
Sulfonate-Modified Polystyrene Nanoparticle at Precited Environmental Concentrations Induces Transgenerational Toxicity Associated with Increase in Germline Notch Signal of Caenorhabditis elegans



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
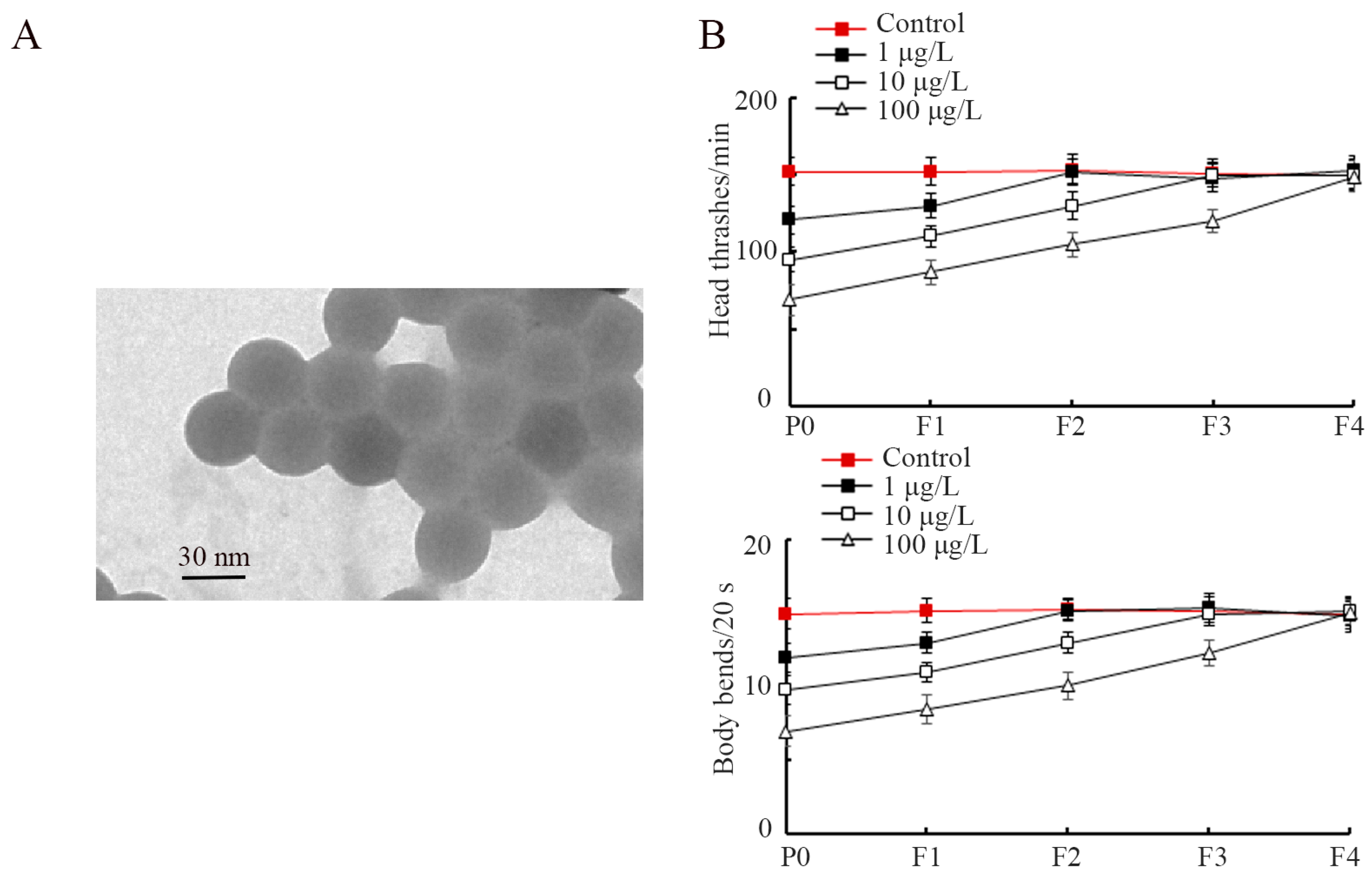
2.1. PS-S NP Characterization
2.2. C. elegans Maintenance
2.3. Exposure
2.4. Locomotion Behaviors
2.5. Reproductive Capacity
2.6. Transcriptional Expression Analysis
2.7. RNA Interference (RNAi)
2.8. Data Analysis
3. Results
3.1. Exposure to PS-S NP Induced Transgenerational Locomotion Inhibition
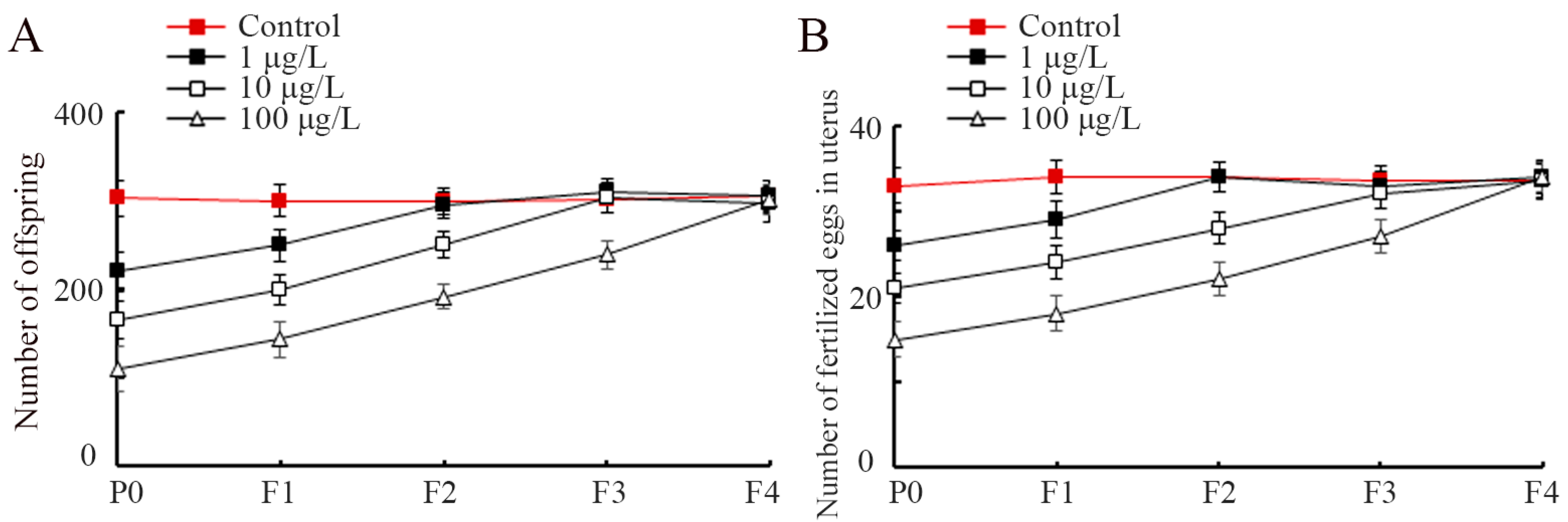
3.2. Exposure to PS-S NP-Induced Transgenerational Inhibition in Reproductive Capacity
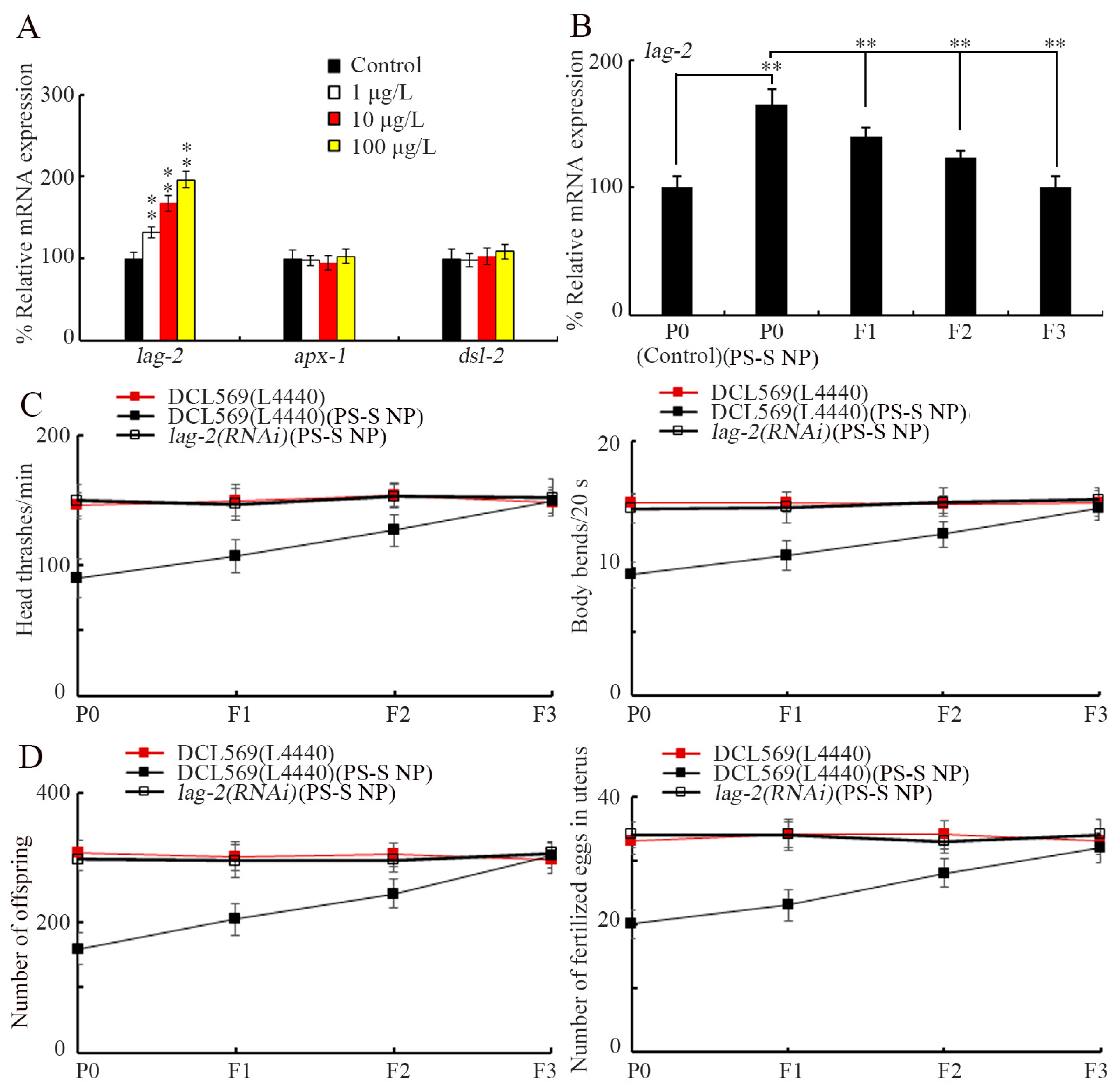
3.3. Exposure to PS-S NP Induced Transgenerational Increase in Expression of Germline lag-2
3.4. Germline Notch Ligand LAG-2 Was Required for Induction of Transgenerational PS-S NP Toxicity
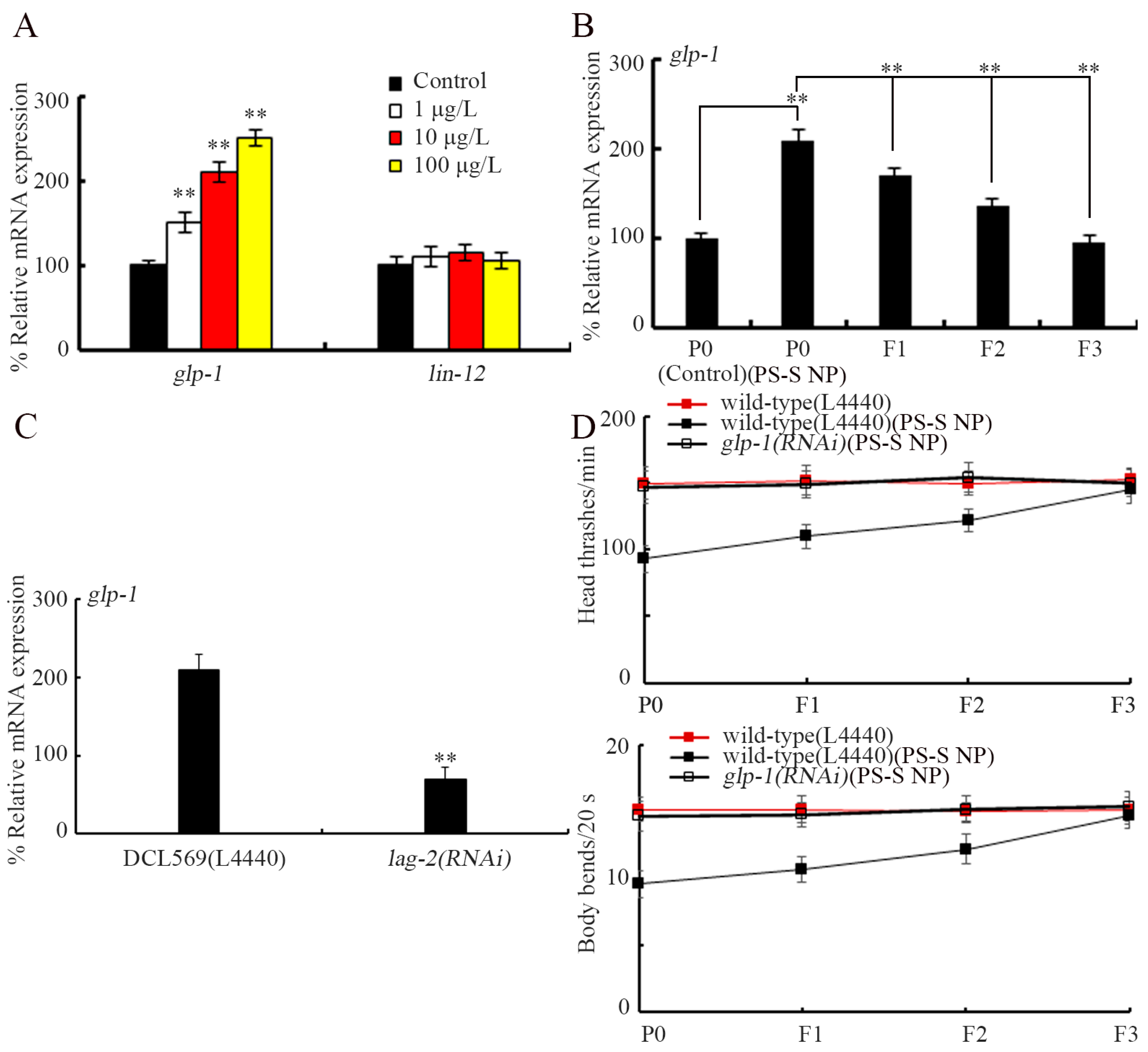
3.5. Exposure to PS-S NP Caused Transgenerational Increase in Expression of glp-1
3.6. Notch Receptor GLP-1 Was Involved in Induction of Transgenerational PS-S NP Toxicity
3.7. Tissue-Specific Activities of GLP-1 in Controlling Transgenerational PS-S NP Toxicity
3.8. Identification of Potential Downstream Targets of Germline GLP-1 in Controlling Transgenerational PS-S NP Toxicity
3.9. Identification of Potential Downstream Targets of Neuronal GLP-1 in Controlling Transgenerational PS-S NP Toxicity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Geyer, R.; Jambeck, J.R.; Law, K.L. Production, use, and fate of all plastics ever made. Sci. Adv. 2017, 3, e1700782. [Google Scholar] [CrossRef] [Green Version]
- Gigault, J.; Halle, A.T.; Baudrimont, M.; Pascal, P.Y.; Gauffre, F.; Phi, T.L.; El Hadri, H.; Grassl, B.; Reynaud, S. Current opinion: What is a nanoplastic? Environ. Pollut. 2018, 235, 1030–1034. [Google Scholar] [CrossRef]
- Tang, M.; Ding, G.; Lu, X.; Huang, Q.; Du, H.; Xiao, G.; Wang, D. Exposure to nanoplastic particles enhances Acinetobacter survival, biofilm formation, and serum resistance. Nanomaterials 2022, 12, 4222. [Google Scholar] [CrossRef]
- Lebreton, L.; Andrady, A. Future scenarios of global plastic waste generation and disposal. Palgrave Commun. 2019, 5, 6. [Google Scholar] [CrossRef] [Green Version]
- Marcelino, R.C.; Cardoso, R.M.; Domingues, E.L.B.C.; Gonçalves, R.V.; Lima, G.D.A.; Novaes, R.D. The emerging risk of microplastics and nanoplastics on the microstructure and function of reproductive organs in mammals: A systematic review of preclinical evidence. Life Sci. 2022, 295, 120404. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Nag, R.; Cummins, E. Human health concerns regarding microplastics in the aquatic environment—From marine to food systems. Sci. Total Environ. 2022, 823, 153730. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Fu, D.; Qi, H.; Lan, C.Q.; Yu, H.; Ge, C. Micro- and nano-plastics in marine environment: Source, distribution and threats—A review. Sci. Total Environ. 2020, 698, 134254. [Google Scholar] [CrossRef]
- Wang, L.; Wu, W.M.; Bolan, N.S.; Tsang, D.C.W.; Li, Y.; Qin, M.; Hou, D. Environmental fate, toxicity and risk management strategies of nanoplastics in the environment: Current status and future perspectives. J. Hazard. Mater. 2021, 401, 123415. [Google Scholar] [CrossRef]
- Prata, J.C. Airborne microplastics: Consequences to human health? Environ. Pollut. 2018, 234, 115–126. [Google Scholar] [CrossRef]
- Waring, R.H.; Harris, R.M.; Mitchell, S.C. Plastic contamination of the food chain: A threat to human health? Maturitas 2018, 115, 64–68. [Google Scholar] [CrossRef]
- Lett, Z.; Hall, A.; Skidmore, S.; Alves, N.J. Environmental microplastic and nanoplastic: Exposure routes and effects on coagulation and the cardiovascular system. Environ. Pollut. 2021, 291, 118190. [Google Scholar] [CrossRef] [PubMed]
- Sangkham, S.; Faikhaw, O.; Munkong, N.; Sakunkoo, P.; Arunlertaree, C.; Chavali, M.; Mousazadeh, M.; Tiwari, A. A review on microplastics and nanoplastics in the environment: Their occurrence, exposure routes, toxic studies, and potential effects on human health. Mar. Pollut. Bull. 2022, 181, 113832. [Google Scholar] [CrossRef] [PubMed]
- Walczak, A.P.; Hendriksen, P.J.; Woutersen, R.A.; van der Zande, M.; Undas, A.K.; Helsdingen, R.; van den Berg, H.H.; Rietjens, I.M.; Bouwmeester, H. Bioavailability and biodistribution of differently charged polystyrene nanoparticles upon oral exposure in rats. J. Nanopart. Res. 2015, 17, 231. [Google Scholar] [CrossRef] [Green Version]
- Hua, X.; Wang, D.-Y. Cellular uptake, transport, and organelle response after exposure to microplastics and nanoplastics: Current knowledge and perspectives for environmental and health risks. Rev. Environ. Contam. Toxicol. 2022, 260, 12. [Google Scholar] [CrossRef]
- Smith, M.; Love, D.C.; Rochman, C.M.; Neff, R.A. Microplastics in seafood and the implications for human health. Curr. Environ. Health Rep. 2018, 5, 375–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samadi, A.; Kim, Y.; Lee, S.A.; Kim, Y.J.; Esterhuizen, M. Review on the ecotoxicological impacts of plastic pollution on the freshwater invertebrate Daphnia. Environ. Toxicol. 2022, 37, 2615–2638. [Google Scholar] [CrossRef]
- Chang, X.; Xue, Y.; Li, J.; Zou, L.; Tang, M. Potential health impact of environmental micro- and nanoplastics pollution. J. Appl. Toxicol. 2020, 40, 4–15. [Google Scholar] [CrossRef]
- Bhagat, J.; Zang, L.; Nishimura, N.; Shimada, Y. Zebrafish: An emerging model to study microplastic and nanoplastic toxicity. Sci. Total Environ. 2020, 728, 138707. [Google Scholar] [CrossRef]
- Banerjee, A.; Shelver, W.L. Micro- and nanoplastic induced cellular toxicity in mammals: A review. Sci. Total Environ. 2021, 755, 142518. [Google Scholar] [CrossRef]
- López, A.D.F.; Fabiani, M.; Lassalle, V.L.; Spetter, C.V.; Severini, M.D.F. Critical review of the characteristics, interactions, and toxicity of micro/nanomaterials pollutants in aquatic environments. Mar. Pollut. Bull. 2022, 174, 113276. [Google Scholar] [CrossRef]
- Liu, J.; Ma, Y.; Zhu, D.; Xia, T.; Qi, Y.; Yao, Y.; Guo, X.; Ji, R.; Chen, W. Polystyrene nanoplastics-enhanced contaminant transport: Role of irreversible adsorption in glassy polymeric domain. Environ. Sci. Technol. 2018, 52, 2677–2685. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Zhang, W.; Lin, T.; Liu, S.; Sun, Z.; Liu, F.; Yuan, Y.; Xiang, X.; Kuang, H.; Yang, B.; et al. Maternal exposure to polystyrene nanoplastics during gestation and lactation induces hepatic and testicular toxicity in male mouse offspring. Food Chem. Toxicol. 2022, 160, 112803. [Google Scholar] [CrossRef] [PubMed]
- Junaid, M.; Liu, S.; Chen, G.; Liao, H.; Wang, J. Transgenerational impacts of micro(nano)plastics in the aquatic and terrestrial environment. J. Hazard. Mater. 2023, 443, 130274. [Google Scholar] [CrossRef]
- Nogueira, D.J.; Silva, A.C.O.D.; da Silva, M.L.N.; Vicentini, D.S.; Matias, W.G. Individual and combined multigenerational effects induced by polystyrene nanoplastic and glyphosate in Daphnia magna (Strauss, 1820). Sci. Total Environ. 2022, 811, 151360. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Wang, C.; Li, H.; Ma, R.; Yu, Z.; Li, L.; Xiang, M.; Chen, X.; Hua, X.; Yu, Y. A review of toxicity induced by persistent organic pollutants (POPs) and endocrine-disrupting chemicals (EDCs) in the nematode Caenorhabditis elegans. J. Environ. Manag. 2019, 237, 519–525. [Google Scholar] [CrossRef]
- Hartman, J.H.; Widmayer, S.J.; Bergemann, C.M.; King, D.E.; Morton, K.S.; Romersi, R.F.; Jameson, L.E.; Leung, M.C.K.; Andersen, E.C.; Taubert, S.; et al. Xenobiotic metabolism and transport in Caenorhabditis elegans. J. Toxicol. Environ. Health B Crit. Rev. 2021, 24, 51–94. [Google Scholar] [CrossRef] [PubMed]
- Hua, X.; Feng, X.; Liang, G.-Y.; Chao, J.; Wang, D.-Y. Exposure to 6-PPD quinone at environmentally relevant concentrations causes abnormal locomotion behaviors and neurodegeneration in Caenorhabditis elegans. Environ. Sci. Technol. 2023, 57, 4940–4950. [Google Scholar] [CrossRef]
- Hua, X.; Feng, X.; Liang, G.-Y.; Chao, J.; Wang, D.-Y. Long-term exposure to 6-PPD quinone reduces reproductive capacity by enhancing germline apoptosis associated with activation of both DNA damage and cell corpse engulfment in Caenorhabditis elegans. J. Hazard. Mater. 2023, 454, 131495. [Google Scholar] [CrossRef]
- Wang, Y.-X.; Yuan, X.-A.; Zhou, R.; Bu, Y.-Q.; Wang, D.-Y. Combinational exposure to hydroxyatrazine increases neurotoxicity of polystyrene nanoparticles on Caenorhabditis elegans. Sci. Total Environ. 2023, 880, 163283. [Google Scholar] [CrossRef]
- Qu, M.; Kong, Y.; Yuan, Y.-J.; Wang, D.-Y. Neuronal damage induced by nanopolystyrene particles in nematode Caenorhabditis elegans. Environ. Sci. Nano 2019, 6, 2591–2601. [Google Scholar] [CrossRef]
- Hua, X.; Cao, C.; Zhang, L.; Wang, D.-Y. Activation of FGF signal in germline mediates transgenerational toxicity of polystyrene nanoparticles at predicted environmental concentrations in Caenorhabditis elegans. J. Hazard. Mater. 2023, 451, 131174. [Google Scholar] [CrossRef]
- Schultz, C.L.; Bart, S.; Lahive, E.; Spurgeon, D.J. What is on the outside matters-surface charge and dissolve organic matter association affect the toxicity and physiological mode of action of polystyrene nanoplastics to C. elegans. Environ. Sci. Technol. 2021, 55, 6065–6075. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.M.; Lee, D.K.; Long, N.P.; Kwon, S.W.; Park, J.H. Uptake of nanopolystyrene particles induces distinct metabolic profiles and toxic effects in Caenorhabditis elegans. Environ. Pollut. 2019, 246, 578–586. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.-Y. Exposure Toxicology in Caenorhabditis elegans; Springer Nature Singapore Pte Ltd.: Singapore, 2020. [Google Scholar]
- Chen, H.; Hua, X.; Li, H.; Wang, C.; Dang, Y.; Ding, P.; Yu, Y. Transgenerational neurotoxicity of polystyrene microplastics induced by oxidative stress in Caenorhabditis elegans. Chemosphere 2021, 272, 129642. [Google Scholar] [CrossRef]
- Ni, S.; Zhang, H.; Sun, L.; Zhao, Y.; Pei, C.; Nie, Y.; Liu, X.; Wu, L.; Xu, A. Transgenerational reproductive toxicity of 2,4,6-trinitrotoluene (TNT) and its metabolite 4-ADNT in Caenorhabditis elegans. Environ. Toxicol. Pharmacol. 2022, 92, 103865. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, S.-T.; Zhao, Y.-L.; Nurdebek, B.; Bu, Y.-Q.; Wang, D.-Y. Long-term exposure to polystyrene nanoparticles causes transgenerational toxicity by affecting the function and expression of MEV-1 and DAF-2 signals in Caenorhabditis elegans. NanoImpact 2022, 26, 100403. [Google Scholar] [CrossRef]
- Zhao, Y.; Hua, X.; Bian, Q.; Wang, D.-Y. Nanoplastic exposure at predicted environmental concentrations induces activation of germline Ephrin signal associated with toxicity formation in the Caenorhabditis elegans offspring. Toxics 2022, 10, 699. [Google Scholar] [CrossRef]
- Hua, X.; Zhao, Y.; Yuan, Y.-J.; Zhang, L.; Bian, Q.; Wang, D.-Y. Nanoplastics cause transgenerational toxicity through inhibiting germline microRNA mir-38 in C. elegans. J. Hazard. Mater. 2022, 437, 129302. [Google Scholar] [CrossRef]
- Yu, C.W.; Luk, T.C.; Liao, V.H. Long-term nanoplastics exposure results in multi and trans-generational reproduction decline associated with germline toxicity and epigenetic regulation in Caenorhabditis elegans. J. Hazard. Mater. 2021, 412, 125173. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, S.-T.; Zhao, Y.; Bi, K.; Wang, D.-Y. Increase in germline methyltransferases governing methylation of histone H3K9 is associated with transgenerational nanoplastic toxicity in Caenorhabditis elegans. Environ. Sci. Nano 2022, 9, 265–274. [Google Scholar] [CrossRef]
- Wang, X.; Chen, X.; Peng, Y.; Pan, J. Silver-modified porous polystyrene sulfonate derived from Pickering high internal phase emulsions for capturing lithium-ion. RSC Adv. 2019, 9, 7228–7237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, J.P.; Suhali-Amacher, N.; Ray, S.D. Metals and metal antagonists. Side Eff. Drugs Annu. 2017, 39, 197–208. [Google Scholar]
- Nashruddin, S.N.A.; Abdullah, J.; Mohammad Haniff, M.A.S.; Mat Zaid, M.H.; Choon, O.P.; Mohd Razip Wee, M.F. Label free glucose electrochemical biosensor based on poly(3,4-ethylenedioxy thiophene):Polystyrene sulfonate/titanium carbide/graphene quantum dots. Biosensors 2021, 11, 267. [Google Scholar] [CrossRef]
- Qu, M.; Wang, D.-Y. Toxicity comparison between pristine and sulfonate modified nanopolystyrene particles in affecting locomotion behavior, sensory perception, and neuronal development in Caenorhabditis elegans. Sci. Total Environ. 2020, 703, 134817. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.-L.; Chen, J.-Y.; Wang, R.; Pu, X.-X.; Wang, D.-Y. A review of transgenerational and multigenerational toxicology in the in vivo model animal Caenorhabditis elegans. J. Appl. Toxicol. 2023, 43, 122–145. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.-R.; Hua, X.; Rui, Q.; Wang, D.-Y. Polystyrene nanoparticles caused dynamic alteration in mitochondrial unfolded protein response from parents to the offspring in C. elegans. Chemosphere 2022, 308, 136154. [Google Scholar] [CrossRef]
- Kopan, R.; Ilagan, M.X. The canonical Notch signaling pathway: Unfolding the activation mechanism. Cell 2009, 137, 216–233. [Google Scholar] [CrossRef] [Green Version]
- Artavanis-Tsakonas, S.; Rand, M.D.; Lake, R.J. Notch signaling: Cell fate control and signal integration in development. Science 1999, 284, 770–776. [Google Scholar] [CrossRef] [Green Version]
- Chen, N.; Greenwald, I. The lateral signal for LIN-12/Notch in C. elegans vulval development comprises redundant secreted and transmembrane DSL proteins. Dev. Cell 2004, 6, 183–192. [Google Scholar] [CrossRef] [Green Version]
- Priess, J. Notch Signaling in the C. elegans Embryo. WormBook. 2005. Available online: http://www.wormbook.org/chapters/www_notchembryo/notchembryo.html (accessed on 16 April 2023). [CrossRef] [Green Version]
- Liu, H.-L.; Tian, L.-J.; Wang, D.-Y. Notch receptor GLP-1 regulates toxicity of simulated microgravity stress by activating germline-intestine communication of insulin signaling in C. elegans. Biochem. Biophys. Res. Commun. 2021, 534, 248–253. [Google Scholar] [CrossRef]
- Qu, M.; Qiu, Y.; Lv, R.; Yue, Y.; Liu, R.; Yang, F.; Wang, D.; Li, Y. Exposure to MPA-capped CdTe quantum dots causes reproductive toxicity effects by affecting oogenesis in nematode Caenorhabditis elegans. Ecotoxicol. Environ. Saf. 2019, 173, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, Y.; Cao, C.; Zhu, Y.; Huang, W.; Yang, Y.; Qiu, H.; Liu, S.; Wang, D. Beneficial effect of Xuebijing against Pseudomonas aeruginosa infection in Caenorhabditis elegans. Front. Pharmacol. 2022, 13, 949608. [Google Scholar] [CrossRef]
- Wang, Y.-X.; Zhang, L.; Yuan, X.-A.; Wang, D.-Y. Treatment with paeoniflorin increases lifespan of Pseudomonas aeruginosa infected Caenorhabditis elegans by inhibiting bacterial accumulation in intestinal lumen and biofilm formation. Front. Pharmacol. 2023, 14, 1114219. [Google Scholar] [CrossRef]
- Tsalik, E.L.; Hobert, O. Functional mapping of neurons that control locomotory behavior in Caenorhabditis elegans. J. Neurobiol. 2003, 56, 178–197. [Google Scholar] [CrossRef] [PubMed]
- Hua, X.; Feng, X.; Hua, Y.-S.; Wang, D.-Y. Paeoniflorin attenuates polystyrene nanoparticle-induced reduction in reproductive capacity and increase in germline apoptosis through suppressing DNA damage checkpoints in Caenorhabditis elegans. Sci. Total Environ. 2023, 871, 162189. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.-Y.; Xu, R.-R.; Hua, X.; Rui, Q.; Wang, D.-Y. Multi-walled carbon nanotubes induce transgenerational toxicity associated with activation of germline long non-coding RNA linc-7 in C. elegans. Chemosphere 2022, 301, 134687. [Google Scholar] [CrossRef]
- Zhao, Y.-Y.; Hua, X.; Rui, Q.; Wang, D.-Y. Exposure to multi-walled carbon nanotubes causes suppression in octopamine signal associated with transgenerational toxicity induction in C. elegans. Chemosphere 2023, 318, 137986. [Google Scholar] [CrossRef]
- Yang, Y.-H.; Wu, Q.-L.; Wang, D.-Y. Neuronal Gα subunits required for the control of response to polystyrene nanoparticles in the range of μg/L in C. elegans. Ecotoxicol. Environ. Saf. 2021, 225, 112732. [Google Scholar] [CrossRef]
- Hua, X.; Feng, X.; Liang, G.-Y.; Chao, J.; Wang, D.-Y. Long-term exposure to tire-derived 6-PPD quinone causes intestinal toxicity by affecting functional state of intestinal barrier in Caenorhabditis elegans. Sci. Total Environ. 2023, 861, 160591. [Google Scholar] [CrossRef]
- Yang, Y.-H.; Wu, Q.-L.; Wang, D.-Y. Epigenetic response to nanopolystyrene in germline of nematode Caenorhabditis elegans. Ecotoxicol. Environ. Saf. 2020, 206, 111404. [Google Scholar] [CrossRef]
- Austin, J.; Kimble, J. glp-1 is required in the germ line for regulation of the decision between mitosis and meiosis in C. elegans. Cell 1987, 51, 589–599. [Google Scholar] [CrossRef]
- Pepper, A.S.-R.; Killian, D.J.; Hubbard, E.J.A. Genetic analysis of Caenorhabditis elegans glp-1 mutants suggests receptor interaction or competition. Genetics 2003, 163, 115–132. [Google Scholar] [CrossRef]
- Ouellet, J.; Li, S.; Roy, R. Notch signalling is required for both dauer maintenance and recovery in C. elegans. Development 2008, 135, 2583–2592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crittenden, S.L.; Troemel, E.R.; Evans, T.C.; Kimble, J. GLP-1 is localized to the mitotic region of the C. elegans germ line. Development 1994, 120, 2901–2911. [Google Scholar] [CrossRef] [PubMed]
- Jantschplunger, V.; Fire, A. Combinatorial structure of a body muscle-specific transcriptional enhancer in Caenorhabditis elegans. J. Biol. Chem. 1994, 269, 27021–27028. [Google Scholar] [CrossRef]
- Liu, H.-L.; Zhao, Y.-L.; Hua, X.; Wang, D.-Y. Induction of transgenerational toxicity is associated with the activated germline insulin signals in nematodes exposed to nanoplastic at predicted environmental concentrations. Ecotoxicol. Environ. Saf. 2022, 243, 114022. [Google Scholar] [CrossRef]
- Xu, R.-R.; Hua, X.; Rui, Q.; Wang, D.-Y. Alteration in Wnt signaling mediates induction of transgenerational toxicity of polystyrene nanoplastics in C. elegans. NanoImpact 2022, 28, 100425. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.-L.; Zhang, R.-J.; Wang, D.-Y. Response of DBL-1/TGF-β signaling-mediated neuron-intestine communication to nanopolystyrene in nematode Caenorhabditis elegans. Sci. Total Environ. 2020, 745, 1141047. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.-L.; Zhao, Y.-Y.; Bi, Q.; Rui, Q.; Wang, D.-Y. Dysregulated mir-76 mediated a protective response to nanopolystyrene by modulating heme homeostasis related molecular signaling in nematode Caenorhabditis elegans. Ecotoxicol. Environ. Saf. 2021, 212, 112018. [Google Scholar] [CrossRef]
- Wang, S.-T.; Liu, H.-L.; Zhao, Y.-Y.; Rui, Q.; Wang, D.-Y. Dysregulated mir-354 enhanced the protective response to nanopolystyrene by affecting the activity of TGF-β signaling pathway in nematode Caenorhabditis elegans. NanoImpact 2020, 20, 100256. [Google Scholar] [CrossRef]
- Qu, M.; Li, D.; Zhao, Y.-L.; Yuan, Y.-J.; Wang, D.-Y. Exposure to low-dose nanopolystyrene induces the response of neuronal JNK MAPK signaling pathway in nematode Caenorhabditis elegans. Environ. Sci. Eur. 2020, 32, 58. [Google Scholar] [CrossRef] [Green Version]
- Qu, M.; Li, D.; Qiu, Y.-X.; Wang, D.-Y. Neuronal ERK MAPK signaling in response to low-dose nanopolystyrene exposure by suppressing insulin peptide expression in Caenorhabditis elegans. Sci. Total Environ. 2020, 724, 138378. [Google Scholar] [CrossRef] [PubMed]
- Kokilathasan, N.; Dittrich, M. Nanoplastics: Detection and impacts in aquatic environments—A review. Sci. Total Environ. 2022, 849, 157852. [Google Scholar] [CrossRef]
- Pashaei, R.; Dzingelevičienė, R.; Abbasi, S.; Szultka-Młyńska, M.; Buszewski, B. Determination of the pharmaceuticals-nano/microplastics in aquatic systems by analytical and instrumental methods. Environ. Monit. Assess. 2022, 194, 93. [Google Scholar] [CrossRef] [PubMed]
- Kögel, T.; Bjorøy, Ø.; Toto, B.; Bienfait, A.M.; Sanden, M. Micro- and nanoplastic toxicity on aquatic life: Determining factors. Sci. Total Environ. 2020, 709, 136050. [Google Scholar] [CrossRef] [PubMed]
- Trevisan, R.; Ranasinghe, P.; Jayasundara, N.; Di Giulio, R.T. Nanoplastics in aquatic environments: Impacts on aquatic species and interactions with environmental factors and pollutants. Toxics 2022, 10, 326. [Google Scholar] [CrossRef]
- Liu, H.-L.; Tian, L.-J.; Wang, S.-T.; Wang, D.-Y. Size-dependent transgenerational toxicity induced by nanoplastics in nematode Caenorhabditis elegans. Sci. Total Environ. 2021, 790, 148217. [Google Scholar] [CrossRef]
- Sun, L.-M.; Liao, K.; Wang, D.-Y. Comparison of transgenerational reproductive toxicity induced by pristine and amino modified nanoplastics in Caenorhabditis elegans. Sci. Total Environ. 2021, 768, 144362. [Google Scholar] [CrossRef]
- Lenz, R.; Enders, K.; Nielsen, T.G. Microplastic exposure studies should be environmentally realistic. Proc. Natl. Acad. Sci. USA 2016, 113, E4121–E4122. [Google Scholar] [CrossRef] [Green Version]
- Guimarães, A.T.B.; Estrela, F.N.; Rodrigues, A.S.L.; Chagas, T.Q.; Pereira, P.S.; Silva, F.G.; Malafaia, G. Nanopolystyrene particles at environmentally relevant concentrations causes behavioral and biochemical changes in juvenile grass carp (Ctenopharyngodon idella). J. Hazard. Mater. 2021, 403, 123864. [Google Scholar] [CrossRef] [PubMed]
- Sprinzak, D.; Blacklow, S.C. Biophysics of Notch signaling. Annu. Rev. Biophys. 2021, 50, 157–189. [Google Scholar] [CrossRef] [PubMed]
- Tax, F.E.; Yeargers, J.J.; Thomas, J.H. Sequence of C. elegans lag-2 reveals a cell-signalling domain shared with Delta and Serrate of Drosophila. Nature 1994, 368, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Greenwald, I. Spatial regulation of lag-2 transcription during vulval precursor cell fate patterning in Caenorhabditis elegans. Genetics 2011, 188, 847–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertrand, V.; Bisso, P.; Poole, R.J.; Hobert, O. Notch-dependent induction of left/right asymmetry in C. elegans interneurons and motoneurons. Curr. Biol. 2011, 21, 1225–1231. [Google Scholar] [CrossRef] [Green Version]
- Henderson, S.T.; Gao, D.; Lambie, E.J.; Kimble, J. lag-2 may encode a signaling ligand for the GLP-1 and LIN-12 receptors of C. elegans. Development 1994, 120, 2913–2924. [Google Scholar] [CrossRef]
- Henderson, S.T.; Gao, D.; Christensen, S.; Kimble, J. Functional domains of LAG-2, a putative signaling ligand for LIN-12 and GLP-1 receptors in Caenorhabditis elegans. Mol. Biol. Cell 1997, 8, 1751–1762. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, H.A.; Fitzgerald, K.; Greenwald, I. Reciprocal changes in expression of the receptor lin-12 and its ligand lag-2 prior to commitment in a C. elegans cell fate decision. Cell 1994, 79, 1187–1198. [Google Scholar] [CrossRef]
- Mango, S.E.; Maine, E.M.; Kimble, J. Carboxy-terminal truncation activates glp-1 protein to specify vulval fates in Caenorhabditis elegans. Nature 1991, 352, 811–815. [Google Scholar] [CrossRef] [Green Version]
- Fitzgerald, K.; Wilkinson, H.A.; Greenwald, I. glp-1 can substitute for lin-12 in specifying cell fate decisions in Caenorhabditis elegans. Development 1993, 119, 1019–1027. [Google Scholar] [CrossRef]
- Moskowitz, I.P.; Rothman, J.H. lin-12 and glp-1 are required zygotically for early embryonic cellular interactions and are regulated by maternal GLP-1 signaling in Caenorhabditis elegans. Development 1996, 122, 4105–4117. [Google Scholar] [CrossRef]
- Berry, L.W.; Westlund, B.; Schedl, T. Germ-line tumor formation caused by activation of glp-1, a Caenorhabditis elegans member of the Notch family of receptors. Development 1997, 124, 925–936. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, S.; Martin, D.; Meléndez, A.; Bargonetti, J. C. elegans CEP-1/p53 and BEC-1 are involved in DNA repair. PLoS ONE 2014, 9, e88828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, D.; Kahler, D.J.; Yun, C.; Hubbard, E.J.A. Functional interactions between rsks-1/S6K, glp-1/Notch, and regulators of Caenorhabditis elegans fertility and germline stem cell maintenance. G3 2018, 8, 3293–3309. [Google Scholar] [CrossRef]
- Singh, K.; Chao, M.Y.; Somers, G.A.; Komatsu, H.; Corkins, M.E.; Larkins-Ford, J.; Tucey, T.; Dionne, H.M.; Walsh, M.B.; Beaumont, E.K.; et al. C. elegans Notch signaling regulates adult chemosensory response and larval molting quiescence. Curr. Biol. 2011, 21, 825–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, W.; Gu, A.; Wang, D. Sulfonate-Modified Polystyrene Nanoparticle at Precited Environmental Concentrations Induces Transgenerational Toxicity Associated with Increase in Germline Notch Signal of Caenorhabditis elegans. Toxics 2023, 11, 511. https://doi.org/10.3390/toxics11060511
He W, Gu A, Wang D. Sulfonate-Modified Polystyrene Nanoparticle at Precited Environmental Concentrations Induces Transgenerational Toxicity Associated with Increase in Germline Notch Signal of Caenorhabditis elegans. Toxics. 2023; 11(6):511. https://doi.org/10.3390/toxics11060511
Chicago/Turabian StyleHe, Wenmiao, Aihua Gu, and Dayong Wang. 2023. "Sulfonate-Modified Polystyrene Nanoparticle at Precited Environmental Concentrations Induces Transgenerational Toxicity Associated with Increase in Germline Notch Signal of Caenorhabditis elegans" Toxics 11, no. 6: 511. https://doi.org/10.3390/toxics11060511
APA StyleHe, W., Gu, A., & Wang, D. (2023). Sulfonate-Modified Polystyrene Nanoparticle at Precited Environmental Concentrations Induces Transgenerational Toxicity Associated with Increase in Germline Notch Signal of Caenorhabditis elegans. Toxics, 11(6), 511. https://doi.org/10.3390/toxics11060511