

Exploration of the Effects of Cadmium Stress on Photosynthesis in Oenanthe javanica (Blume) DC.

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Cd Treatment
2.3. Determination of PF, MR, and DF
2.4. Quantification of Pigment, H2O2, MDA and Cd Contents
2.5. Data Processing
3. Results
3.1. Growth, Pigment Content, H2O2 Content, MDA Content and Cd Concentration
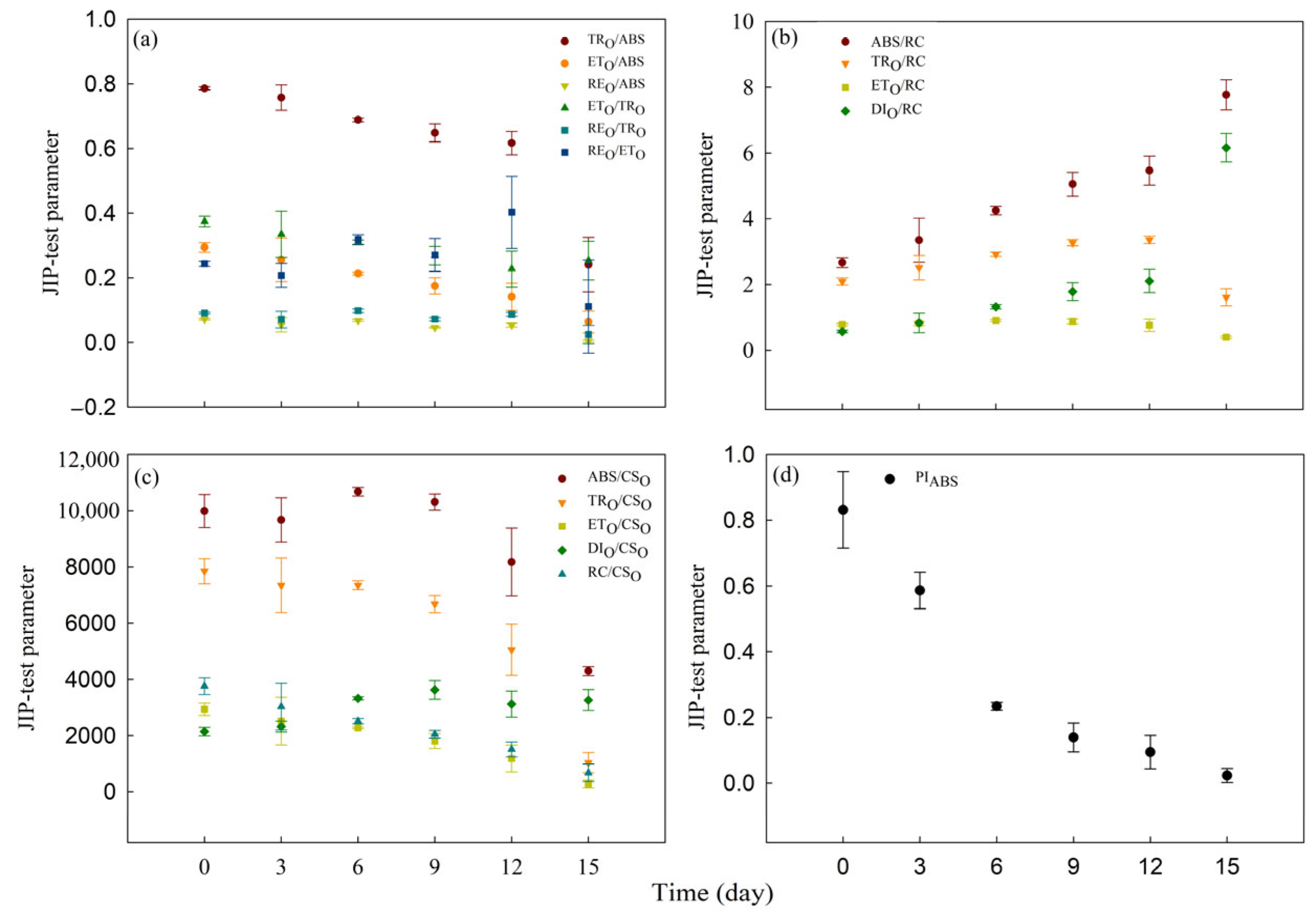
3.2. Effect of Cd Stress on Rapid Chlorophyll Fluorescence Kinetic Curves and JIP-Test Parameters
3.3. Effect of Cd Stress on MR
3.4. Effect of Cd Stress on DF
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tötsch, W. Cadmium-towards a rational use of a toxic element. Environ. Manag. 1990, 14, 333–338. [Google Scholar] [CrossRef]
- Grant, C.A.; Clarke, J.M.; Duguid, S.; Chaney, R.L. Selection and breeding of plant cultivars to minimize cadmium accumulation. Sci. Total Environ. 2008, 390, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Rizwan, M.; Ali, S.; Adrees, M.; Rizvi, H.; Ziaur-Rehman, M.; Hannan, F.; Qayyum, M.F.; Hafeez, F.; Ok, Y.S. Cadmium stress in rice: Toxic effects, tolerance mechanisms, and management: A critical review. Environ. Sci. Pollut. R 2016, 23, 17859–17879. [Google Scholar] [CrossRef] [PubMed]
- Nagajyoti, P.C.; Lee, K.D.; Sreekanth, T.V.M. Heavy metals, occurrence and toxicity for plants: A review. Environ. Chem. Lett. 2010, 8, 199–216. [Google Scholar] [CrossRef]
- Lysenko, E.A.; Klaus, A.A.; Kartashov, A.V.; Kusnetsov, V.V. Distribution of Cd and other cations between the stroma and thylakoids: A quantitative approach to the search for Cd targets in chloroplasts. Photosynth. Res. 2018, 139, 337–358. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.P.; Mahajan, P.; Kaur, S.; Batish, D.R.; Kohli, R.K. Cadmium: Toxicity and tolerance in plants. Environ. Chem. Lett. 2013, 11, 229–254. [Google Scholar] [CrossRef]
- Tran, T.A.; Popova, L.P. Functions and toxicity of cadmium in plants: Recent advances and future prospects. Turk. J. Bot. 2013, 37, 1–13. [Google Scholar] [CrossRef]
- He, S.; Yang, X.; He, Z.; Baligar, V.C. Morphological and physiological responses of plants to cadmium toxicity: A review. Pedosphere 2017, 27, 421–438. [Google Scholar] [CrossRef]
- Papazoglou, E.G.; Karantounias, G.A.; Vemmos, S.N.; Bouranis, D.L. Photosynthesis and growth responses of giant reed (Arundo donax L.) to the heavy metals Cd and Ni. Environ. Int. 2005, 31, 243–249. [Google Scholar] [CrossRef]
- Hattab, S.; Dridi, B.; Chouba, L.; Kheder, M.B.; Bousetta, H. Photosynthesis and growth responses of pea Pisum sativum L. under heavy metals stress. J. Environ. Sci. 2009, 21, 1552–1556. [Google Scholar] [CrossRef]
- Pogrzeba, M.; Rusinowski, S.; Sitko, K.; Krzyżak, J.; Skalska, A.; Małkowski, E.; Ciszek, D.; Werle, S.; McCalmont, J.P.; Mos, M.; et al. Relationships between soil parameters and physiological status of Miscanthus x giganteus cultivated on soil contaminated with trace elements under NPK fertilisation vs. microbial inoculation. Environ. Pollut. 2017, 225, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Lysenko, E.A.; Klaus, A.A.; Kartashov, A.V.; Kusnetsov, V.V. Specificity of Cd, Cu, and Fe effects on barley growth, metal contents in leaves and chloroplasts, and activities of photosystem I and photosystem II. Plant Physiol. Biochem. 2020, 147, 191–204. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Xu, L.; Liu, Y.; Su, W.; Yan, J.; Xu, D. Effect of Biochar on the Growth, Photosynthesis, Antioxidant System and Cadmium Content of Mentha piperita ‘Chocolate’ and Mentha spicata in Cadmium-Contaminated Soil. Agronomy 2022, 12, 2737. [Google Scholar] [CrossRef]
- Rodriguez, E.; Santos, C.; Azevedo, R.; Moutinho-Pereira, J.; Correia, C.; Dias, M.C. Chromium (VI) induces toxicity at different photosynthetic levels in pea. Plant Physiol. Bioch. 2012, 53, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Moradi, L.; Ehsanzadeh, P. Effects of Cd on photosynthesis and growth of safflower (Carthamus tinctorius L.) genotypes. Photosynthetica 2015, 53, 506–518. [Google Scholar] [CrossRef]
- Zhang, X.F.; Zhang, X.H.; Gao, B.; Li, Z.; Xia, H.P.; Li, H.F.; Li, J. Effect of cadmium on growth, photosynthesis, mineral nutrition and metal accumulation of an energy crop, king grass (Pennisetum americanum × P. purpureum). Biomass Bioenergy 2014, 67, 179–187. [Google Scholar] [CrossRef]
- Subrahmanyam, D.; Rathore, V.S. Influence of Manganese Toxicity on Photosynthesis in Ricebean (Vigna Umbellata) Seedlings. Photosynthetica 2021, 38, 449–453. [Google Scholar] [CrossRef]
- Qin, J.; Jiang, X.; Qin, J.; Zhao, H.; Dai, M.; Liu, H.; Chen, X. Effects of Lead Pollution on Photosynthetic Characteristics and Chlorophyll Fluorescence Parameters of Different Populations of Miscanthus floridulus. Processes 2023, 11, 1562. [Google Scholar] [CrossRef]
- Strasser, R.J.; Tsimilli-Michael, M.; Qiang, S.; Goltsev, V. Simultaneous in vivo recording of prompt and delayed fluorescence and 820-nm reflection changes during drying and after rehydration of the resurrection plant Haberlea rhodopensis. Biochim. Biophys. Acta 2010, 1797, 1313–1326. [Google Scholar] [CrossRef]
- Oukarroum, A.; Goltsev, V.; Strasser, R.J. Temperature effects on pea plants probed by simultaneous measurements of the kinetics of prompt fluorescence, delayed fluorescence and modulated 820 nm reflection. PLoS ONE 2013, 8, e59433. [Google Scholar] [CrossRef]
- Gao, J.; Li, P.; Ma, F.; Goltsev, V. Photosynthetic performance during leaf expansion in Malus micromalus probed by chlorophyll a fluorescence and modulated 820 nm reflection. J. Photochem. Photobiol. B Biol. 2014, 137, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Zhang, M.; Gao, J.; Li, P.; Goltsev, V.; Ma, F. Thermotolerance of apple tree leaves probed by chlorophyll a fluorescence and modulated 820 nm reflection during seasonal shift. J. Photochem. Photobiol. B Biol. 2015, 152, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Schansker, G.; Srivastava, A.; Strasser, R.J. Characterization of the 820-nm transmission signal paralleling the chlorophyll a fluorescence rise (OJIP) in pea leaves. Funct. Plant Biol. 2003, 30, 785–796. [Google Scholar] [CrossRef] [PubMed]
- Mehta, P.; Jajoo, A.; Mathur, S.; Bharti, S. Chlorophyll a fluorescence study revealing effects of high salt stress on Photosystem II in wheat leaves. Plant Physiol. Biochem. 2010, 48, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Yang, J.; Zhang, M.; Strasser, R.J.; Qiang, S. Classification and characteristics of heat tolerance in Ageratina adenophora populations using fast chlorophyll a fluorescence rise O-J-I-P. Environ. Exp. Bot. 2016, 122, 126–140. [Google Scholar] [CrossRef]
- Dąbrowski, P.; Kalaji, M.H.; Baczewska, A.H.; Pawluśkiewicz, B.; Mastalerczuk, G.; Borawska-Jarmułowicz, B.; Paunov, M.; Goltsev, V. Delayed chlorophyll a fluorescence, MR 820, and gas exchange changes in perennial ryegrass under salt stress. J. Lumin. 2017, 183, 322–333. [Google Scholar] [CrossRef]
- Paunov, M.; Koleva, L.; Vassilev, A.; Vangronsveld, J.; Goltsev, V. Effects of different metals on photosynthesis: Cadmium and zinc affect chorophyll fluorescence in durum wheat. Int. J. Mol. Sci. 2018, 19, 787. [Google Scholar] [CrossRef] [PubMed]
- Yruela, I.; Pueyo, J.J.; Alonso, P.J.; Picorel, R. Photoinhibition of Photosystem II from Higher Plants. J. Biol. Chem. 1996, 271, 27408–27415. [Google Scholar] [CrossRef] [PubMed]
- Vassilev, A.; Lidon, F.; Scotti, P.; Da Graca, M.; Yordanov, I. Cadmium-induced Changes in Chloroplast Lipids and Photosystem Activities in Barley Plants. Biol. Plant. 2004, 48, 153–156. [Google Scholar] [CrossRef]
- Janeczko, A.; Koscielniak, J.; Pilipowicz, M.; Szarek-Lukaszewska, G.; Skoczowski, A. Protection of winter rape photosystem 2 by 24-epibrassinolide under cadmium stress. Photosynthetica 2005, 43, 293–298. [Google Scholar] [CrossRef]
- Liu, M.S.; Huang, X.H.; Wang, R.J.; Xu, H.Y.; Zhu, F. Inhibition of photosynthesis in Melia azedarach and Ligustrum lucidum induced by manganese toxicity using OJIP chlorophyll a fluorescence transient. Photosynthetica 2021, 59, 148–159. [Google Scholar] [CrossRef]
- Baszyński, T. Interference of Cd2+ in Functioning of the Photosynthetic Apparatus of Higher Plants. Acta Soc. Bot. Pol. 2014, 55, 291–304. [Google Scholar] [CrossRef]
- Xue, Z.C.; Gao, H.Y.; Zhang, L.T. Effects of Cadmium on Growth, Photosynthetic Rate and Chlorophyll Content in Leaves of Soybean Seedlings. Biol. Plant. 2013, 57, 587–590. [Google Scholar] [CrossRef]
- Kan, X.; Ren, J.; Chen, T.; Cui, M.; Li, C.; Zhou, R.; Zhang, Y.; Liu, H.; Deng, D.; Yin, Z.T. Effects of salinity on photosynthesis in maize probed by prompt fluorescence, delayed fluorescence and P700 signals. Environ. Exp. Bot. 2017, 140, 56–64. [Google Scholar] [CrossRef]
- Zhou, R.; Kan, X.; Chen, J.; Hua, H.; Li, Y.; Ren, J.; Feng, K.; Liu, H.; Deng, D.; Yin, Z. Drought-induced changes in photosynthetic electron transport in maize probed by prompt fluorescence, delayed fluorescence, P700 and cyclic electron flow signals. Environ. Exp. Bot. 2019, 158, 51–62. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembrans. Methods Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Satterfield, C.N.; Bonnell, A.H. Interferences in Titanium Sulfate Method for Hydrogen Peroxide. Anal. Chem. 1955, 27, 1174–1175. [Google Scholar] [CrossRef]
- Senthilkumar, M.; Amaresan, N.; Sankaranarayanan, A. Estimation of Malondialdehyde (MDA) by Thiobarbituric Acid (TBA) Assay. In Plant-Microbe Interactions; Springer: Berlin/Heidelberg, Germany, 2021; pp. 103–105. [Google Scholar] [CrossRef]
- Noël, L.; Dufailly, V.; Lemahieu, N.; Vastel, C.; Guérin, T. Simultaneous analysis of cadmium, lead, mercury, and arsenic content in foodstuffs of animal origin by inductively coupled plasma/mass spectrometry after closed vessel microwave digestion: Method validation. J AOAC Int. 2005, 88, 1811–1821. [Google Scholar] [CrossRef]
- Anjum, S.A.; Tanveer, M.; Hussain, S.; Shahzad, B.; Ashraf, U.; Fahad, S.; Hassan, W.; Jan, S.; Khan, L.; Saleem, M.F.; et al. Osmoregulation and antioxidant production in maize under combined cadmium and arsenic stress. Environ. Sci. Pollut. Res. Int. 2016, 23, 11864–11875. [Google Scholar] [CrossRef] [PubMed]
- Baczek-Kwinta, R.; JuzoŃ, K.; Borek, M.; Antonkiewicz, J. Photosynthetic response of cabbage in cadmium-spiked soil. Photosynthetica 2019, 57, 731–739. [Google Scholar] [CrossRef]
- Satarug, S.; Garrett, S.H.; Sens, M.A.; Sens, D.A. Cadmium, environmental exposure, and health outcomes. Environ. Health Perspect. 2010, 118, 182–190. [Google Scholar] [CrossRef]
- Liu, B.; An, C.; Jiao, S.; Jia, F.; Liu, R.; Wu, Q.; Dong, Z. Impacts of the Inoculation of Piriformospora indica on Photosynthesis, Osmoregulatory Substances, and Antioxidant Enzymes of Alfalfa Seedlings under Cadmium Stress. Agriculture 2022, 12, 1928. [Google Scholar] [CrossRef]
- Ci, D.; Jiang, D.; Wollenweber, B.; Dai, T.; Jing, Q.; Cao, W. Cadmium stress in wheat seedlings: Growth, cadmium accumulation and photosynthesis. Acta Physiol. Plant 2010, 32, 365–373. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, L.; Huang, X.; Zhou, Y.; Quan, Q.; Li, Y.; Zhu, X. Response of photosynthesis to different concentrations of heavy metals in Davidia involucrata. PLoS ONE 2020, 15, e0228563. [Google Scholar] [CrossRef] [PubMed]
- Baruah, N.; Gogoi, N.; Roy, S.; Bora, P.; Chetia, J.; Zahra, N.; Ali, N.; Gogoi, P.; Farooq, M. Phytotoxic Responses and Plant Tolerance Mechanisms to Cadmium Toxicity. J. Soil Sci. Plant Nutr. 2023, 23, 4805–4826. [Google Scholar] [CrossRef]
- Popova, A.V.; Borisova, P.; Vasilev, D. Response of Pea Plants (Pisum sativum cv. Ran 1) to NaCl Treatment in Regard to Membrane Stability and Photosynthetic Activity. Plants 2023, 12, 324. [Google Scholar] [CrossRef]
- Desoky, E.-S.M.; Merwad, A.-R.M.A.; Abo El-Maati, M.F.; Mansour, E.; Arnaout, S.M.A.I.; Awad, M.F.; Ramadan, M.F.; Ibrahim, S.A. Physiological and Biochemical Mechanisms of Exogenously Applied Selenium for Alleviating Destructive Impacts Induced by Salinity Stress in Bread Wheat. Agronomy 2021, 11, 926. [Google Scholar] [CrossRef]
- Yaghoubian, Y.; Siadat, S.A.; Moradi Telavat, M.R.; Pirdashti, H. Quantify the response of purslane plant growth, photosynthesis pigments and photosystem II photochemistry to cadmium concentration gradients in the soil. Russ. J. Plant Physiol. 2016, 63, 77–84. [Google Scholar] [CrossRef]
- Chen, X.; Tao, H.; Wu, Y.; Xu, X. Effects of Cadmium on metabolism of photosynthetic pigment and photosynthetic system in Lactuca sativa L. revealed by physiological and proteomics analysis. Sci. Hortic. 2022, 305, 111371. [Google Scholar] [CrossRef]
- Takabayashi, A.; Kurihara, K.; Kuwano, M.; Kasahara, Y.; Tanaka, R.; Tanaka, A. The Oligomeric States of the Photosystems and the Light-Harvesting Complexes in the Chl b-Less Mutant. Plant Cell Physiol. 2011, 52, 2103–2114. [Google Scholar] [CrossRef]
- Dias, M.C.; Monteiro, C.; Moutinho-Pereira, J.; Correia, C.; Goncalves, B.; Santos, C. Cadmium toxicity affects photosynthesis and plant growth at different levels. Acta Physiol. Plant. 2013, 35, 1281–1289. [Google Scholar] [CrossRef]
- Toth, S.Z.; Schansker, G.; Garab, G.; Strasser, R.J. Photosynthetic electron transport activity in heat-treated barley leaves: The role of internal alternative electron donors to photosystem II. Biochim. Biophys. Acta 2007, 1767, 295–305. [Google Scholar] [CrossRef]
- Gao, Y.; Liu, W.; Wang, X.; Yang, L.; Han, S.; Chen, S.; Strasser, R.J.; Valverde, B.E.; Qiang, S. Comparative phytotoxicity of usnic acid, salicylic acid, cinnamic acid and benzoic acid on photosynthetic apparatus of Chlamydomonas reinhardtii. Plant Physiol. Biochem. 2018, 128, 1–12. [Google Scholar] [CrossRef]
- Yamane, Y.; Kashino, Y.; Koike, H.; Satoh, K. Increases in the fluorescence Fo level and reversible inhibition of photosystem II reaction center by high-temperature treatments in higher plants. Photosynth. Res. 1997, 52, 57–64. [Google Scholar] [CrossRef]
- Oukarroum, A.; Bussotti, F.; Goltsev, V.; Kalaji, H.M. Correlation between reactive oxygen species production and photochemistry of photosystems I and II in Lemna gibba l plants under salt stress. Environ. Exp. Bot. 2015, 109, 80–88. [Google Scholar] [CrossRef]
- Oukarroum, A.; Gharous, M.E.; Goltsev, V.; Strasser, R.J. Delayed fluorescence emission as a probe for the response of photosynthetic organisms to high temperature exposure: A comparative study. J. Lumin. 2016, 180, 321–327. [Google Scholar] [CrossRef]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the chlorophyll a fluorescence transient. In Chlorophyll a fluorescence: A Signature of Photosynthesis; Papageorgiou, G.C., Ed.; Springer: Berlin/Heidelberg, Germany, 2004; pp. 321–362. [Google Scholar] [CrossRef]
- Dąbrowski, P.; Baczewska, A.H.; Bussotti, F.; Pollastrini, M.; Piekut, K.; Kowalik, W.; Wróbel, J.; Kalaji, H.M. Photosynthetic efficiency of Microcystis ssp. under salt stress. Environ. Exp. Bot. 2021, 186, 104459. [Google Scholar] [CrossRef]
- He, Y.; Lu, C.; Jiang, Z.; Sun, Y.; Liu, H.; Yin, Z. NADH dehydrogenase-like complex L subunit improves salt tolerance by enhancing photosynthetic electron transport. Plant Physiol. Biochem. 2024, 207, 108420. [Google Scholar] [CrossRef]
- Dąbrowski, P.; Baczewska, A.H.; Pawluskiewicz, B.; Paunov, M.; Alexantrov, V.; Goltsev, V.; Kalaji, M.H. Prompt chlorophyll a fluorescence as a rapid tool for diagnostic changes in PSII structure inhibited by salt stress in perennial ryegrass. J. Photochem. Photobiol. B Biol. 2016, 157, 22–31. [Google Scholar] [CrossRef]
- Dabrowski, P.; Baczewska, A.H.; Kalaji, H.M.; Goltsev, V.; Paunov, M.; Rapacz, M.; Wójcik-Jagła, M.; Pawluśkiewicz, B.; Bąba, W.; Brestic, M. Exploration of chlorophyll a fluorescence and plant gas exchange parameters as indicators of drought tolerance in perennial ryegrass. Sensors 2019, 19, 2736. [Google Scholar] [CrossRef]
- Rastogi, A.; Zivcak, M.; Tripathi, D.K.; Yadav, S.; Kalaji, H.M. Phytotoxic effect of silver nanoparticles in Triticum aestivum: Improper regulation of photosystem I activity as the reason for oxidative damage in the chloroplast. Photosynthetica 2019, 57, 209–216. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Račková, L.; Paganová, V.; Swoczyna, T.; Rusinowski, S.; Sitko, K. Can chlorophyll-a fluorescence parameters be used as bio-indicators to distinguish between drought and salinity stress in Tilia cordata Mill? Environ. Exp. Bot. 2018, 152, 149–157. [Google Scholar] [CrossRef]
- Pavlović, I.; Mlinarić, S.; Tarkowská, D.; Oklestkova, J.; Novak, O.; Lepeduš, H.; Bok, V.V.; Brkanac, S.R.; Strnad, M.; Salopek-Sondi, B. Early Brassica crops responses to salinity stress: A comparative analysis between Chinese cabbage, white cabbage, and kale. Front. Plant. Sci. 2019, 10, 450. [Google Scholar] [CrossRef]
- Chen, W.; Jia, B.; Chen, J.; Feng, Y.; Li, Y.; Chen, M.; Liu, H.; Yin, Z. Effects of Different Planting Densities on Photosynthesis in Maize Determined via Prompt Fluorescence, Delayed Fluorescence and P700 Signals. Plants 2021, 10, 276. [Google Scholar] [CrossRef]
- Gan, T.; Yin, G.; Zhao, N.; Tan, X.; Wang, Y. A Sensitive Response Index Selection for Rapid Assessment of Heavy Metals Toxicity to the Photosynthesis of Chlorella pyrenoidosa Based on Rapid Chlorophyll Fluorescence Induction Kinetics. Toxics 2023, 11, 468. [Google Scholar] [CrossRef]
- Dąbrowski, P.; Keutgen, A.J.; Keutgen, N.; Sierka, E.; Baczewska- Dąbrowska, A.H.; Mojski, J.; Pawluśkiewicz, B.; Sieczko, L.; Kalaji, A.H. Photosynthetic efficiency of perennial ryegrass (Lolium perenne L.) seedlings in response to Ni and Cd stress. Sci. Rep. 2023, 13, 5357. [Google Scholar] [CrossRef]
- Salvatori, E.; Fusaro, L.; Gottardini, E.; Pollastrini, M.; Goltsev, V.; Strasser, R.J.; Bussotti, F. Plant stress analysis: Application of prompt, delayed chlorophyll fluorescence and 820 nm modulated reflectance. Insights from independent experiments. Plant Physiol. Biochem. 2014, 85, 105–113. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Goltsev, V.; Bosa, K.; Allakhverdiev, S.L.; Strasser, R.J.; Goltsev, V. Experimental in vivo measurements of light emission in plants: A perspective dedicated to David Walker. Photosynth. Res. 2012, 114, 69–96. [Google Scholar] [CrossRef]
- Goltsev, V.; Chernev, P.; Zaharieva, I.; Lambrev, P.; Strasser, R. Kinetics of delayed chlorophyll a fluorescence registered in milliseconds time rang. Photosynth. Res. 2005, 84, 209–215. [Google Scholar] [CrossRef]
- Oukarroum, A.; Lebrihi, A.; Gharous, M.E.; Goltsev, V.; Strasser, R.J. Desiccation-induced changes of photosynthetic transport in Parmelina tiliacea (Hoffm.) Ach. analysed by simultaneous measurements of the kinetics of prompt fluorescence, delayed fluorescence and modulated 820 nm reflection. J. Lumin. 2018, 198, 302–308. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Technical Fluorescence Parameters | Meaning |
|---|---|
| FO | Minimal fluorescence yield of the dark-adapted state |
| FK | PF intensity at the K-step (0.3 ms) |
| FJ | PF intensity at the J-step (2 ms) |
| FI | PF intensity at the I-step (30 ms) |
| FM = FP | Maximal fluorescence yield of the dark-adapted state |
| Vt = (Ft − FO)/(FM − FO) | Relative variable fluorescence at time t |
| VJ = (FJ − FO)/(FM − FO) | Relative variable fluorescence at the J-step (2 ms) |
| VI = (FI − FO)/(FM − FO) | Relative variable fluorescence at the I-step (30 ms) |
| MO = 4·(F300μs − FO)/(FM − FO) | Approximated initial slope of the fluorescence transient |
| Quantum efficiencies or flux ratios | |
| TRO/ABS = φPo = 1 − FO/FM | Maximum quantum yield for primary photochemistry |
| ETO/ABS = φEo = 1 − FJ/FM | Quantum yield of the electron transport flux from QA to QB |
| REO/ABS = φRo = 1 − FI/FM | Quantum yield of the electron transport flux until the PSI electron acceptors |
| ETO/TRO = ψEo = 1 − VJ | The efficiency of electron movement at QA |
| REO/TRO = ψRo = 1 − VI | Efficiency with which a PSII trapped electron is transferred until PSI acceptors |
| REO/ETO = δRo = (1 − VI)/(1 − VJ) | The efficiency of an electron beyond QA reduced PSI acceptors |
| Specific energy fluxes [per QA-reducing PSII reaction center (RC)] | |
| ABS/RC = MO·(1/VJ)/(1/φPo) | Absorption flux per RC |
| TRO/RC = MO·(1/VJ) | Trapped energy flux per RC (at t = 0) |
| ETO/RC = MO·(1/VJ)·ψEo | Electron transport flux per RC (at t = 0) |
| DIO/RC = (ABS/RC) − (TRO/RC) | Dissipated energy flux per RC (at t = 0) |
| Phenomenological energy fluxes [per excited cross-section (CS)] | |
| ABS/CSO ≈ FO | Absorption flux per CS (at t = 0) |
| TRO/CSO = φPo·(ABS/CSO) | Trapped energy flux per CS (at t = 0) |
| ETO/CSO = φEo·(ABS/CSO) | Electron transport flux per CS (at t = 0) |
| DIO/CSO = (ABS/CSO) − (TRO/CSO) | Dissipated energy flux per CS (at t = 0) |
| Density of reaction centers | |
| RC/CSO = φPo·(VJ/MO)·(ABS/CSO) | Density of RCs (QA-reducing PSII reaction centers) |
| Performance indexes | |
| PIABS = (RC/ABS)·[φPo/(1 − φPo)]·[ψEo/(1 − ψEo)] | Performance index (potential) for energy conservation from photons absorbed by PSII to the reduction of intersystem electron acceptors |
| Treatment Duration in Days | VPSI | VPSII−PSI | VPSII |
|---|---|---|---|
| 0 d | 0.616 ± 0.097 a | 0.041 ± 0.004 a | 0.657 ± 0.101 a |
| 3 d | 0.470 ± 0.141 b | 0.029 ± 0.012 b | 0.499 ± 0.152 b |
| 6 d | 0.342 ± 0.022 c | 0.021 ± 0.002 c | 0.363 ± 0.023 c |
| 9 d | 0.300 ± 0.028 c | 0.015 ± 0.001 d | 0.315 ± 0.029 c |
| 12 d | 0.160 ± 0.081 d | 0.012 ± 0.004 e | 0.171 ± 0.085 d |
| 15 d | 0.111 ± 0.061 e | 0.000 ± 0.003 f | 0.068 ± 0.064 e |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, R.; Xu, J.; Li, L.; Yin, Y.; Xue, B.; Li, J.; Sun, F. Exploration of the Effects of Cadmium Stress on Photosynthesis in Oenanthe javanica (Blume) DC. Toxics 2024, 12, 307. https://doi.org/10.3390/toxics12050307
Zhou R, Xu J, Li L, Yin Y, Xue B, Li J, Sun F. Exploration of the Effects of Cadmium Stress on Photosynthesis in Oenanthe javanica (Blume) DC. Toxics. 2024; 12(5):307. https://doi.org/10.3390/toxics12050307
Chicago/Turabian StyleZhou, Ronghua, Jun Xu, Liangjun Li, Yulai Yin, Bowen Xue, Jingjing Li, and Fangfang Sun. 2024. "Exploration of the Effects of Cadmium Stress on Photosynthesis in Oenanthe javanica (Blume) DC." Toxics 12, no. 5: 307. https://doi.org/10.3390/toxics12050307
APA StyleZhou, R., Xu, J., Li, L., Yin, Y., Xue, B., Li, J., & Sun, F. (2024). Exploration of the Effects of Cadmium Stress on Photosynthesis in Oenanthe javanica (Blume) DC. Toxics, 12(5), 307. https://doi.org/10.3390/toxics12050307