, nickel-free, and titanium orthodontic alloys: Toxicity and DNA damage. Am. J. Orthod. Dentofac. Orthop. 2011, 140, e115–e122. [Google Scholar] [CrossRef]
- Gonçalves, T.S.; de Menezes, L.M.; Trindade, C.; da Silva Machado, M.; Thomas, P.; Fenech, M.; Henriques, J.A.P. Cytotoxicity and genotoxicity of orthodontic bands with or without silver soldered joints. Mutat. Res. Genet. Toxicol. Environ. Mutagenesis 2014, 762, 1–8. [Google Scholar] [CrossRef]
- Jacoby, L.S.; Junior, V.d.S.R.; Campos, M.M.; de Menezes, L.M. Cytotoxic outcomes of orthodontic bands with and without silver solder in different cell lineages. Am. J. Orthod. Dentofac. Orthop. 2017, 151, 957–963. [Google Scholar] [CrossRef]
- Grimsdottir, M.R.; Gjerdet, N.R.; Hensten-Pettersen, A. Composition and in vitro corrosion of orthodontic appliances. Am. J. Orthod. Dentofac. Orthop. 1992, 101, 525–532. [Google Scholar] [CrossRef]
- Locci, P.; Marinucci, L.; Lilli, C.; Belcastro, S.; Staffolani, N.; Bellocchio, S.; Damiani, F.; Becchetti, E. Biocompatibility of alloys used in orthodontics evaluated by cell culture tests. J. Biomed. Mater. Res. Part A 2000, 51, 561–568. [Google Scholar] [CrossRef]
- Kao, C.-T.; Ding, S.-J.; Min, Y.; Hsu, T.C.; Chou, M.-Y.; Huang, T.-H. The cytotoxicity of orthodontic metal bracket immersion media. Eur. J. Orthod. 2007, 29, 198–203. [Google Scholar] [CrossRef]
- Haddad, A.C.S.S.; Tortamano, A.; Souza, A.L.d.; Oliveira, P.V.d. An in vitro comparison of nickel and chromium release from brackets. Braz. Oral Res. 2009, 23, 399–406. [Google Scholar] [CrossRef]
- Tomakidi, P.; Koke, U.; Kern, R.; Erdinger, L.; Krüger, H.; Kohl, A.; Komposch, G. Assessment of acute cyto-and genotoxicity of corrosion eluates obtained from orthodontic materials using monolayer cultures of immortalized human gingival keratinocytes. J. Orofac. Orthop./Fortschr. Kieferorthopädie 2000, 61, 2–19. [Google Scholar] [CrossRef]
- Wataha, J.C. Biocompatibility of dental casting alloys: A review. J. Prosthet. Dent. 2000, 83, 223–234. [Google Scholar] [CrossRef]
- Craig, R.; Hanks, C. Cytotoxicity of experimental casting alloys evaluated by cell culture tests. J. Dent. Res. 1990, 69, 1539–1542. [Google Scholar] [CrossRef]
- Faccioni, F.; Franceschetti, P.; Cerpelloni, M.; Fracasso, M.E. In vivo study on metal release from fixed orthodontic appliances and DNA damage in oral mucosa cells. Am. J. Orthod. Dentofac. Orthop. 2003, 124, 687–693. [Google Scholar] [CrossRef]
- Rose, E.C.; Jonas, I.E.; Kappert, H.F. In vitro investigation into the biological assessment of orthodontic wires. J. Orofac. Orthop. Fortschr. Kieferorthopädie 1998, 59, 253–264. [Google Scholar] [CrossRef]
- Ryhänen, J.; Niemi, E.; Serlo, W.; Niemelä, E.; Sandvik, P.; Pernu, H.; Salo, T. Biocompatibility of nickel-titanium shape memory metal and its corrosion behavior in human cell cultures. J. Biomed. Mater. Res. Part A 1997, 35, 451–457. [Google Scholar] [CrossRef]
- Celebi, F.; Altun, A.; Bicakci, A.A. Cytotoxicity of bracket identification dyes. Angle Orthod. 2019, 89, 426–431. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cr | Mn | Fe | Ni | Cu | Mo | Pb | Zn | Pd | Ag | Co | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | 0.6 (11%) | 1.0 (11%) | 85.7 (4%) | 0.8 (30%) | 3.7 (54%) | 0.2 (13%) | 0.2 (23%) | 1.6 (2%) | 0.4 (17%) | 0.1 (8%) | 0.0 (16%) |
| P1 | 16.6 (5%) | 10.2 (5%) | 68.5 (11%) | 65.6 (2%) | 19,500.1 (1%) | 1.6 (17%) | 0.1 (41%) | 1.7 (6%) | 16.2 (4%) | 3.7 (16%) | 0.0 (9%) |
| P2 | 9.3 (2%) | 3.9 (7%) | 75.9 (9%) | 10.6 (8%) | 101.1 (5%) | 2.8 (5%) | 0.1 (26%) | 1.1 (5%) | 0.5 (6%) | 0.0 (17%) | 0.0 (9%) |
| G1 | 5.5 (3%) | 7.3 (9%) | 77.7 (4%) | 16.0 (2%) | 337.0 (2%) | 5.8 (6%) | 0.1 (22%) | 3.6 (7%) | 0.3 (4%) | 0.0 (80%) | 0.9 (3%) |
| G2 | 2.5 (4%) | 2.2 (8%) | 60.3 (14%) | 7.4 (9%) | 432.6 (1%) | 0.7 (4%) | 0.1 (36%) | 1.8 (16%) | 0.3 (14%) | 0.0 (45%) | 0.0 (19%) |
| G3 | 3.5 (3%) | 1.7 (15%) | 52.3 (6%) | 8.7 (4%) | 316.1 (2%) | 3.9 (10%) | 0.1 (89%) | 1.2 (21%) | 0.2 (2%) | 0.0 (142%) | 0.0 (6%) |
| G4 | 5.4 (5%) | 1.7 (7%) | 78.5 (9%) | 1923.7 (4%) | 1147.3 (2%) | 22.8 (6%) | 0.1 (63%) | 0.8 (17%) | 0.4 (10%) | 0.0 (27%) | 0.1 (15%) |
| G5 | 3.4 (5%) | 3.9 (5%) | 40.8 (1%) | 6.3 (26%) | 10.7 (9%) | 0.2 (10%) | 0.6 (4%) | 1.8 (7%) | 0.2 (16%) | 0.0 (22%) | 0.0 (10%) |
| Treatment | Total Cells Tracked | Cell Divisions | Lost Cells | Apoptosis |
|---|---|---|---|---|
| Control | 94 | 34 | 7 | 1 |
| P1 | 59 | 16 | 0 | 1 |
| P2 | 55 | 13 | 1 | 1 |
| G1 | 39 | 6 | 0 | 0 |
| G2 | 71 | 23 | 0 | 0 |
| G3 | 63 | 15 | 0 | 0 |
| G4 | 58 | 17 | 0 | 0 |
| G5 | 52 | 12 | 3 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wishney, M.; Mahadevan, S.; Cornwell, J.A.; Savage, T.; Proschogo, N.; Darendeliler, M.A.; Zoellner, H. Toxicity of Orthodontic Brackets Examined by Single Cell Tracking. Toxics 2022, 10, 460. https://doi.org/10.3390/toxics10080460
Wishney M, Mahadevan S, Cornwell JA, Savage T, Proschogo N, Darendeliler MA, Zoellner H. Toxicity of Orthodontic Brackets Examined by Single Cell Tracking. Toxics. 2022; 10(8):460. https://doi.org/10.3390/toxics10080460
Chicago/Turabian StyleWishney, Morgan, Swarna Mahadevan, James Anthony Cornwell, Tom Savage, Nick Proschogo, M. Ali Darendeliler, and Hans Zoellner. 2022. "Toxicity of Orthodontic Brackets Examined by Single Cell Tracking" Toxics 10, no. 8: 460. https://doi.org/10.3390/toxics10080460
APA StyleWishney, M., Mahadevan, S., Cornwell, J. A., Savage, T., Proschogo, N., Darendeliler, M. A., & Zoellner, H. (2022). Toxicity of Orthodontic Brackets Examined by Single Cell Tracking. Toxics, 10(8), 460. https://doi.org/10.3390/toxics10080460