
Role of Macrophages in Cytotoxicity, Reactive Oxygen Species Production and DNA Damage in 1,2-Dichloropropane-Exposed Human Cholangiocytes In Vitro

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Cell Cultures
2.2. Co-Culture Method
2.3. Determination of 1,2-DCP Concentrations
2.4. Preparation of 1,2-DCP Exposure Solution
2.5. Cell Exposure to 1,2-DCP
2.6. Assessment of Cell Proliferation
2.7. MTS Assay
2.8. BrdU Assay
2.9. Cell Count
2.10. LDH Cytotoxicity Assay
2.11. γH2AX Immunofluorescence
2.12. Alkaline Comet Assay
2.13. ROS Detection
2.14. ELISA for Proinflammatory Cytokines
2.15. Statistical Analysis
3. Results
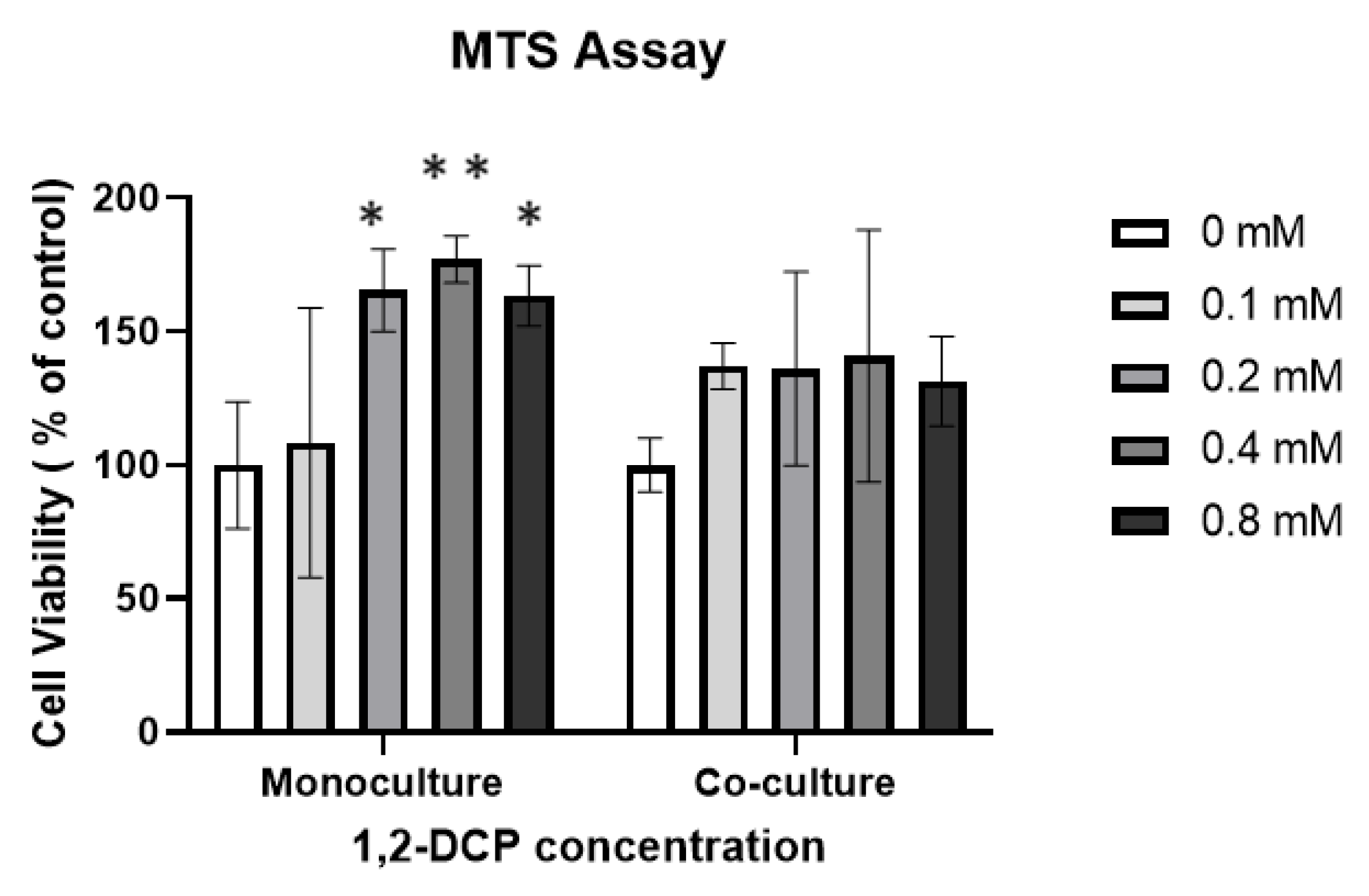
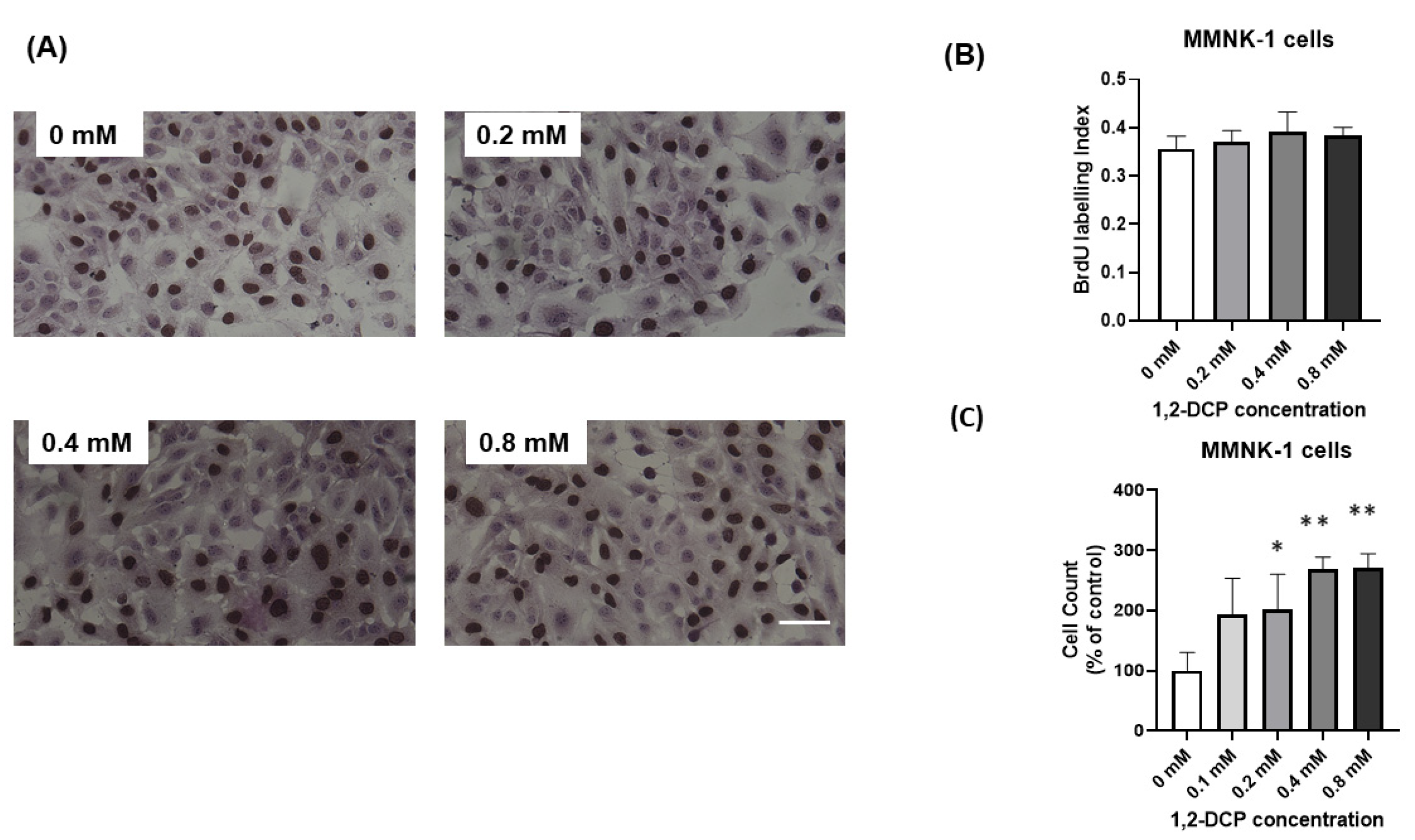
3.1. 1,2-DCP Increases Cell Proliferation of Monocultured Cells but Not Co-Cultured Cells
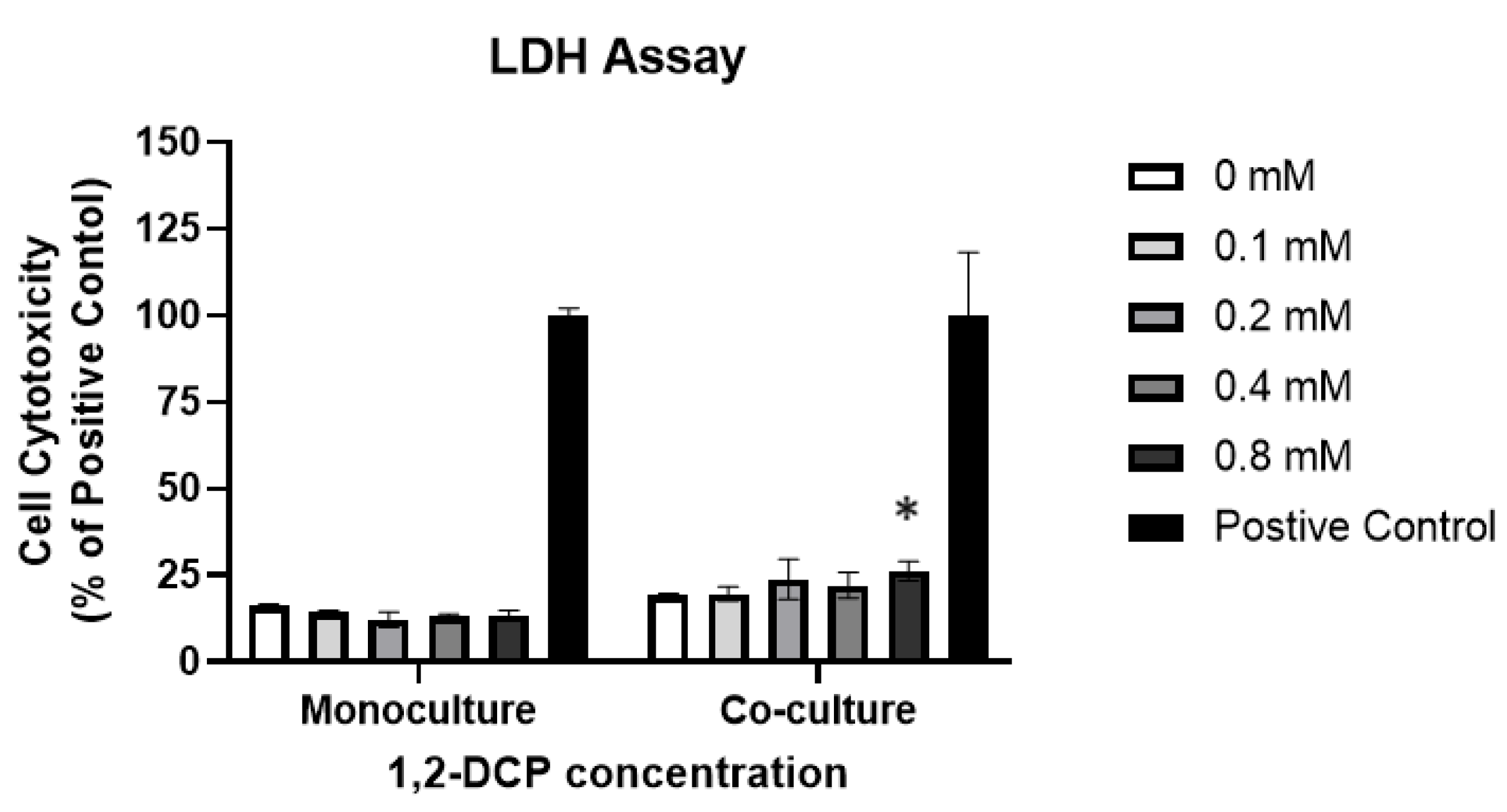
3.2. 1,2-DCP Is Cytotoxic to Co-Cultured Cells but Not Monocultured Cells
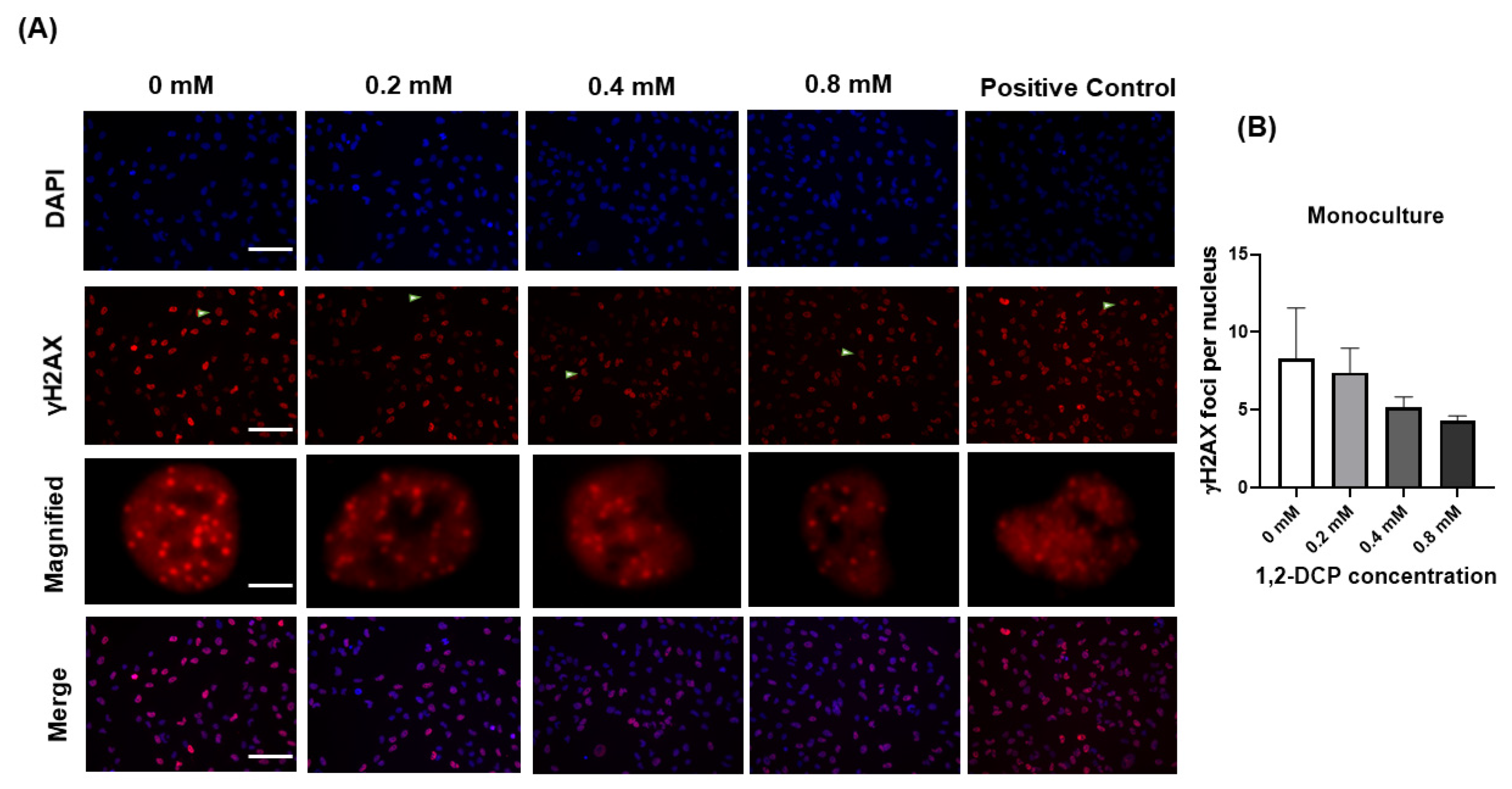
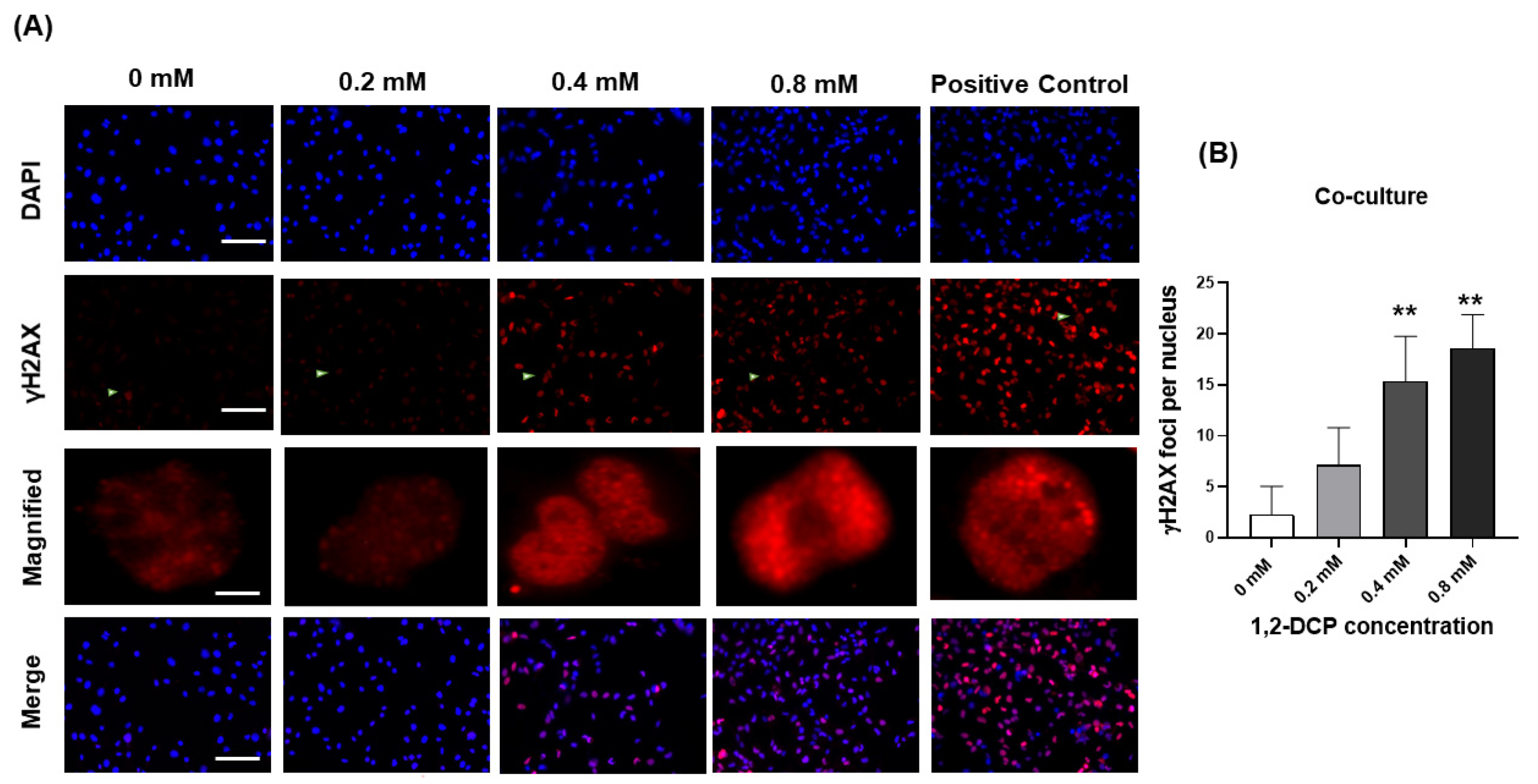
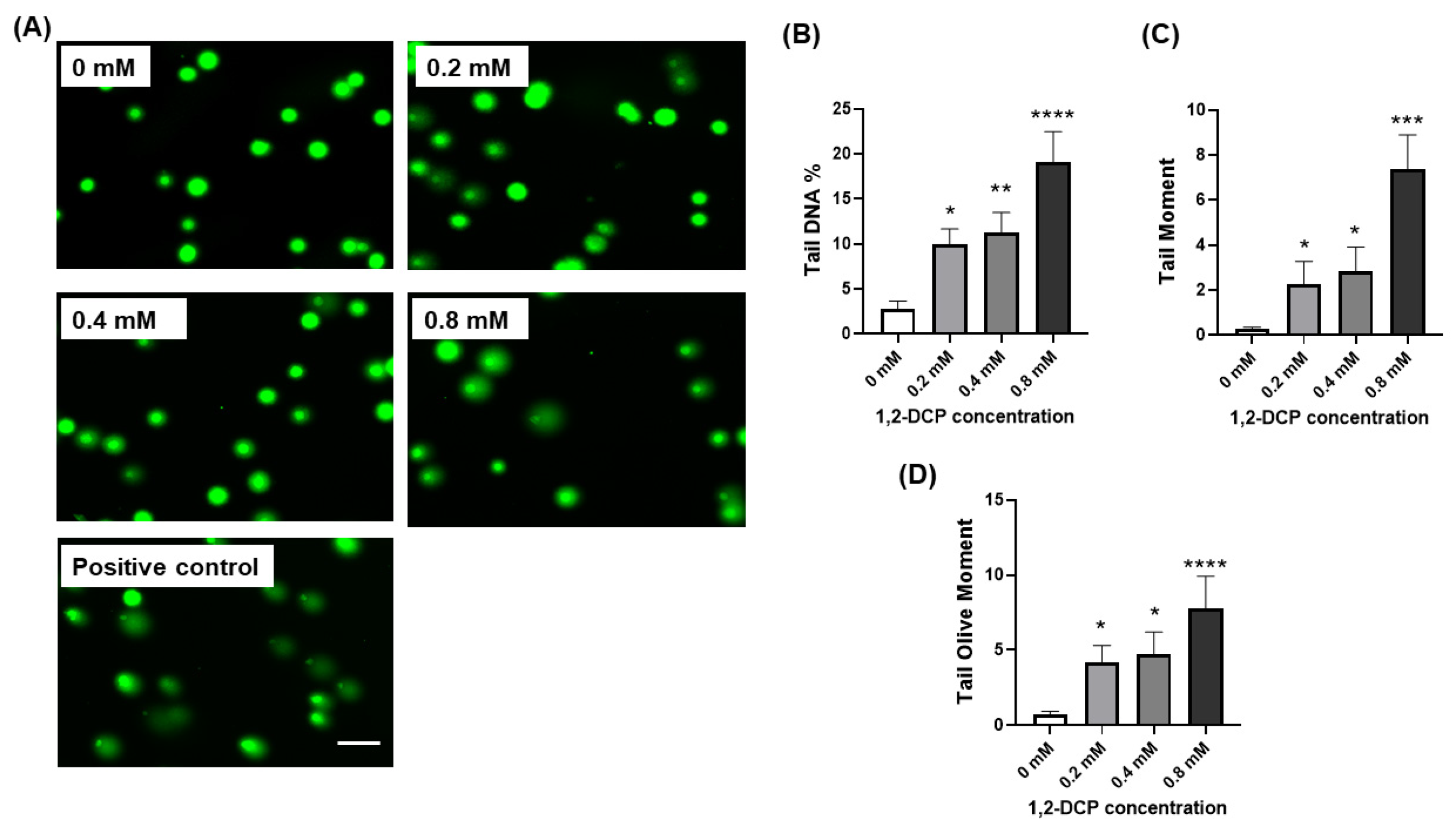
3.3. 1,2-DCP Causes DNA Damage in Co-Cultured Cells but Not Monocultured Cells
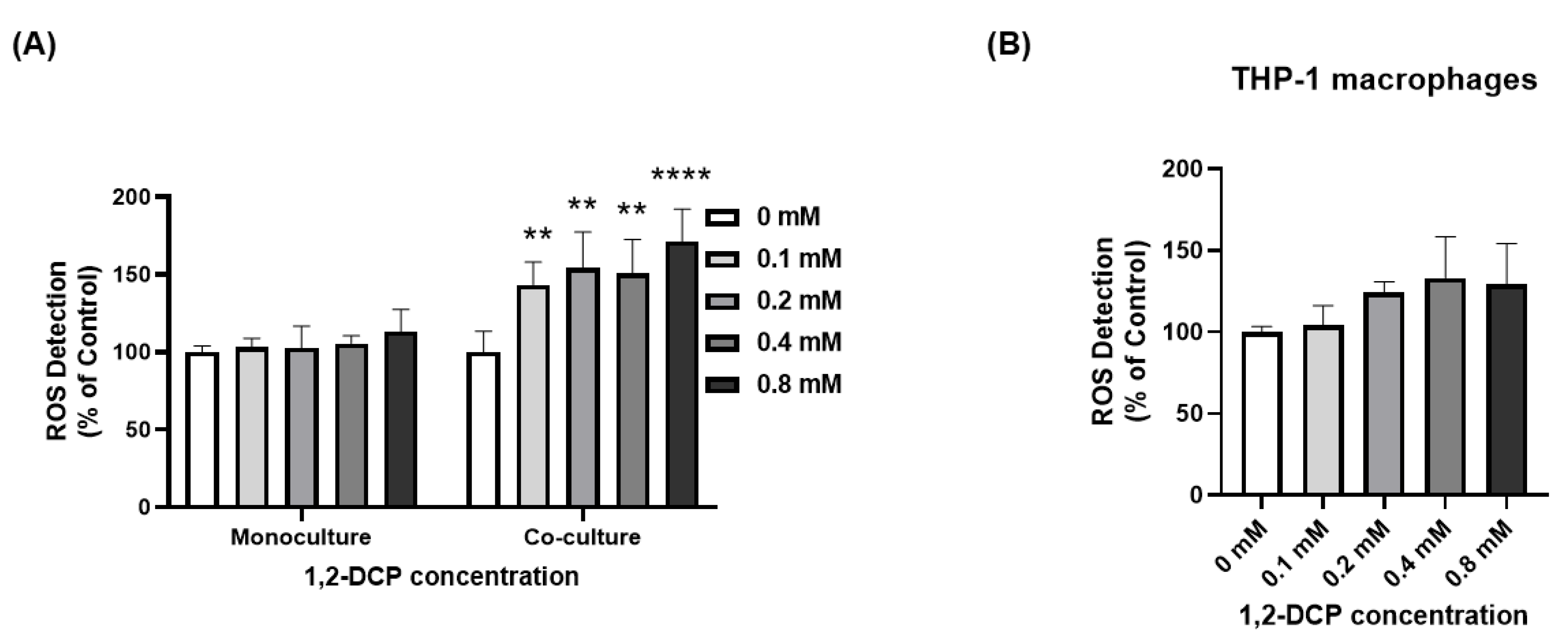
3.4. 1,2-DCP Enhances ROS Production in Co-Cultured Cells but Not Monocultured Cells
3.5. 1,2-DCP Induces TNF-α and IL-1β in THP-1 Macrophages
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- ECHA. 1,2-Dichloropropane- Substance Information. 2016. Available online: https://echa.europa.eu/substance-information/-/substanceinfo/100.001.048 (accessed on 11 November 2020).
- IARC. 1,2-Dichloropropane-IARC Monographs. Some Chemicals Used as Solvents and in Polymer Manufacture. IARC Monogr. Eval. Carcinog Risks Hum. 2017, 110, 142–175. [Google Scholar]
- Kumagai, S.; Kurumatani, N.; Arimoto, A.; Ichihara, G. Cholangiocarcinoma among offset colour proof-printing workers exposed to 1,2-dichloropropane and/or dichloromethane. Occup. Environ. Med. 2013, 70, 508–510. [Google Scholar] [CrossRef] [PubMed]
- Kubo, S.; Nakanuma, Y.; Takemura, S.; Sakata, C.; Urata, Y.; Nozawa, A.; Nishioka, T.; Kinoshita, M.; Hamano, G.; Terajima, H.; et al. Case series of 17 patients with cholangiocarcinoma among young adult workers of a printing company in Japan. J. Hepato-Biliary-Pancreat. Sci. 2014, 21, 479–488. [Google Scholar] [CrossRef] [PubMed]
- Sobue, T.; Utada, M.; Makiuchi, T.; Ohno, Y.; Uehara, S.; Hayashi, T.; Sato, K.K.; Endo, G. Risk of bile duct cancer among printing workers exposed to 1,2-dichloropropane and/or dichloromethane. J. Occup. Health 2015, 57, 230–236. [Google Scholar] [CrossRef] [Green Version]
- Kumagai, S. Two offset printing workers with cholangiocarcinoma. J. Occup. Health 2014, 56, 164–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, K.; Kumagai, S.; Nagoya, T.; Endo, G. Chemical exposure levels in printing workers with cholangiocarcinoma. J. Occup. Health 2014, 56, 332–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ministry of Health, Labour and Welfare of Japan. Available online: http://www.mhlw.go.jp/english/policy/employ-labour/labour-standards/Occupational.html (accessed on 15 November 2020).
- Sato, Y.; Kubo, S.; Takemura, S.; Sugawara, Y.; Tanaka, S.; Fujikawa, M.; Arimoto, A.; Harada, K.; Sasaki, M.; Nakanuma, Y. Different carcinogenic process in cholangiocarcinoma cases epidemically developing among workers of a printing company in Japan. Int. J. Clin. Exp. Pathol. 2014, 7, 4745–4754. [Google Scholar] [PubMed]
- Mimaki, S.; Totsuka, Y.; Suzuki, Y.; Nakai, C.; Goto, M.; Kojima, M.; Arakawa, H.; Takemura, S.; Tanaka, S.; Marubashi, S.; et al. Hypermutation and unique mutational signatures of occupational cholangiocarcinoma in printing workers exposed to haloalkanes. Carcinogenesis 2016, 37, 817–826. [Google Scholar] [CrossRef] [Green Version]
- Gi, M.; Fujioka, M.; Yamano, S.; Shimomura, E.; Kanki, M.; Kawachi, S.; Tachibana, H.; Tatsumi, K.; Fang, H.; Ishii, N.; et al. Modifying effects of 1,2-dichloropropane on N-nitrosobis(2-oxopropyl)amine-induced cholangiocarcinogenesis in male Syrian hamsters. J. Toxicol. Sci. 2015, 40, 647–656. [Google Scholar] [CrossRef] [Green Version]
- NTP. Toxicology and Carcinogenesis Studies of 1,2-Dichloropropane (Propylene Dichloride) (CAS No. 78-87-5) in F344/N Rats and B6C3F1 Mice (Gavage Studies). Natl. Toxicol. Program Tech. Rep. Ser. 1986, 263, 1–182. [Google Scholar]
- Umeda, Y.; Matsumoto, M.; Aiso, S.; Nishizawa, T.; Nagano, K.; Arito, H.; Fukushima, S. Inhalation carcinogenicity and toxicity of 1,2-dichloropropane in rats. Inhal. Toxicol. 2010, 22, 1116–1126. [Google Scholar] [CrossRef]
- Guengerich, F.P.; Peterson, L.A.; Cmarik, J.L.; Koga, N.; Inskeep, P.B. Activation of Dihaloalkanes by Glutathione Conjugation and Formation of DNA Adducts. Environ. Health Perspect. 1987, 76, 15–18. [Google Scholar] [CrossRef] [PubMed]
- Bartels, M.J.; Timchalk, C. 1,2-Dichloropropane: Investigation of the mechanism of mercapturic acid formation in the rat. Xenobiotica Fate Foreign Compd. Biol. Syst. 1990, 20, 1035–1042. [Google Scholar] [CrossRef]
- De Lorenzo, F.; Degl’Innocenti, S.; Ruocco, A.; Silengo, L.; Cortese, R. Mutagenicity of pesticides containing 1,3-dichloropropene. Cancer Res. 1977, 37, 1915–1917. [Google Scholar] [PubMed]
- Galloway, S.M.; Armstrong, M.J.; Reuben, C.; Colman, S.; Brown, B.; Cannon, C.; Bloom, A.D.; Nakamura, F.; Ahmed, M.; Duk, S.; et al. Chromosome aberrations and sister chromatid exchanges in Chinese hamster ovary cells: Evaluations of 108 chemicals. Environ. Mol. Mutagenesis 1987, 10 (Suppl. 10), 1–175. [Google Scholar] [CrossRef]
- Hirata, T.; Cho, Y.M.; Toyoda, T.; Akagi, J.I.; Suzuki, I.; Nishikawa, A.; Ogawa, K. Lack of in vivo mutagenicity of 1,2-dichloropropane and dichloromethane in the livers of gpt delta rats administered singly or in combination. J. Appl. Toxicol. 2017, 37, 683–691. [Google Scholar] [CrossRef]
- Principe, P.; Dogliotti, E.; Bignami, M.; Crebelli, R.; Falcone, E.; Fabrizi, M.; Conti, G.; Comba, P. Mutagenicity of chemicals of industry and agricultural relevance in Salmonella, Streptomyces and Aspergillus. J. Sci. Food Agric. 1981, 32, 826–832. [Google Scholar] [CrossRef]
- Suzuki, T.; Yanagiba, Y.; Suda, M.; Wang, R.S. Assessment of the genotoxicity of 1,2-dichloropropane and dichloromethane after individual and co-exposure by inhalation in mice. J. Occup. Health 2014, 56, 205–214. [Google Scholar] [CrossRef] [Green Version]
- Haworth, S.; Lawlor, T.; Mortelmans, K.; Speck, W.; Zeiger, E. Salmonella mutagenicity test results for 250 chemicals. Environ. Mutagenesis 1983, 5 (Suppl. 1), 1–142. [Google Scholar] [CrossRef]
- EPA. Final Study Report: Propylene Dichloride: Dominant Lethal Study in Sprague-Dawley Rats (Final) with Cover Letter. 1989. Available online: https://ntrl.ntis.gov/NTRL/dashboard/searchResults/titleDetail/OTS0527736.xhtmlarchived (accessed on 5 December 2020).
- Trush, M.A.; Kensler, T.W. An overview of the relationship between oxidative stress and chemical carcinogenesis. Free Radic. Biol. Med. 1991, 10, 201–209. [Google Scholar] [CrossRef]
- Laskin, D.L. Macrophages and inflammatory mediators in chemical toxicity: A battle of forces. Chem. Res. Toxicol. 2009, 22, 1376–1385. [Google Scholar] [CrossRef] [Green Version]
- Laskin, D.L.; Pendino, K.J. Macrophages and inflammatory mediators in tissue injury. Annu. Rev. Pharmacol. Toxicol. 1995, 35, 655–677. [Google Scholar] [CrossRef]
- Bruckner, J.V.; MacKenzie, W.F.; Ramanathan, R.; Muralidhara, S.; Kim, H.J.; Dallas, C.E. Oral toxicity of 1,2-dichloropropane: Acute, short-term, and long-term studies in rats. Fundam. Appl. Toxicol. Off. J. Soc. Toxicol. 1989, 12, 713–730. [Google Scholar] [CrossRef]
- Zong, C.; Kimura, Y.; Kinoshita, K.; Takasu, S.; Zhang, X.; Sakurai, T.; Sekido, Y.; Ichihara, S.; Endo, G.; Ichihara, G. Exposure to 1,2-Dichloropropane Upregulates the Expression of Activation-Induced Cytidine Deaminase (AID) in Human Cholangiocytes Co-Cultured With Macrophages. Toxicol. Sci. Off. J. Soc. Toxicol. 2019, 168, 137–148. [Google Scholar] [CrossRef]
- Oliveira, P.A.; Colaço, A.; Chaves, R.; Guedes-Pinto, H.; De-La-Cruz, P.L.; Lopes, C. Chemical carcinogenesis. Ann. Acad. Bras. Cienc. 2007, 79, 593–616. [Google Scholar] [CrossRef] [Green Version]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eastmond, D.A.; Smith, M.T. Xenobiotic activation by stimulated human polymorphonuclear leukocytes and myeloperoxidase. Methods Enzymol. 1990, 186, 579–585. [Google Scholar] [CrossRef]
- Trush, M.A.; Seed, J.L.; Kensler, T.W. Oxidant-dependent metabolic activation of polycyclic aromatic hydrocarbons by phorbol ester-stimulated human polymorphonuclear leukocytes: Possible link between inflammation and cancer. Proc. Natl. Acad. Sci. USA 1985, 82, 5194–5198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Twerdok, L.E.; Trush, M.A. Neutrophil-derived oxidants as mediators of chemical activation in bone marrow. Chem. Biol. Interact. 1988, 65, 261–273. [Google Scholar] [CrossRef]
- Parke, D.V.; Sapota, A. Chemical toxicity and reactive oxygen species. Int. J. Occup. Med. Environ. Health 1996, 9, 331–340. [Google Scholar]
- Klaunig, J.E.; Kamendulis, L.M.; Xu, Y. Epigenetic mechanisms of chemical carcinogenesis. Hum. Exp. Toxicol. 2000, 19, 543–555. [Google Scholar] [CrossRef]
- Klaunig, J.E.; Wang, Z. Oxidative stress in carcinogenesis. Curr. Opin. Toxicol. 2018, 7, 116–121. [Google Scholar] [CrossRef]
- Maruyama, M.; Kobayashi, N.; Westerman, K.A.; Sakaguchi, M.; Allain, J.E.; Totsugawa, T.; Okitsu, T.; Fukazawa, T.; Weber, A.; Stolz, D.B.; et al. Establishment of a highly differentiated immortalized human cholangiocyte cell line with SV40T and hTERT. Transplantation 2004, 77, 446–451. [Google Scholar] [CrossRef] [Green Version]
- Tsuchiya, S.; Yamabe, M.; Yamaguchi, Y.; Kobayashi, Y.; Konno, T.; Tada, K. Establishment and characterization of a human acute monocytic leukemia cell line (THP-1). Int. J. Cancer 1980, 26, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Tada-Oikawa, S.; Ichihara, G.; Yabata, M.; Izuoka, K.; Suzuki, M.; Sakai, K.; Ichihara, S. Zinc oxide nanoparticles induce migration and adhesion of monocytes to endothelial cells and accelerate foam cell formation. Toxicol. Appl. Pharmacol. 2014, 278, 16–25. [Google Scholar] [CrossRef]
- Sato, A.; Nakajima, T. A structure-activity relationship of some chlorinated hydrocarbons. Arch. Environ. Health 1979, 34, 69–75. [Google Scholar] [CrossRef]
- Malich, G.; Markovic, B.; Winder, C. The sensitivity and specificity of the MTS tetrazolium assay for detecting the in vitro cytotoxicity of 20 chemicals using human cell lines. Toxicology 1997, 124, 179–192. [Google Scholar] [CrossRef]
- Mead, T.J.; Lefebvre, V. Proliferation assays (BrdU and EdU) on skeletal tissue sections. Methods Mol. Biol. 2014, 1130, 233–243. [Google Scholar] [CrossRef] [Green Version]
- Tolnai, S. A method for viable cell count. Tca Man. Tissue Cult. Assoc. 1975, 1, 37–38. [Google Scholar] [CrossRef]
- Kaja, S.; Payne, A.J.; Naumchuk, Y.; Koulen, P. Quantification of Lactate Dehydrogenase for Cell Viability Testing Using Cell Lines and Primary Cultured Astrocytes. Curr. Protoc. Toxicol. 2017, 72, 21–26. [Google Scholar] [CrossRef] [Green Version]
- Au-Popp, H.D.; Au-Brendel, S.; Au-Hofmann, W.-K.; Au-Fabarius, A. Immunofluorescence Microscopy of gamma H2AX and 53BP1 for Analyzing the Formation and Repair of DNA Double-strand Breaks. J. Vis. Exp. 2017, 129, e56617. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Lu, Y.; Liu, Y.; Yang, C. Evaluating In Vitro DNA Damage Using Comet Assay. J. Vis. Exp. 2017, 128, 450. [Google Scholar] [CrossRef] [PubMed]
- Zeineldin, M.; Federico, S.; Chen, X.; Fan, Y.; Xu, B.; Stewart, E.; Zhou, X.; Jeon, J.; Griffiths, L.; Nguyen, R.; et al. MYCN amplification and ATRX mutations are incompatible in neuroblastoma. Nat. Commun. 2020, 11, 913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, K.; Li, W.; Chen, Y.; Zhu, J. Effect of PM2.5 on invasion and proliferation of HeLa cells and the expression of inflammatory cytokines IL-1 and IL-6. Oncol. Lett. 2018, 16, 7068–7073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imberti, R.; Mapelli, A.; Colombo, P.; Richelmi, P.; Bertè, F.; Bellomo, G. 1,2-Dichloropropane (DCP) toxicity is correlated with DCP-induced glutathione (GSH) depletion and is modulated by factors affecting intracellular GSH. Arch. Toxicol. 1990, 64, 459–465. [Google Scholar] [CrossRef]
- Brandt, R.; Keston, A.S. Synthesis of diacetyldichlorofluorescin: A stable reagent for fluorometric analysis. Anal. Biochem. 1965, 11, 6–9. [Google Scholar] [CrossRef]
- Tacke, F.; Zimmermann, H.W. Macrophage heterogeneity in liver injury and fibrosis. J. Hepatol. 2014, 60, 1090–1096. [Google Scholar] [CrossRef] [Green Version]
- Romar, G.A.; Kupper, T.S.; Divito, S.J. Research Techniques Made Simple: Techniques to Assess Cell Proliferation. J. Investig. Dermatol. 2016, 136, e1–e7. [Google Scholar] [CrossRef] [Green Version]
- Cory, A.H.; Owen, T.C.; Barltrop, J.A.; Cory, J.G. Use of an aqueous soluble tetrazolium/formazan assay for cell growth assays in culture. Cancer Commun. 1991, 3, 207–212. [Google Scholar] [CrossRef]
- Frowein, J. Hypothesis: Chemical carcinogenesis mediated by a transiently active carcinogen receptor. Cytogenet. Cell Genet. 2000, 91, 102–104. [Google Scholar] [CrossRef]
- Trosko, J.E. Commentary: Is the concept of “tumor promotion” a useful paradigm? Mol. Carcinog. 2001, 30, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.M. Analysis of modifying factors in chemical carcinogenesis. Prog. Exp. Tumor Res. 1991, 33, 21–40. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zong, C.; Zhang, L.; Garner, E.; Sugie, S.; Huang, C.; Wu, W.; Chang, J.; Sakurai, T.; Kato, M.; et al. Exposure of Mice to 1,2-Dichloropropane Induces CYP450-Dependent Proliferation and Apoptosis of Cholangiocytes. Toxicol. Sci. Off. J. Soc. Toxicol. 2018, 162, 559–569. [Google Scholar] [CrossRef]
- Takizawa, R.; Zong, C.; Kinoshita, K.; Sakurai, T.; Ichihara, S.; Ichihara, G. 1,2-dichloropropane induces γ-H2AX expression in human cholangiocytes only in the presence of macrophages. Toxicol. Sci. Off. J. Soc. Toxicol. 2021, 180, 73. [Google Scholar]
- Mah, L.J.; El-Osta, A.; Karagiannis, T.C. gammaH2AX: A sensitive molecular marker of DNA damage and repair. Leukemia 2010, 24, 679–686. [Google Scholar] [CrossRef] [Green Version]
- Podhorecka, M.; Skladanowski, A.; Bozko, P. H2AX Phosphorylation: Its Role in DNA Damage Response and Cancer Therapy. J. Nucleic Acids 2010, 2010, 161. [Google Scholar] [CrossRef] [Green Version]
- Solomon, E.; Borrow, J.; Goddard, A.D. Chromosome aberrations and cancer. Science 1991, 254, 1153–1160. [Google Scholar] [CrossRef]
- Toyooka, T.; Yanagiba, Y.; Suda, M.; Ibuki, Y.; Wang, R.S. 1,2-Dichloropropane generates phosphorylated histone H2AX via cytochrome P450 2E1-mediated metabolism. Toxicol. Lett. 2017, 272, 60–67. [Google Scholar] [CrossRef]
- Sinha, K.; Das, J.; Pal, P.B.; Sil, P.C. Oxidative stress: The mitochondria-dependent and mitochondria-independent pathways of apoptosis. Arch. Toxicol. 2013, 87, 1157–1180. [Google Scholar] [CrossRef]
- Biswas, S.K. Does the Interdependence between Oxidative Stress and Inflammation Explain the Antioxidant Paradox? Oxidative Med. Cell. Longev. 2016, 2016, 5698931. [Google Scholar] [CrossRef] [Green Version]
- Arango, D.G.; Descoteaux, A. Macrophage cytokines: Involvement in immunity and infectious diseases. Front. Immunol. 2014, 5, 491. [Google Scholar] [CrossRef] [Green Version]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.; Elner, S.G.; Bian, Z.M.; Till, G.O.; Petty, H.R.; Elner, V.M. Pro-inflammatory cytokines increase reactive oxygen species through mitochondria and NADPH oxidase in cultured RPE cells. Exp. Eye Res. 2007, 85, 462–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goossens, V.; Grooten, J.; De Vos, K.; Fiers, W. Direct evidence for tumor necrosis factor-induced mitochondrial reactive oxygen intermediates and their involvement in cytotoxicity. Proc. Natl. Acad. Sci. USA 1995, 92, 8115–8119. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.J.; Lee, S.B.; Park, J.K.; Yoo, Y.D. TNF-alpha-induced ROS production triggering apoptosis is directly linked to Romo1 and Bcl-X(L). Cell Death Differ. 2010, 17, 1420–1434. [Google Scholar] [CrossRef] [Green Version]
- Brigelius-Flohé, R.; Banning, A.; Kny, M.; Böl, G.F. Redox events in interleukin-1 signaling. Arch. Biochem. Biophys. 2004, 423, 66–73. [Google Scholar] [CrossRef]
- Koop, D.R. Oxidative and reductive metabolism by cytochrome P450 2E1. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 1992, 6, 724–730. [Google Scholar] [CrossRef] [PubMed]
- Linhart, K.; Bartsch, H.; Seitz, H.K. The role of reactive oxygen species (ROS) and cytochrome P-450 2E1 in the generation of carcinogenic etheno-DNA adducts. Redox Biol. 2014, 3, 56–62. [Google Scholar] [CrossRef] [Green Version]
- Yanagiba, Y.; Suzuki, T.; Suda, M.; Hojo, R.; Gonzalez, F.J.; Nakajima, T.; Wang, R.S. Cytochrome P450 2E1 is responsible for the initiation of 1,2-dichloropropane-induced liver damage. Toxicol. Ind. Health 2016, 32, 1589–1597. [Google Scholar] [CrossRef]
- Wang, H.; Chen, J.; Suda, M.; Yanagiba, Y.; Weng, Z.; Wang, R.S. Acute inhalation co-exposure to 1,2-dichloropropane and dichloromethane cause liver damage by inhibiting mitochondrial respiration and defense ability in mice. J. Appl. Toxicol. 2019, 39, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Toyoda, Y.; Takada, T.; Suzuki, H. Spontaneous Production of Glutathione-Conjugated Forms of 1,2-Dichloropropane: Comparative Study on Metabolic Activation Processes of Dihaloalkanes Associated with Occupational Cholangiocarcinoma. Oxidative Med. Cell. Longev. 2017, 2017, 9736836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sesti, F.; Tsitsilonis, O.E.; Kotsinas, A.; Trougakos, I.P. Oxidative stress-mediated biomolecular damage and inflammation in tumorigenesis. In Vivo 2012, 26, 395–402. [Google Scholar]
- Bernadotte, A.; Mikhelson, V.M.; Spivak, I.M. Markers of cellular senescence. Telomere shortening as a marker of cellular senescence. Aging 2016, 8, 3–11. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Research Methods | Cells Seeded (Cells/Well) | |||
|---|---|---|---|---|
| Monoculture | Co-Culture | Monoculture | ||
| MMNK-1 Cells | MMNK-1 Cells | THP-1 Cells | THP-1 Cells | |
| MTS Assay | 1 × 104 (96-well plate) | 3 × 104 (24-well plate) | 6 × 104 (24-well inserts) | - |
| BrdU | 6 × 104 (12-well plate) | - | - | - |
| Cell count | 3 × 104 (24-well plate) | - | - | - |
| LDH Assay | 5 × 103 (96-well plate) | 3 × 104 (24-well plate) | 6 × 104 (24-well inserts) | - |
| ROS detection | 1.3 × 104 (96-well plate) | 1 × 105 (24-well plate) | 2 × 105 (24-well inserts) | 3 × 104 (96-well plate) |
| γH2AX | 2.5 × 104 (24-well plate) | 2.5 × 104 (24-well plate) | 5 × 104 (24-well inserts) | - |
| Comet Assay | - | 1.5 × 105 (6-well plate) | 3 × 105 (6-well inserts) | - |
| ELISA | - | - | - | 2 × 105 (96-well plate) |
| Model | 1,2-DCP Concentration (mM) | Simple Regression | Multiple Regression (p Value) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 0.1 | 0.2 | 0.4 | 0.8 | Effect of 1,2-DCP (p Value) | 1,2-DCP and Macrophage Interaction | Effect of 1,2-DCP | Macrophage Factor | ||
| ROS production (%) | Monocultures | 100 ± 4 | 103 ± 6 | 103 ± 14 | 106 ± 5 | 113 ± 14 | 16 (0.065) | 51 (0.03) | - | - |
| Co-cultures | 100 ± 9 | 143 ± 4 * | 154 ± 3 * | 151 ± 1 * | 171 ± 2 * | 67 (0.007) | ||||
| MTS Assay (%) | Monocultures | 100 ± 21 | 108 ± 45 | 165 ± 14 * | 177 ± 8 * | 163 ± 10 * | 78 (0.03) | −56 (0.18) | 78 (0.012) | −14 (0.24) |
| Co-cultures | 100 ± 6 | 137 ± 4 | 136 ± 17 | 141 ± 22 | 131 ± 8 | 22 (0.42) | ||||
| LDH Assay (%) | Monocultures | 16 ± 3 | 14 ± 3 | 12 ± 14 | 13 ± 3 | 14 ± 7 | −2 (0.20) | 10 (0.006) | - | - |
| Co-cultures | 19 ± 1 | 20 ± 2 | 24 ± 6 | 22 ± 4 | 26 ± 3 * | 8 (0.018) | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ekuban, A.; Zong, C.; Ekuban, F.A.; Kimura, Y.; Takizawa, R.; Morikawa, K.; Kinoshita, K.; Ichihara, S.; Ohsako, S.; Ichihara, G. Role of Macrophages in Cytotoxicity, Reactive Oxygen Species Production and DNA Damage in 1,2-Dichloropropane-Exposed Human Cholangiocytes In Vitro. Toxics 2021, 9, 128. https://doi.org/10.3390/toxics9060128
Ekuban A, Zong C, Ekuban FA, Kimura Y, Takizawa R, Morikawa K, Kinoshita K, Ichihara S, Ohsako S, Ichihara G. Role of Macrophages in Cytotoxicity, Reactive Oxygen Species Production and DNA Damage in 1,2-Dichloropropane-Exposed Human Cholangiocytes In Vitro. Toxics. 2021; 9(6):128. https://doi.org/10.3390/toxics9060128
Chicago/Turabian StyleEkuban, Abigail, Cai Zong, Frederick Adams Ekuban, Yusuke Kimura, Ryoya Takizawa, Kota Morikawa, Kazuo Kinoshita, Sahoko Ichihara, Seiichiroh Ohsako, and Gaku Ichihara. 2021. "Role of Macrophages in Cytotoxicity, Reactive Oxygen Species Production and DNA Damage in 1,2-Dichloropropane-Exposed Human Cholangiocytes In Vitro" Toxics 9, no. 6: 128. https://doi.org/10.3390/toxics9060128
APA StyleEkuban, A., Zong, C., Ekuban, F. A., Kimura, Y., Takizawa, R., Morikawa, K., Kinoshita, K., Ichihara, S., Ohsako, S., & Ichihara, G. (2021). Role of Macrophages in Cytotoxicity, Reactive Oxygen Species Production and DNA Damage in 1,2-Dichloropropane-Exposed Human Cholangiocytes In Vitro. Toxics, 9(6), 128. https://doi.org/10.3390/toxics9060128