Protein Targets of Frankincense: A Reverse Docking Analysis of Terpenoids from Boswellia Oleo-Gum Resins



Abstract
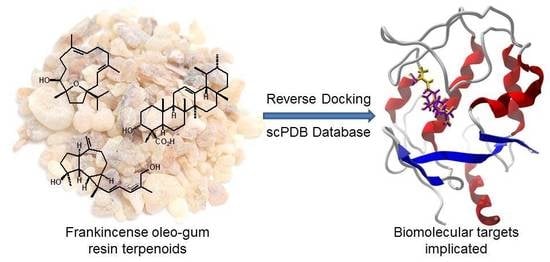
:
1. Introduction
2. Materials and Methods
2.1. Ligand Preparation
2.2. Reverse Molelcular Docking
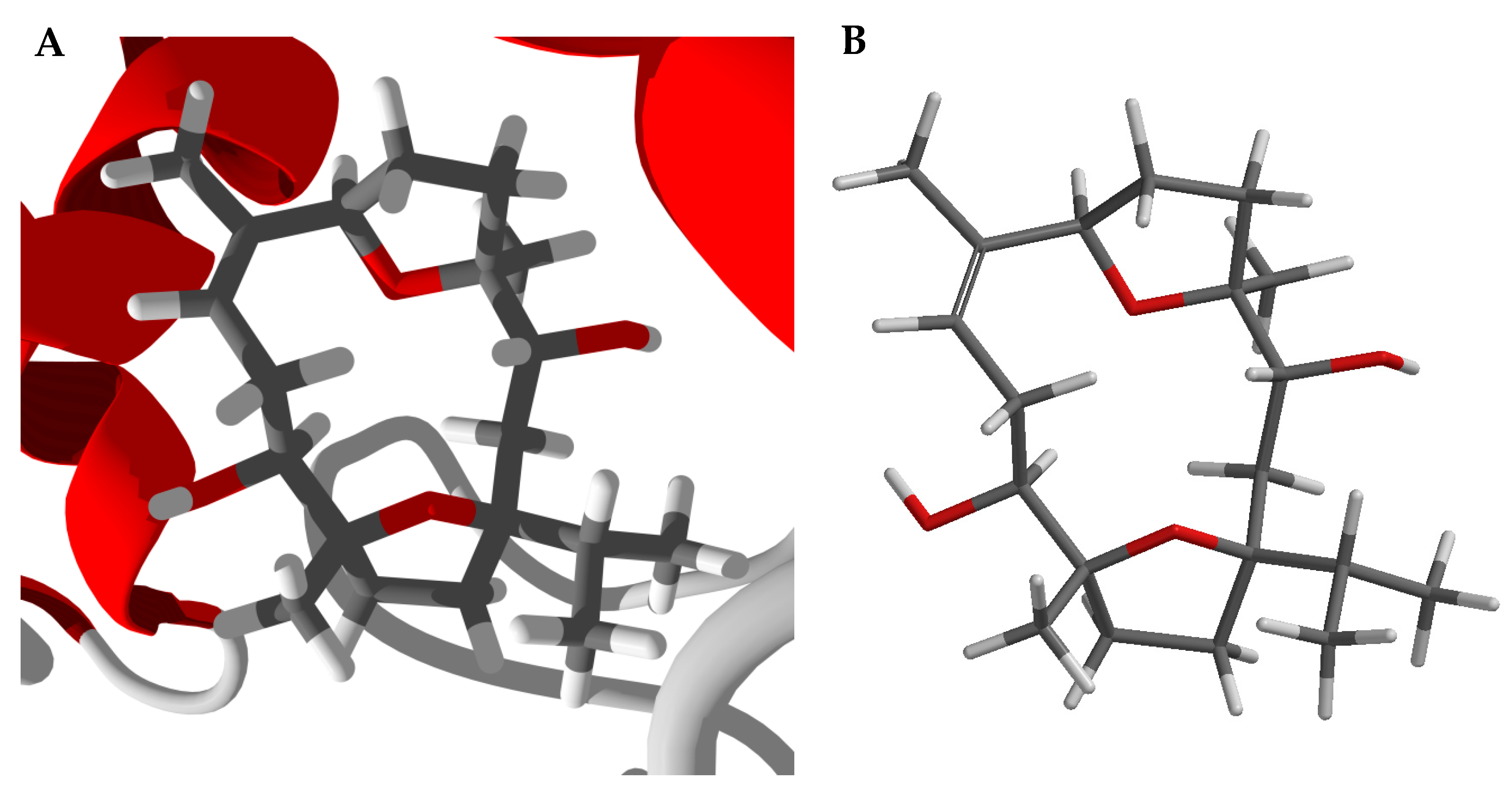
2.3. Conformational Analysis of Boscartol D
3. Results and Discussion
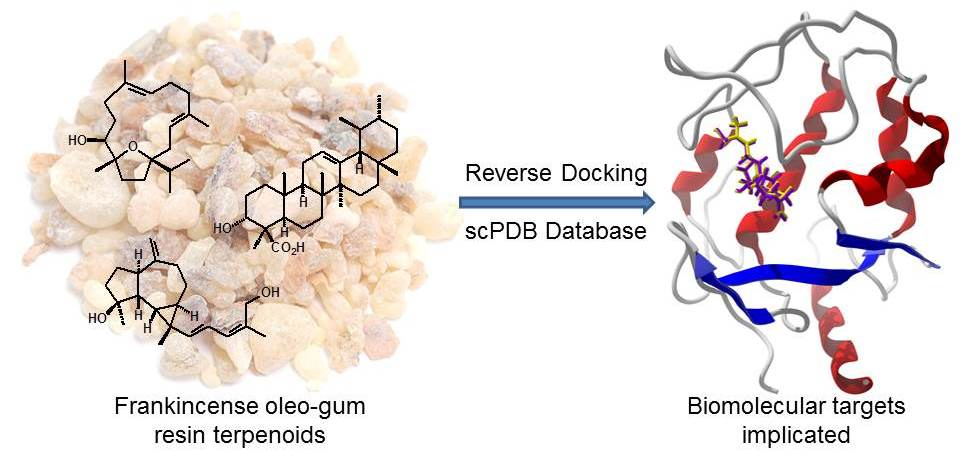
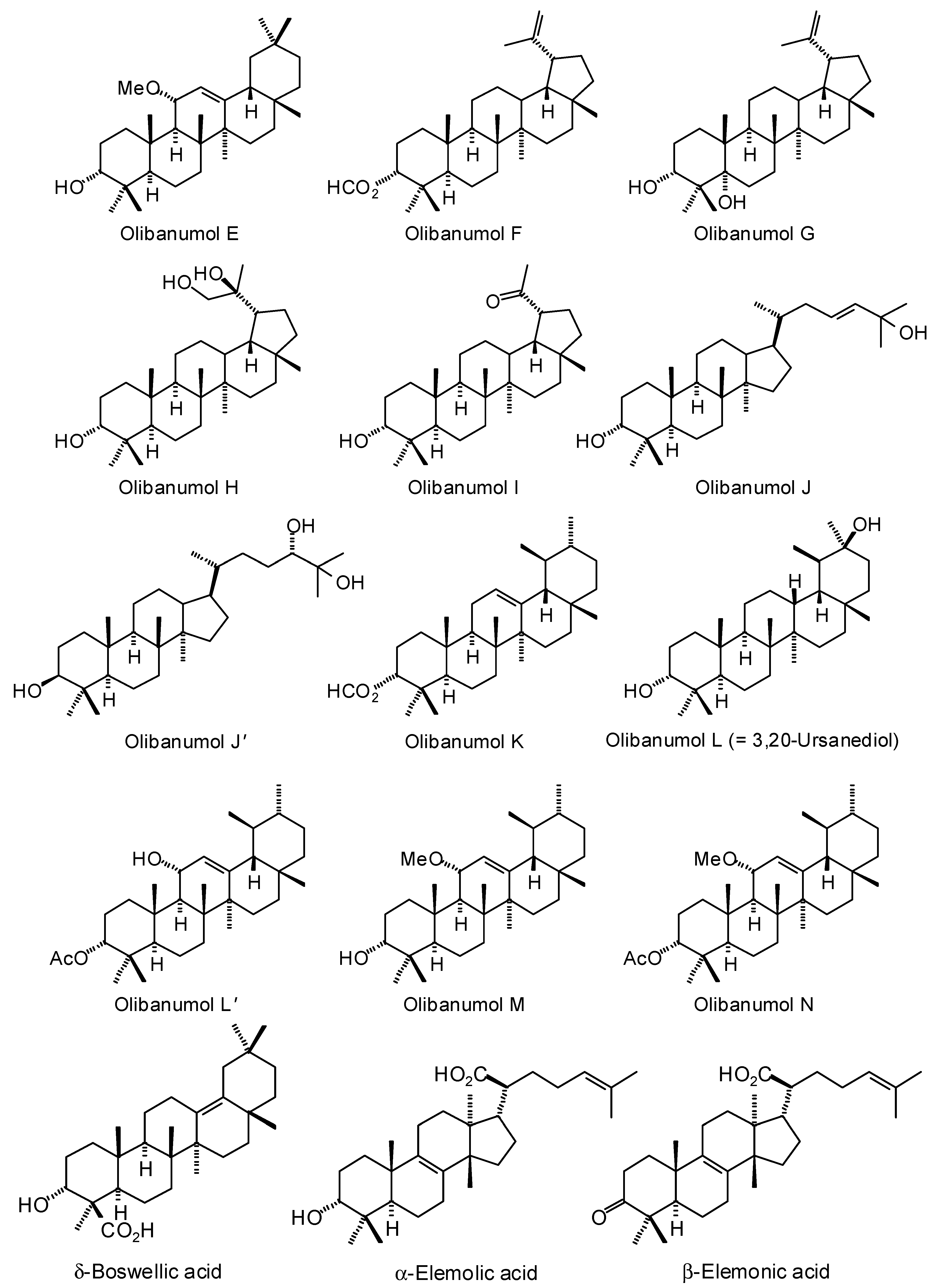
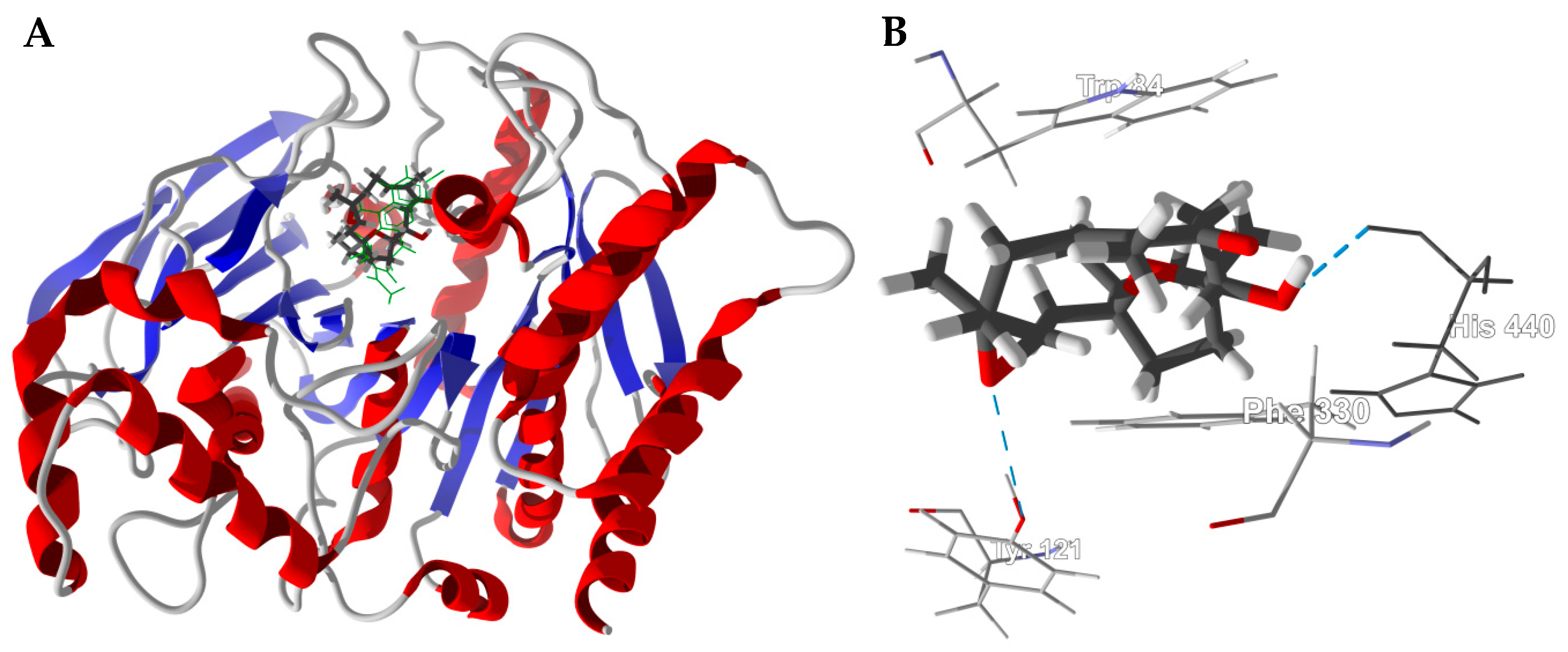
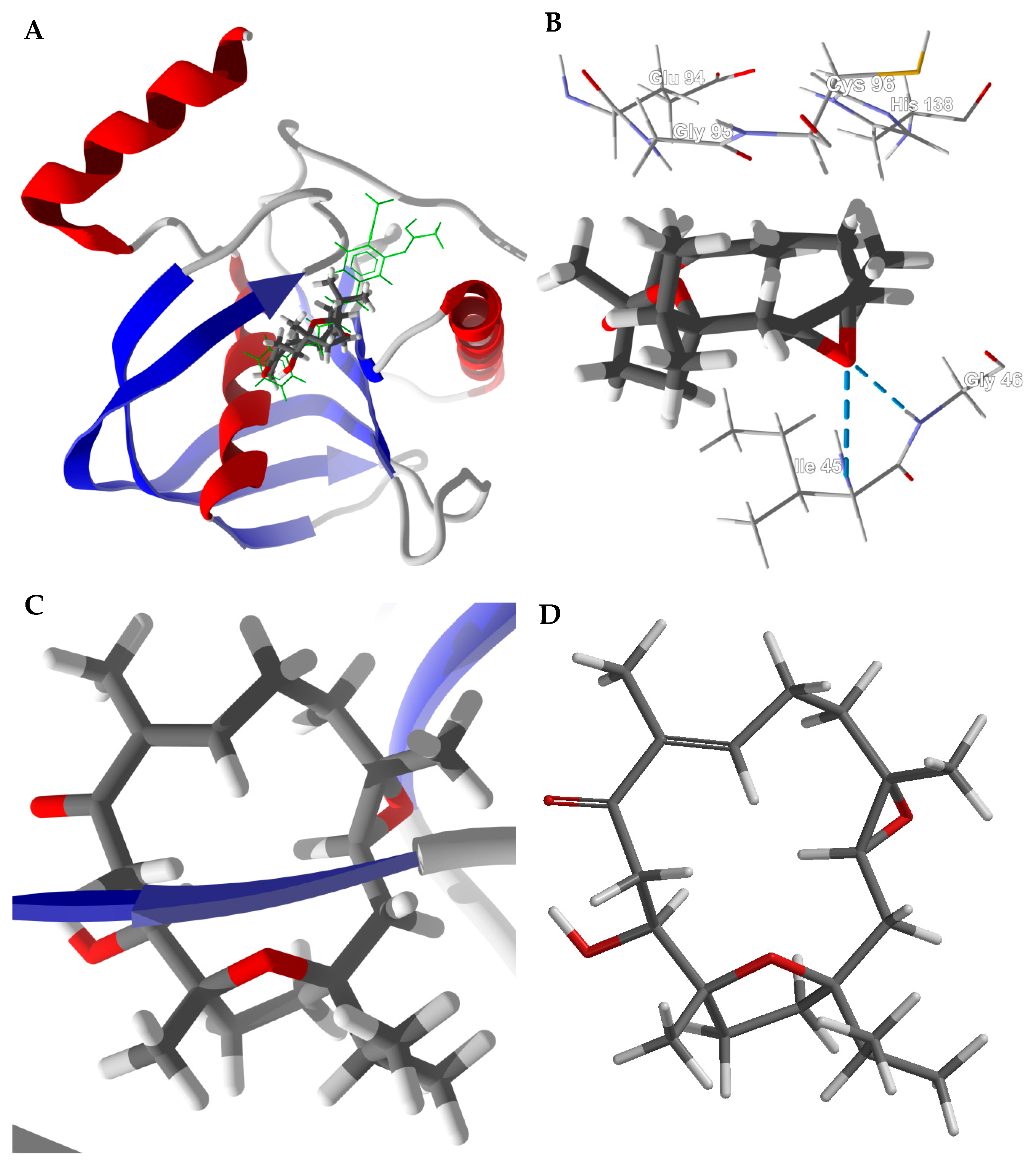
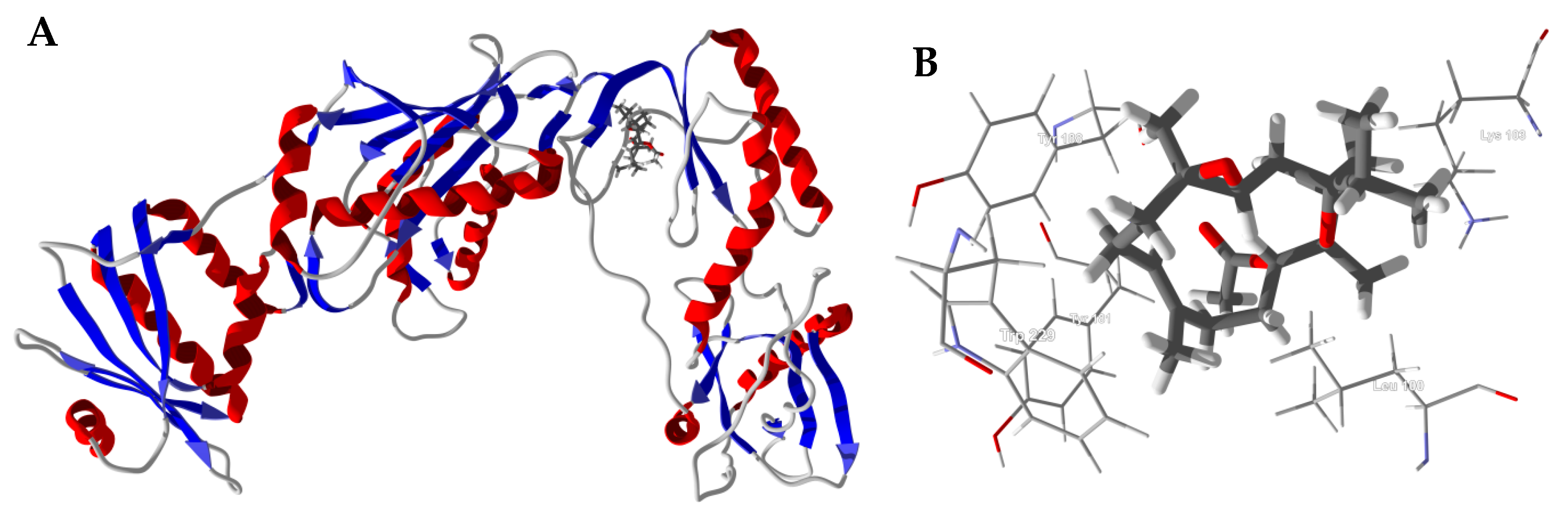
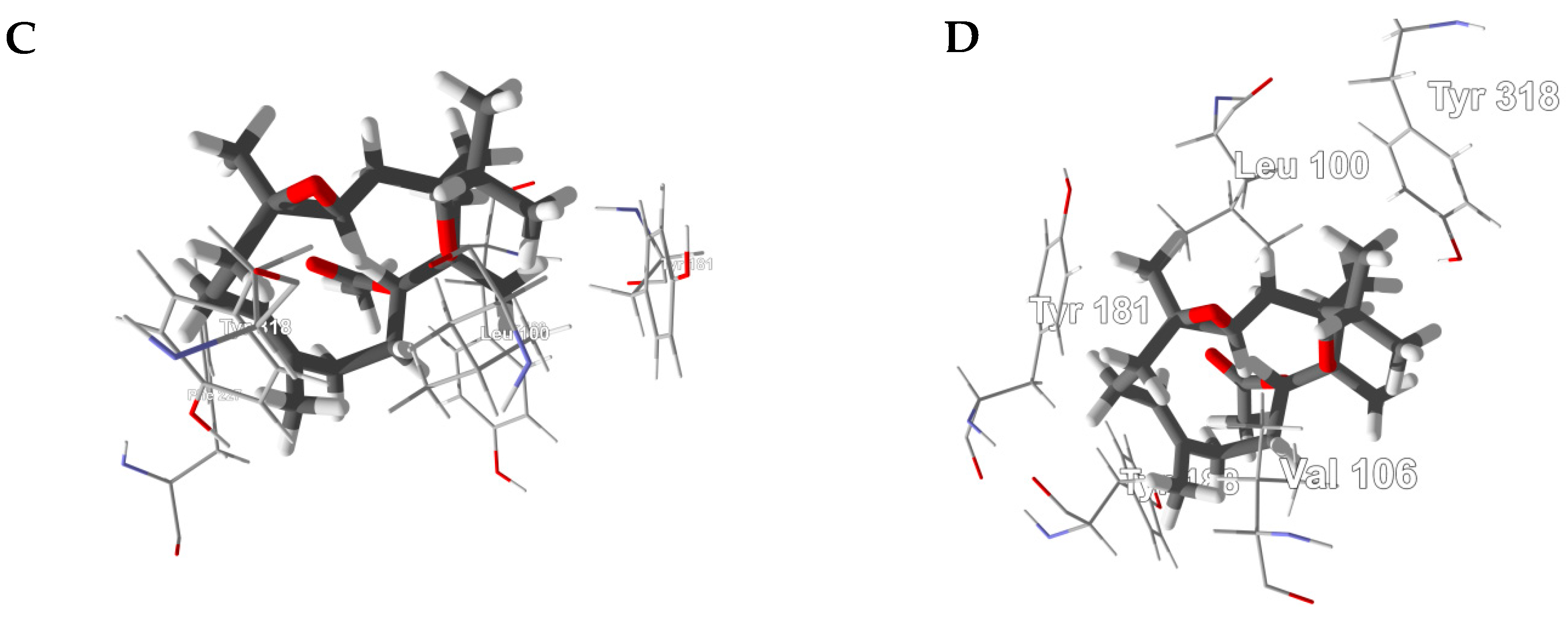
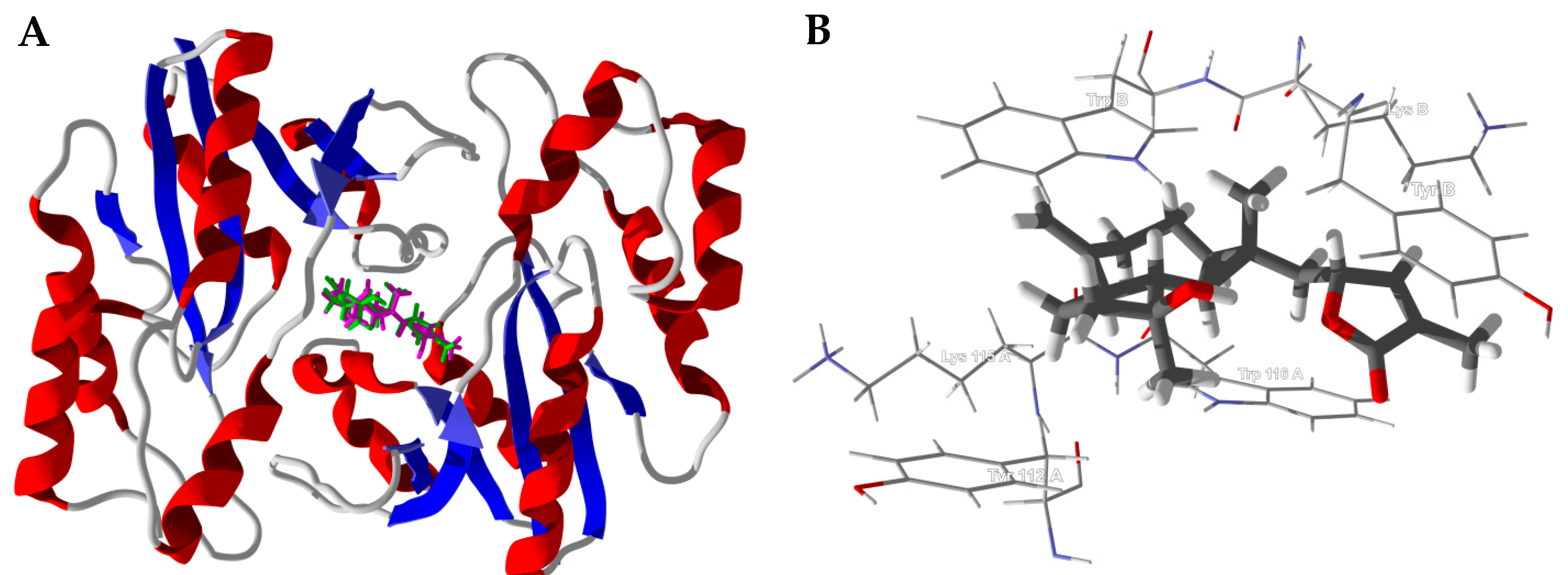
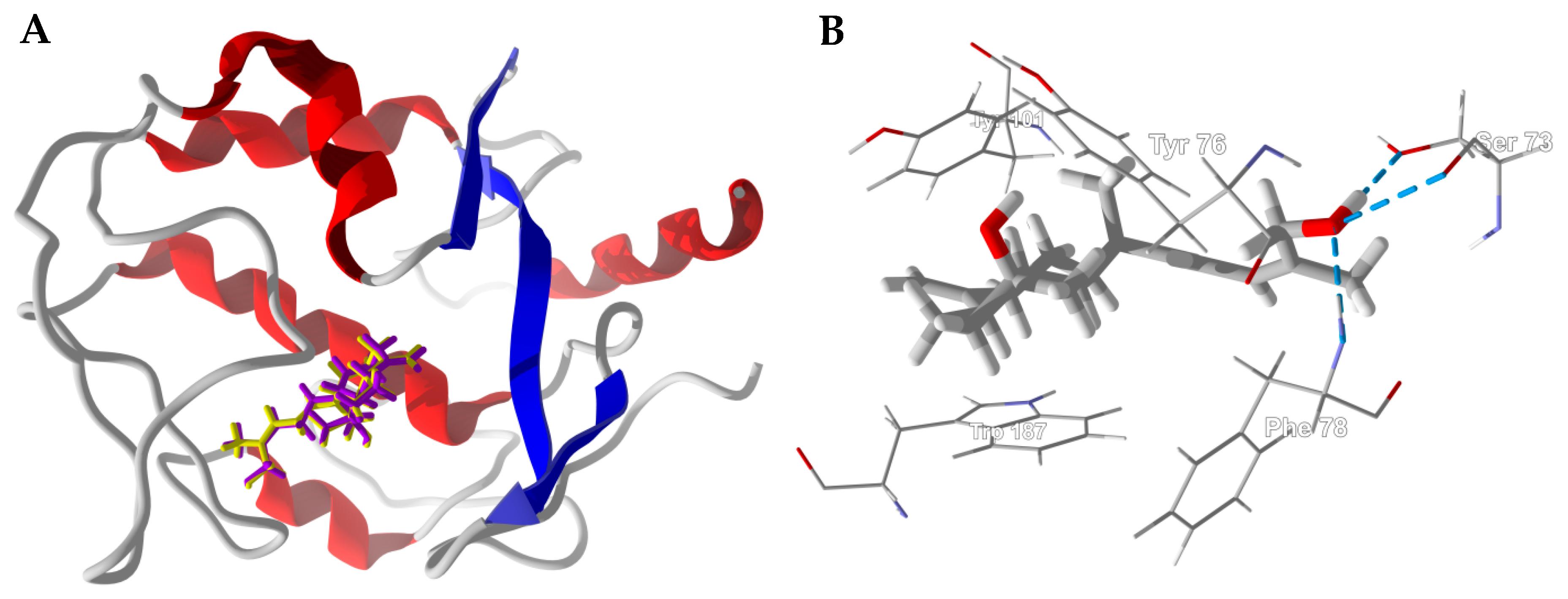
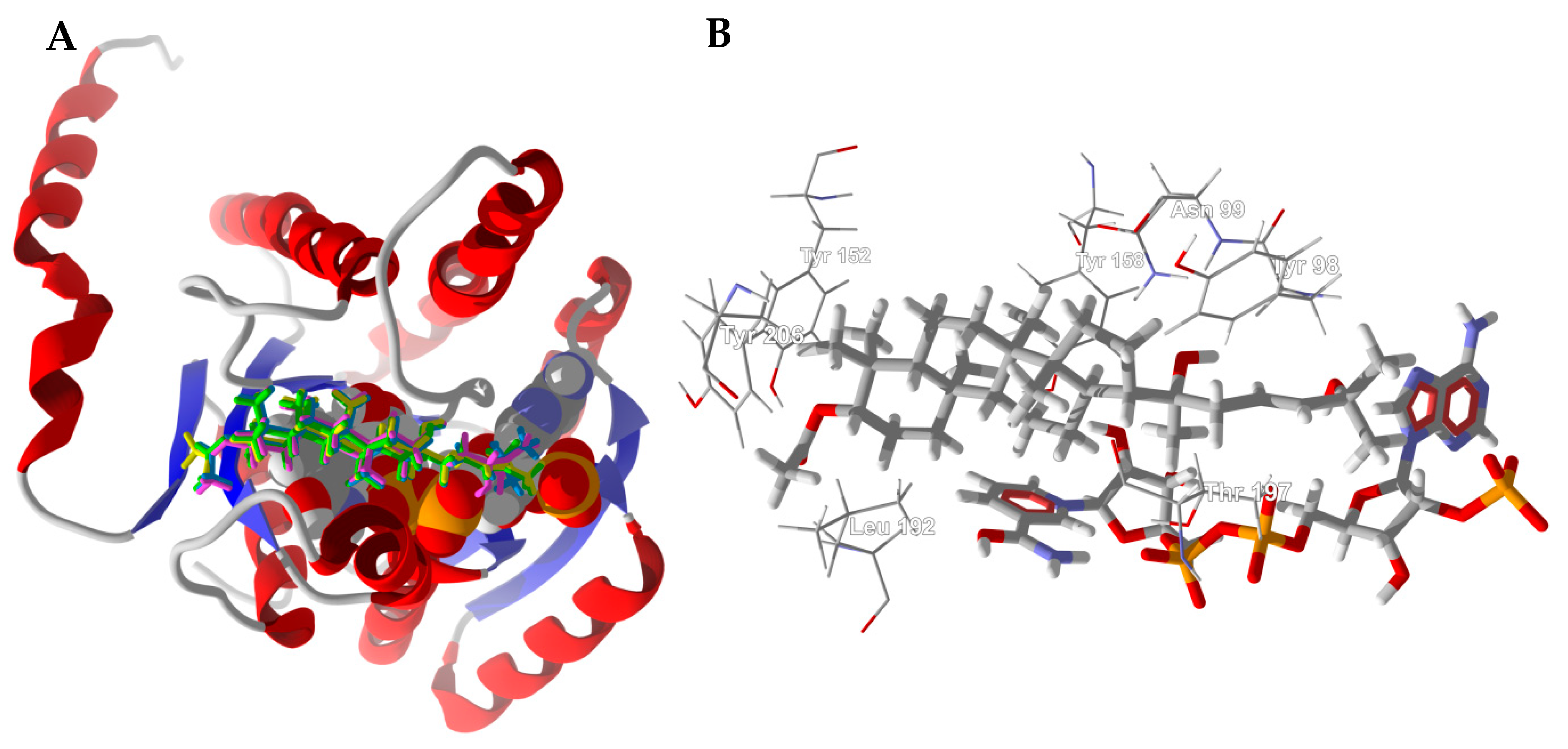
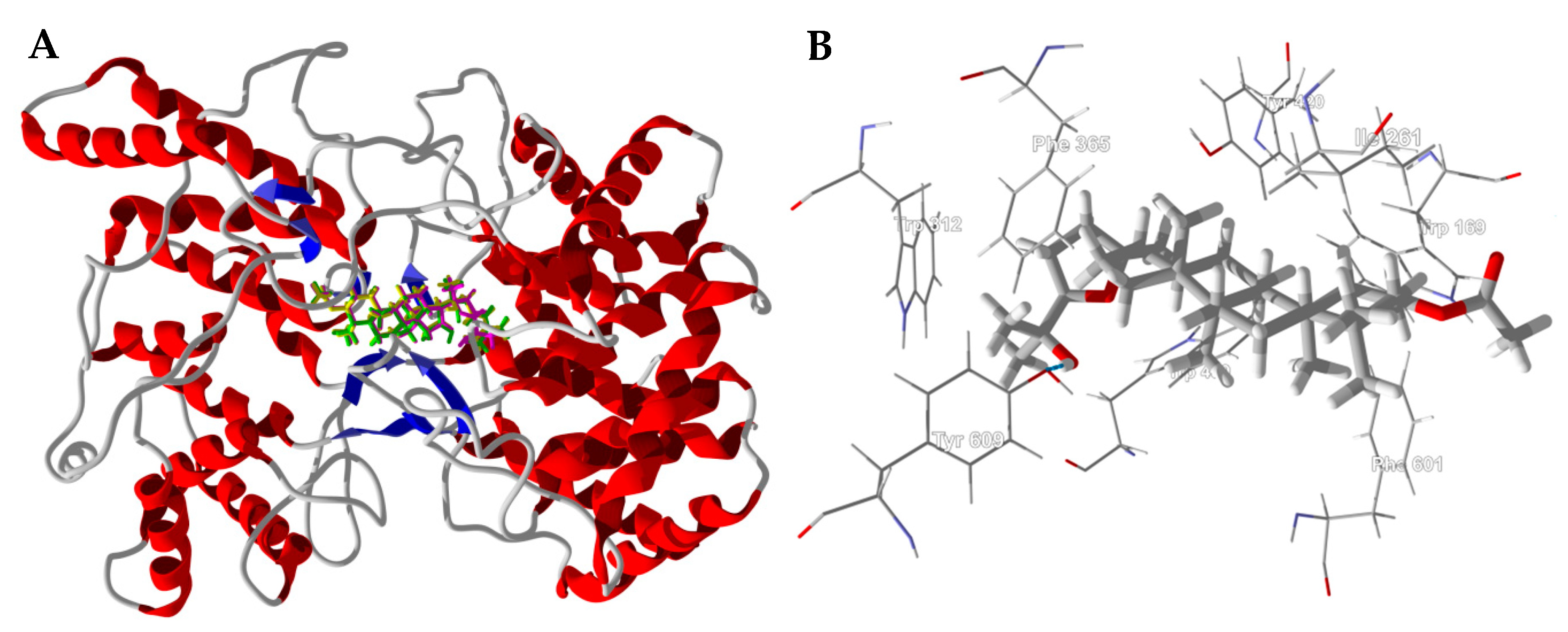
3.1. Cembranoid Diterpenoids
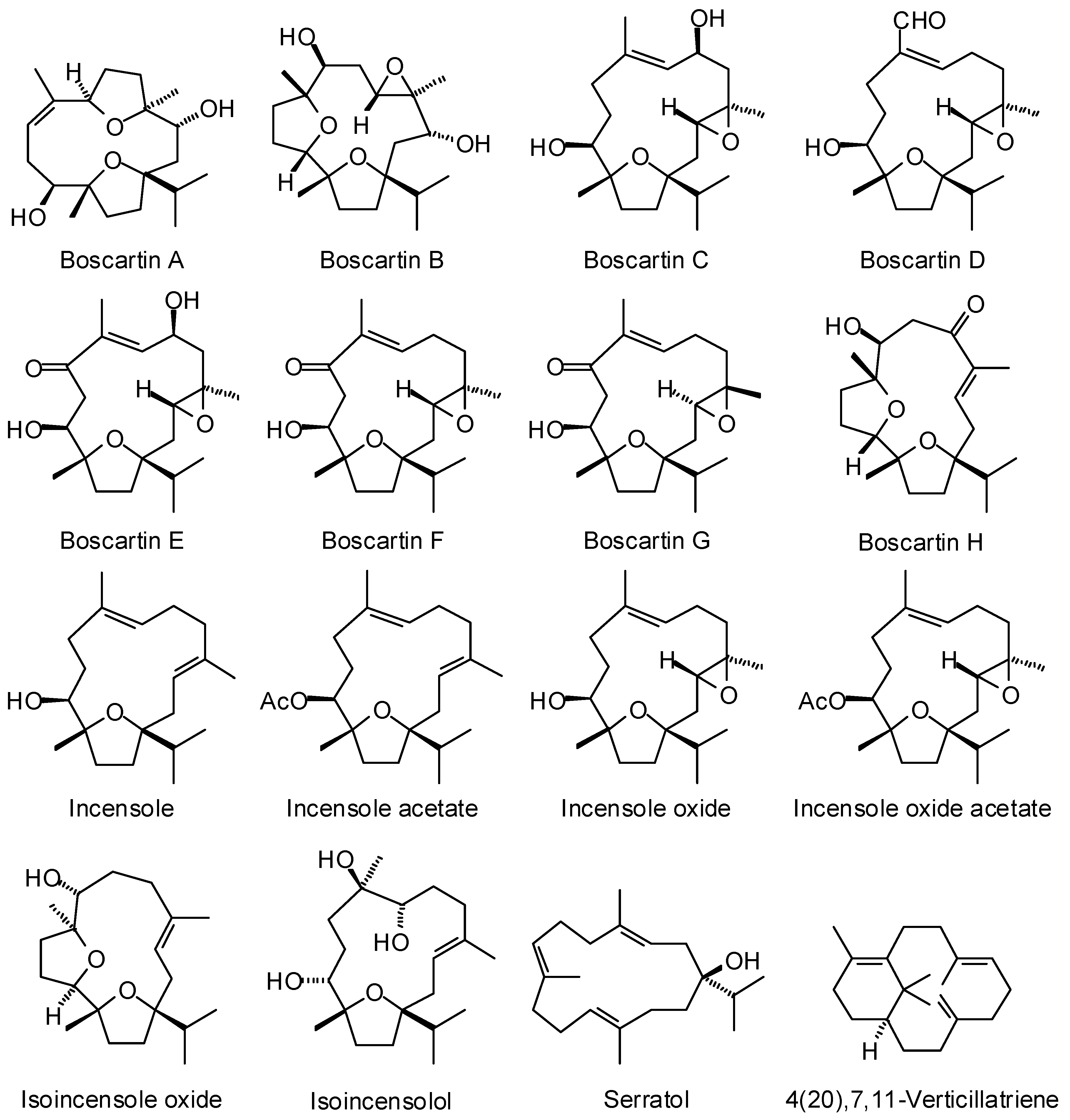
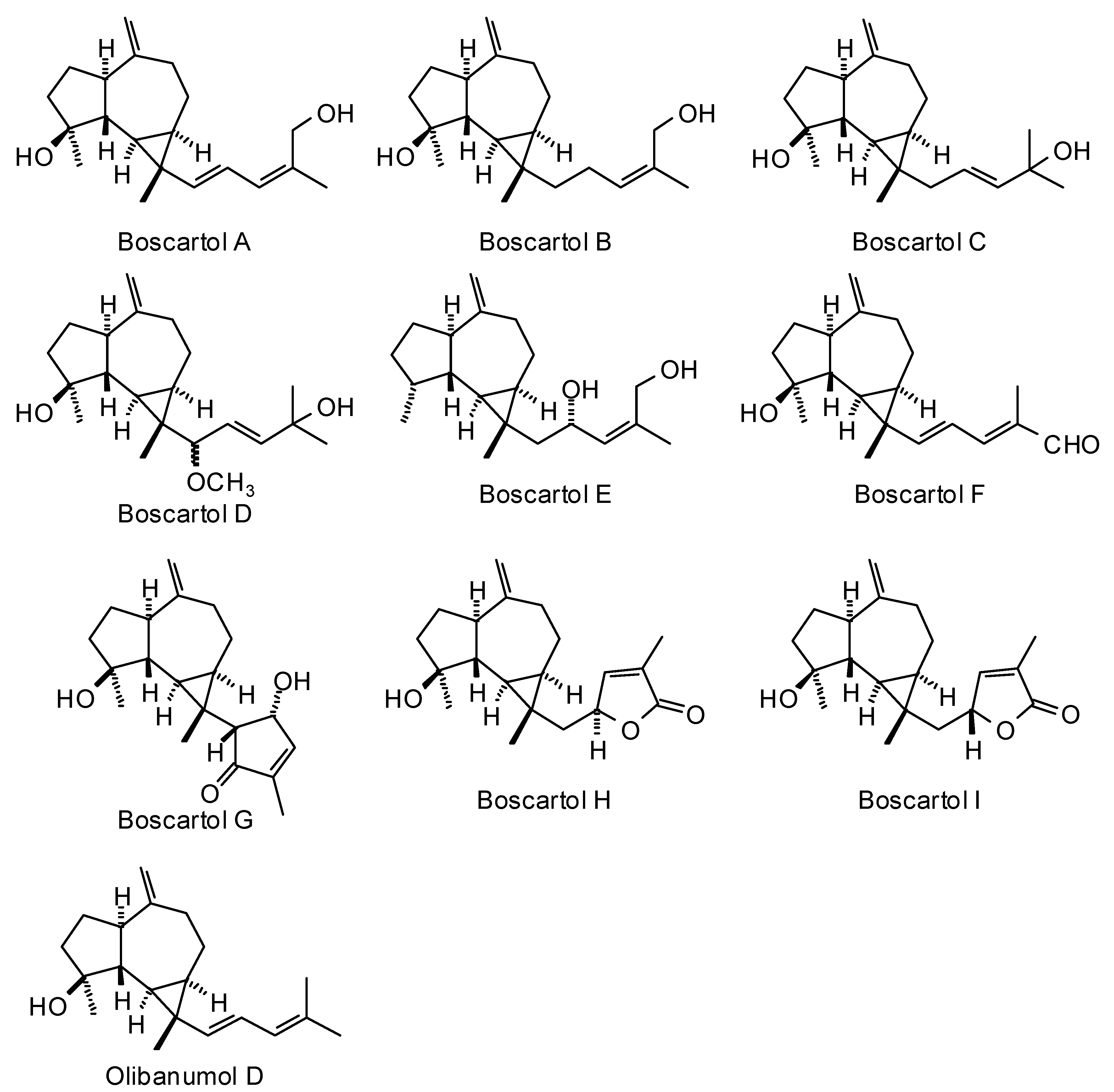
3.2. Cneorubenoid Diterpenoids
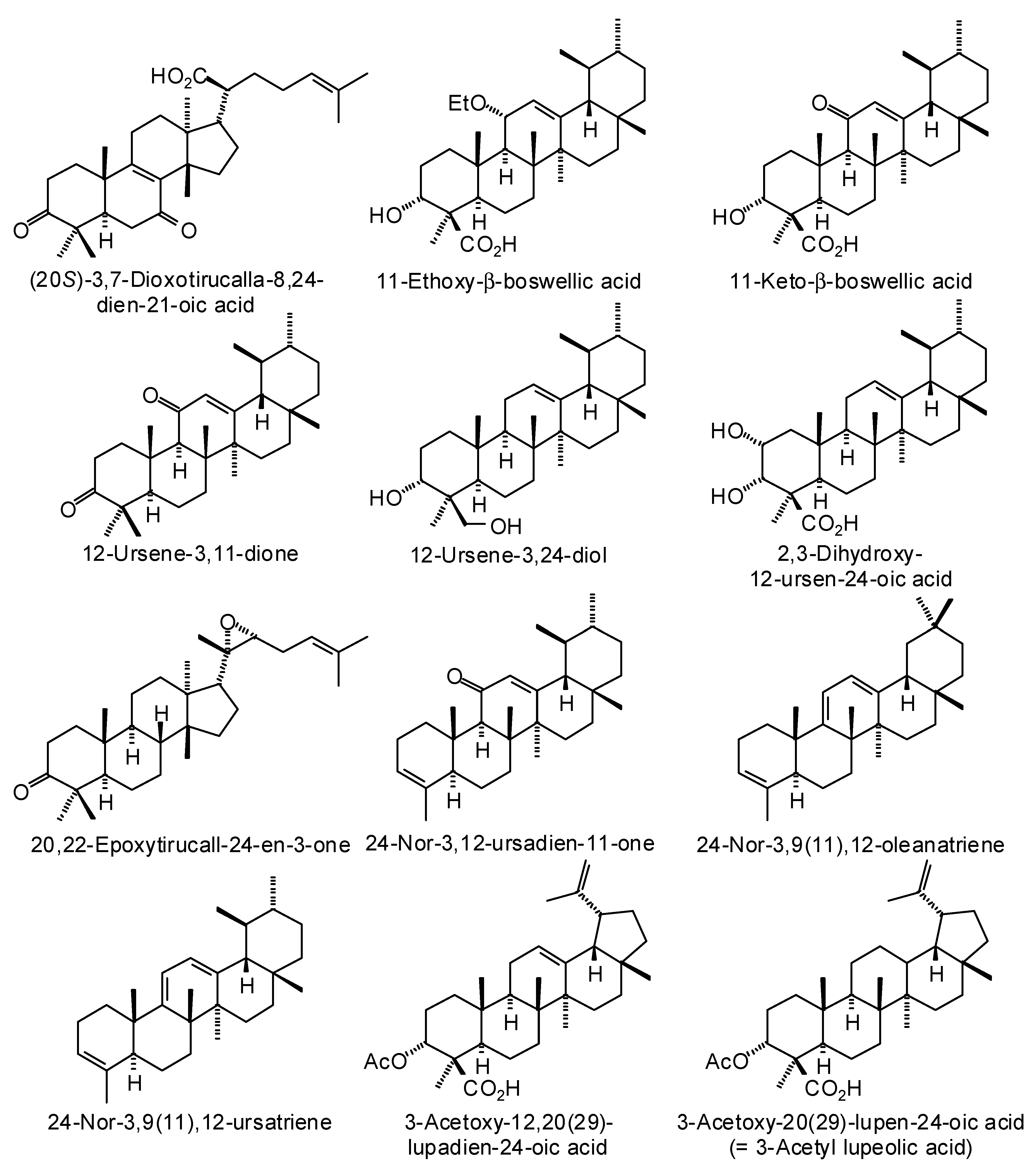
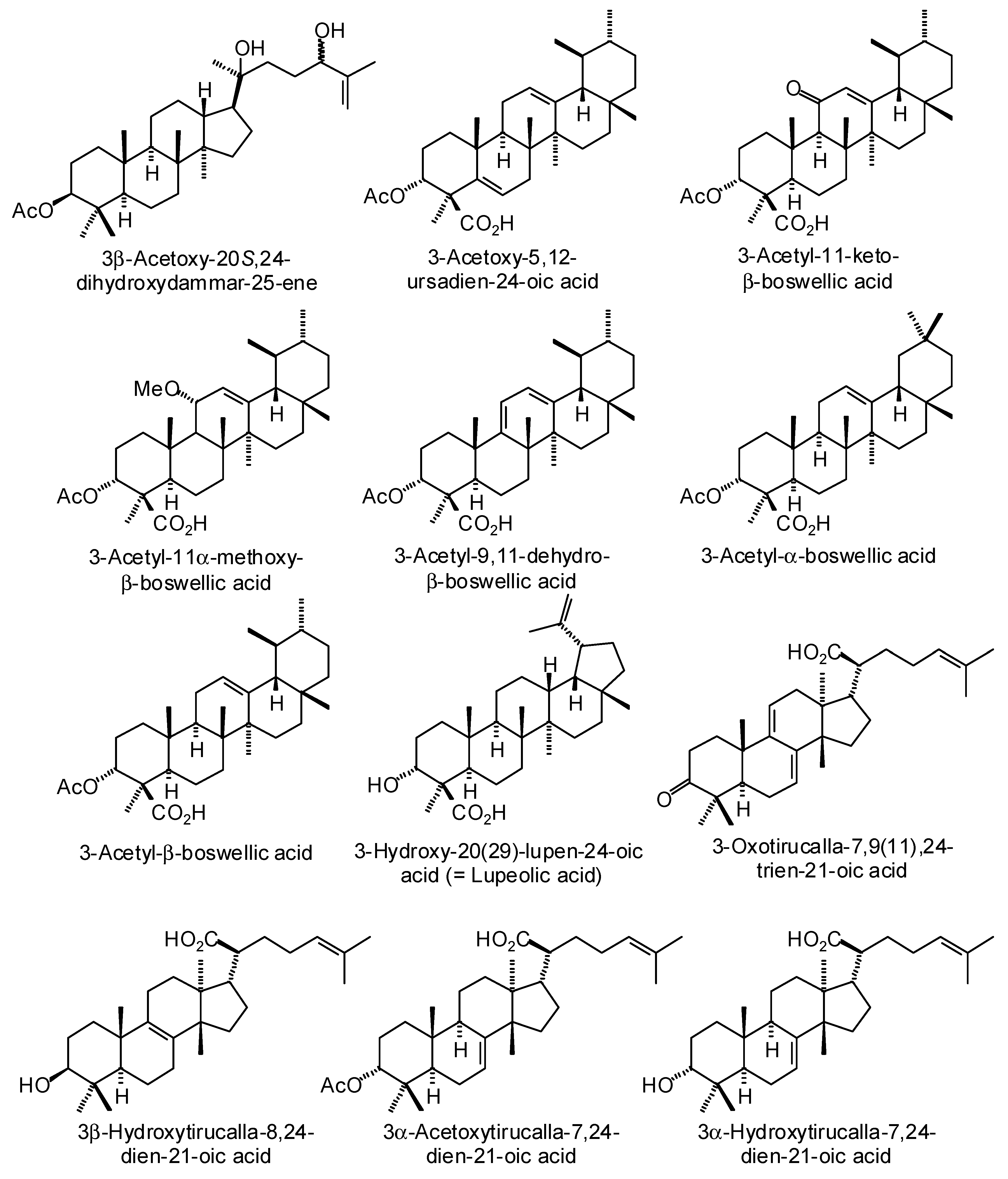
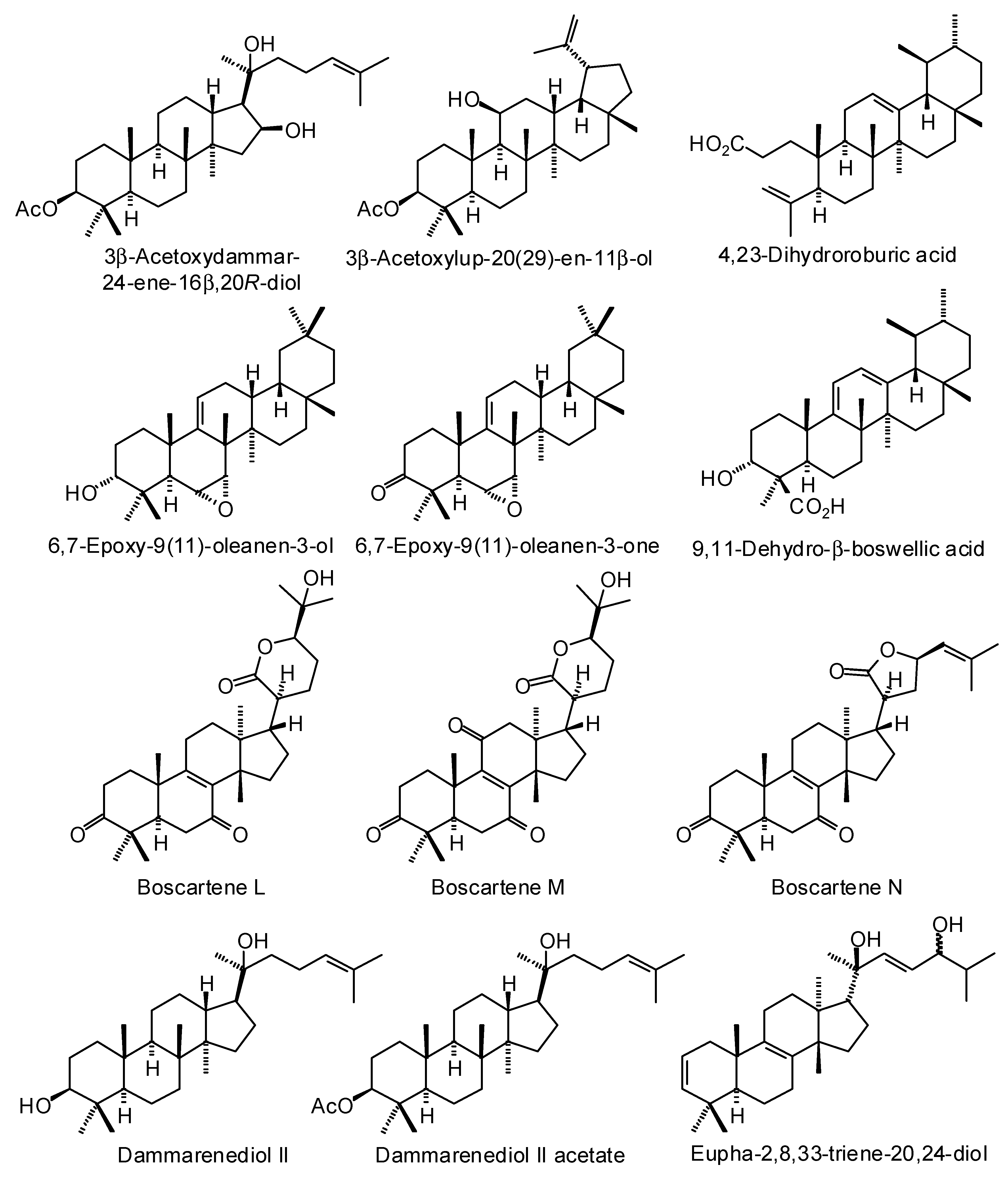
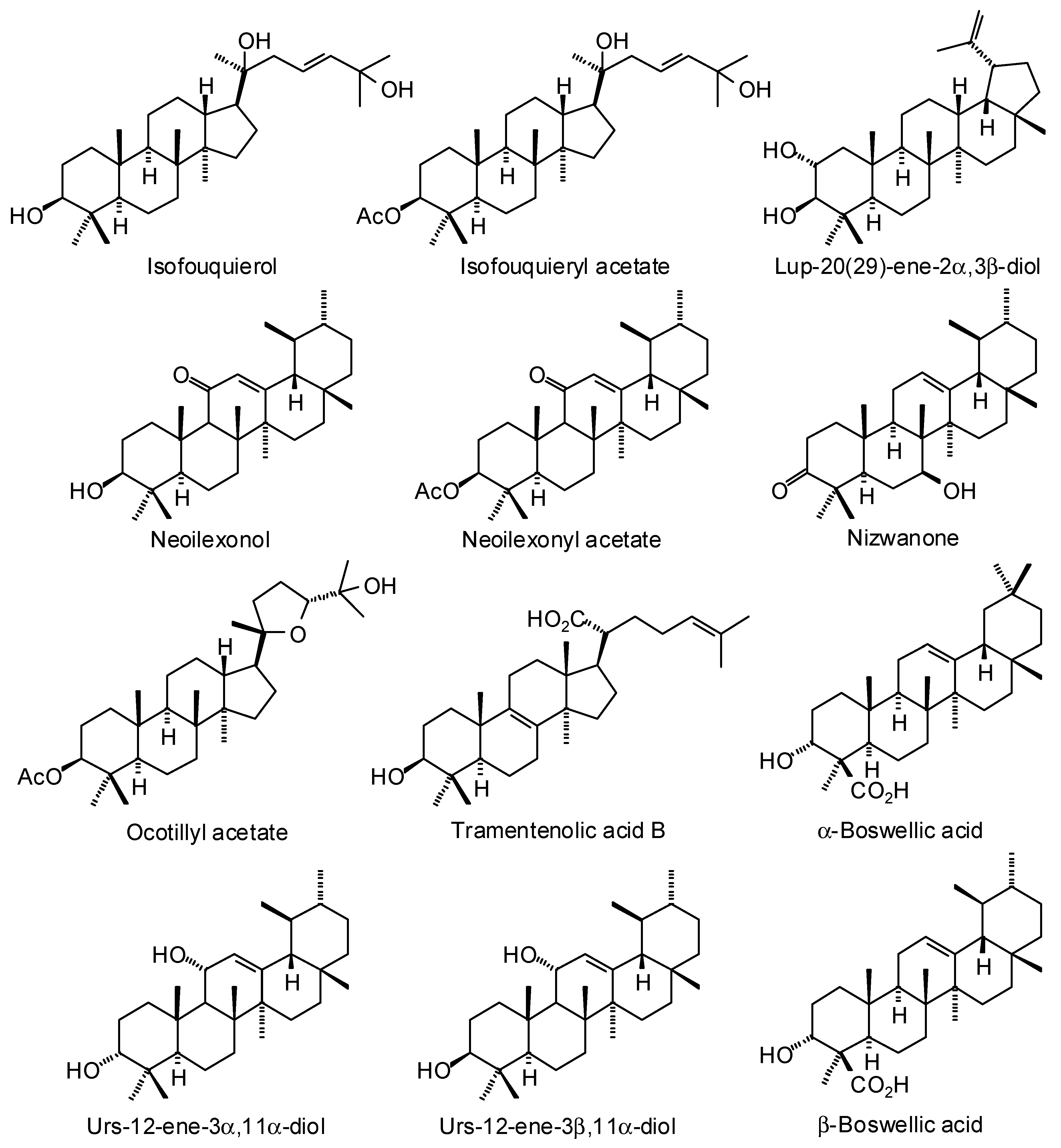
3.3. Boswellia Triterpenoids
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hepper, F.N. Arabian and African frankincense trees. J. Egypt. Archaeol. 1969, 55, 66–72. [Google Scholar] [CrossRef]
- Thulin, M.; Warfa, A.M. The frankincense trees (Boswellia spp., Burseraceae) of northern Somalia and southern Arabia. Kew Bull. 1987, 42, 487–500. [Google Scholar] [CrossRef]
- Langenheim, J.H. Plant Resins: Chemistry, Evolution, Ecology, and Ethnobotany; Timber Press, Inc.: Portland, OR, USA, 2003. [Google Scholar]
- Gebrehiwot, K.; Muys, B.; Haile, M.; Mitloehner, R. Introducing Boswellia papyrifera (Del.) Hochst and its non-timber forest product, frankincense. Int. For. Rev. 2003, 5, 348–353. [Google Scholar] [CrossRef]
- Murthy, T.K.; Shiva, M.P. Salai Guggul from Boswellia serrata Roxb.-its exploitation and utilisation. Indian For. 1977, 103, 466–473. [Google Scholar]
- Moussaieff, A.; Mechoulam, R. Boswellia resin: From religious ceremonies to medical uses; a review of in-vitro, in-vivo and clinical trials. J. Pharm. Pharmacol. 2009, 61, 1281–1293. [Google Scholar] [CrossRef] [PubMed]
- Frawley, D.; Lad, V. The Yoga of Herbs: An Ayurvedic Guide to Herbal Medicine, 2nd ed.; Lotus Press: Twin Lakes, WI, USA, 2001. [Google Scholar]
- Mies, B.A.; Lavranos, J.J.; James, G.J. Frankincense on Soqotra island (Boswellia, Burseraceae; Yemen). Cactus Succul. J. 2000, 72, 265–278. [Google Scholar]
- Getahon, A. Some Common Medicinal and Poisonous Plants Used in Ethiopian Folkmedicine; Addis Abeba University: Addis Abeba, Ethiopia, 1976. [Google Scholar]
- Dannaway, F.R. Strange fires, weird smokes and psychoactive combustibles: Entheogens and incense in ancient traditions. J. Psychoact. Drugs 2010, 42, 485–497. [Google Scholar] [CrossRef] [PubMed]
- Mertens, M.; Buettner, A.; Kirchhoff, E. The volatile constituents of frankincense—A review. Flavour Fragr. J. 2009, 24, 279–300. [Google Scholar] [CrossRef]
- Paul, M.; Brüning, G.; Bergmann, J.; Jauch, J. A thin-layer chromatography method for the identification of three different olibanum resins (Boswellia serrata, Boswellia papyrifera and Boswellia carterii, respectively, Boswellia sacra). Phytochem. Anal. 2012, 23, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Wang, Y.-G.; Wang, A.-G.; Wu, L.-Q.; Zhang, H.-J.; Wang, W.-J.; Su, Y.-L.; Qin, H.-L. Cembranoids from the gum resin of Boswellia carterii as potential antiulcerative colitis agents. J. Nat. Prod. 2015, 78, 2322–2331. [Google Scholar] [CrossRef] [PubMed]
- Boscarelli, A.; Giglio, E.; Quagliata, C. Structure and conformation of incensole oxide. Acta Crystallogr. Sect. B 1981, 37, 744–746. [Google Scholar] [CrossRef] [Green Version]
- Hamm, S.; Bleton, J.; Connan, J.; Tchapla, A. A chemical investigation by headspace SPME and GC–MS of volatile and semi-volatile terpenes in various olibanum samples. Phytochemistry 2005, 66, 1499–1514. [Google Scholar] [CrossRef] [PubMed]
- Forcellese, M.L.; Nicoletti, R.; Petrossi, U. The structure of isoincensole-oxide. Tetrahedron 1972, 28, 325–331. [Google Scholar] [CrossRef]
- Li, F.; Xu, K.; Yuan, S.; Yan, D.; Liu, R.; Tan, J.; Zeng, G.; Zhou, Y.; Tan, G. Macrocyclic diterpenes from Boswellia carterii Birdwood (frankincense). Chin. J. Org. Chem. 2010, 30, 107–111. (In Chinese) [Google Scholar]
- Basar, S.; Koch, A.; König, W.A. A verticillane-type diterpene from Boswellia carterii essential oil. Flavour Fragr. J. 2001, 16, 315–318. [Google Scholar] [CrossRef]
- Schmidt, T.J.; Kaiser, M.; Brun, R. Complete structural assignment of serratol, a cembrane-type diterpene from Boswellia serrata, and evaluation of its antiprotozoal activity. Planta Med. 2011, 77, 849–850. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, T.; Oominami, H.; Matsuda, H.; Yoshikawa, M. New terpenoids, olibanumols D–G, from traditional Egyptian medicine olibanum, the gum-resin of Boswellia carterii. J. Nat. Med. 2011, 65, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ren, J.; Wang, A.; Yang, J.; Ji, T.; Ma, Q.-G.; Tian, J.; Su, Y. Hepatoprotective prenylaromadendrane-type diterpenes from the gum resin of Boswellia carterii. J. Nat. Prod. 2013, 76, 2074–2079. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ning, Z.; Lu, C.; Zhao, S.; Wang, J.; Liu, B.; Xu, X.; Liu, Y. Triterpenoid resinous metabolites from the genus Boswellia: Pharmacological activities and potential species-identifying properties. Chem. Cent. J. 2013, 7, 153. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Khajuria, A.; Taneja, S.C.; Johri, R.K.; Singh, J.; Qazi, G.N. Boswellic acids: A leukotriene inhibitor also effective through topical application in inflammatory disorders. Phytomedicine 2008, 15, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Banno, N.; Akihisa, T.; Yasukawa, K.; Tokuda, H.; Tabata, K.; Nakamura, Y.; Nishimura, R.; Kimura, Y.; Suzuki, T. Anti-inflammatory activities of the triterpene acids from the resin of Boswellia carteri. J. Ethnopharmacol. 2006, 107, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, M.; Morikawa, T.; Oominami, H.; Matsuda, H. Absolute stereostructures of olibanumols A, B, C, H, I, and J from olibanum, gum-resin of Boswellia carterii, and inhibitors of nitric xxide production in lipopolysaccharide-activated mouse peritoneal macrophages. Chem. Pharm. Bull. 2009, 57, 957–964. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, T.; Oominami, H.; Matsuda, H.; Yoshikawa, M. Four new ursane-type triterpenes, olibanumols K, L, M, and N, from traditional Egyptian medicine olibanum, the gum-resin of Boswellia carterii. Chem. Pharm. Bull. 2010, 58, 1541–1544. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Ren, J.; Wang, A. Isolation, characterization, and hepatoprotective activities of terpenes from the gum resin of Boswellia carterii Birdw. Phytochem. Lett. 2018, 23, 73–77. [Google Scholar] [CrossRef]
- Halgren, T.A. Merck Molecular Force Field. I. Basis, form, scope, parameterization, and performance of MMFF94. J. Comput. Chem. 1996, 17, 490–519. [Google Scholar] [CrossRef]
- Zhao, Y.; Truhlar, D.G. The M06 suite of density functionals for main group thermochemistry, thermochemical kinetics, noncovalent interactions, excited states, and transition elements: Two new functionals and systematic testing of four M06-class functionals and 12 other function. Theor. Chem. Acc. 2008, 120, 215–241. [Google Scholar] [CrossRef]
- Marenich, A.V.; Olson, R.M.; Kelly, C.P.; Cramer, C.J.; Truhlar, D.G. Self-consistent reaction field model for aqueous and nonaqueous solutions based on accurate polarized partial charges. J. Chem. Theory Comput. 2007, 3, 2011–2033. [Google Scholar] [CrossRef] [PubMed]
- Setzer, W.N. Conformational analysis of macrocyclic frankincense (Boswellia) diterpenoids. J. Mol. Model. 2018, 24, 74. [Google Scholar] [CrossRef] [PubMed]
- Desaphy, J.; Bret, G.; Rognan, D.; Kellenberger, E. sc-PDB: A 3D-database of ligandable binding sites. Available online: http://bioinfo-pharma.u-strasbg.fr/scPDB/ (accessed on 11 September 2017).
- Thomsen, R.; Christensen, M.H. MolDock: A new technique for high-accuracy molecular docking. J. Med. Chem. 2006, 49, 3315–3321. [Google Scholar] [CrossRef] [PubMed]
- Setzer, M.S.; Sharifi-Rad, J.; Setzer, W.N. The search for herbal antibiotics: An in-silico investigation of antibacterial phytochemicals. Antibiotics 2016, 5, 30. [Google Scholar] [CrossRef] [PubMed]
- Kryger, G.; Silman, I.; Sussman, J.L. Structure of acetylcholinesterase complexed with E2020 (Aricept®): Implications for the design of new anti-Alzheimer drugs. Structure 1999, 7, 297–307. [Google Scholar] [CrossRef]
- Dabaghian, F.; Azadi, A.; Setooni, M.; Zarshenas, M.M. An overview on multi-ingredient memory enhancers and anti-Alzheimer’s formulations from traditional Persian pharmacy. Trends Pharm. Sci. 2017, 3, 215–220. [Google Scholar]
- Hosseinkhani, A.; Sahragard, A.; Namdari, A.; Zarshenas, M.M. Botanical sources for Alzheimer’s: A review on reports from traditional Persian medicine. Am. J. Alzheimer’s Dis. Other Dement. 2017, 32, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Zaker, S.R.; Beheshti, S.; Aghaie, R.; Noorbakhshnia, M. Effect of olibanum on a rat model of Alzheimer’s disease induced by intracerebroventricular injection of streptozotocin. Physiol. Pharmacol. 2015, 18, 477–489. [Google Scholar]
- Beheshti, S.; Aghaie, R. Therapeutic effect of frankincense in a rat model of Alzheimer’s disease. Avicenna J. Phytomed. 2016, 6, 468–475. [Google Scholar] [PubMed]
- Mahboubi, M.; Taghizadeh, M.; Talaei, S.A.; Firozeh, S.M.T.; Rashidi, A.A.; Tamtaji, O.R. Combined administration of Melissa officinalis and Boswellia serrata extracts in an animal model of memory. Iran. J. Psychiatry Behav. Sci. 2016, 10, e681. [Google Scholar] [CrossRef] [PubMed]
- Tajadini, H.; Saifadini, R.; Choopani, R.; Mehrabani, M.; Kamalinejad, M.; Haghdoost, A.A. Herbal medicine Davaie Loban in mild to moderate Alzheimer’s disease: A 12-week randomized double-blind placebo-controlled clinical trial. Complement. Ther. Med. 2015, 23, 767–772. [Google Scholar] [CrossRef] [PubMed]
- Aghajani, M.; Taghizadeh, M.; Maghaminejad, F.; Rahmani, M. Effect of frankincense extract and lemon balm extract co-supplementation on memory of the elderly. Complement. Med. J. Fac. Nurs. Midwifery 2017, 7, 1968–1977. [Google Scholar]
- Ismail, S.M.; Aluru, S.; Sambasivarao, K.R.S.; Matcha, B. Antimicrobial activity of frankincense of Boswellia serrata. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 1095–1101. [Google Scholar]
- Patel, N.B.; Patel, K.C. Antibacterial activity of Boswellia serrata Roxb. ex Colebr. ethanomedicinal plant against Gram negative UTI pathogens. Life Sci. Leafl. 2014, 53, 79–88. [Google Scholar]
- El Kichaoui, A.; Abdelmoneim, A.; Elbaba, H.; El Hindi, M. The antimicrobial effects of Boswellia carterii, Glycyrrhiza glabra and Rosmarinus officinalis some pathogenic microorganisms. IUG J. Nat. Stud. 2017, 25, 208–213. [Google Scholar]
- Michie, C.A.; Cooper, E. Frankincense and myrrh as remedies in children. J. R. Soc. Med. 1991, 84, 602–605. [Google Scholar] [PubMed]
- Abdallah, E.M.; Khalid, A.S.; Ibrahim, N. Antibacterial activity of oleo-gum resins of Commiphora molmol and Boswellia papyrifera against methicillin resistant Staphylococcus aureus (MRSA). Sci. Res. Essays 2009, 4, 351–356. [Google Scholar]
- Khosravi Samani, M.; Mahmoodian, H.; Moghadamnia, A.A.; Poorsattar Bejeh Mir, A.; Chitsazan, M. The effect of frankincense in the treatment of moderate plaque-induced gingivitis: A double blinded randomized clinical trial. DARU J. Pharm. Sci. 2011, 19, 288–294. [Google Scholar]
- Tee, W.; Lambert, J.R.; Dwyer, B. Cytotoxin production by Helicobacter pylori from patients with upper gastrointestinal-tract diseases. J. Clin. Microbiol. 1995, 33, 1203–1205. [Google Scholar] [PubMed]
- Lemenih, M.; Teketay, D. Frankincense and myrrh resources of Ethiopia: II. Medicinal and industrial uses. Ethiop. J. Sci. 2003, 26, 161–172. [Google Scholar]
- El-Mekkawy, S.; Meselhy, M.R.; Kusumoto, I.T.; Kadota, S.; Hattori, M.; Namba, T. Inhibitory effects of Egyptian folk medicines on human immunodeficiency virus (HIV) reverse transcriptase. Chem. Pharm. Bull. 1995, 43, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Karplus, M. Vicinal proton coupling in nuclear magnetic resonance. J. Am. Chem. Soc. 1963, 85, 2870–2871. [Google Scholar] [CrossRef]
- Haasnoot, C.A.G.; de Leeuw, F.A.A. M.; Altona, C. The relationship between proton-proton NMR coupling constants and substituent electronegativities—I: An empirical generalization of the Karplus equation. Tetrahedron 1980, 36, 2783–2792. [Google Scholar] [CrossRef]
- Shen, J.; Hilgenbrink, A.R.; Xia, W.; Feng, Y.; Dimitrov, D.S.; Lockwood, M.B.; Amato, R.J.; Low, P.S. Folate receptor-β constitutes a marker for human proinflammatory monocytes. J. Leukoc. Biol. 2014, 96, 563–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, J.; Putt, K.S.; Visscher, D.W.; Murphy, L.; Cohen, C.; Singhal, S.; Sandusky, G.; Feng, Y.; Dimitrov, D.S.; Low, P.S. Assessment of folate receptor-β expression in human neoplastic tissues. Oncotarget 2015, 6, 14700–14709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wibowo, A.S.; Singh, M.; Reeder, K.M.; Carter, J.J.; Kovach, A.R.; Meng, W.; Ratnam, M.; Zhang, F.; Dann, C.E. Structures of human folate receptors reveal biological trafficking states and diversity in folate and antifolate recognition. Proc. Natl. Acad. Sci. USA 2013, 110, 15180–15188. [Google Scholar] [CrossRef] [PubMed]
- Ernst, E. Frankincense: Systematic review. BMJ 2008, 337, a2813. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Ali, R.; Parveen, R.; Najmi, A.K.; Ahmad, S. Pharmacological evidences for cytotoxic and antitumor properties of boswellic acids from Boswellia serrata. J. Ethnopharmacol. 2016, 191, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Iram, F.; Khan, S.A.; Husain, A. Phytochemistry and potential therapeutic actions of Boswellic acids: A mini-review. Asian Pac. J. Trop. Biomed. 2017, 7, 513–523. [Google Scholar] [CrossRef]
- Al-Yasiry, A.R.M.; Kiczorowska, B. Frankincense—Therapeutic properties. Postepy Hig. Med. Dosw. 2016, 70, 380–391. [Google Scholar] [CrossRef]
- Popa, O.M.; Cherciu, M.; Cherciu, L.I.; Dutescu, M.I.; Bojinca, M.; Bojinca, V.; Bara, C.; Popa, L.O. ERAP1 and ERAP2 gene variations influence the risk of psoriatic arthritis in Romanian population. Arch. Immunol. Ther. Exp. 2016, 64, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Syrovets, T.; Kess, D.; Büchele, B.; Hainzl, H.; Lunov, O.; Weiss, J.M.; Scharffetter-Kochanek, K.; Simmet, T. Targeting NF-κB with a natural triterpenoid alleviates skin inflammation in a mouse model of psoriasis. J. Immunol. 2009, 183, 4755–4763. [Google Scholar] [CrossRef] [PubMed]
- Moustafa, F.; Feldman, S.R. A review of phosphodiesterase-inhibition and the potential role for phosphodiesterase 4-inhibitors in clinical dermatology. Dermatol. Online J. 2014, 20, 22608. [Google Scholar] [PubMed]
- Kimmatkar, N.; Thawani, V.; Hingorani, L.; Khiyani, R. Efficacy and tolerability of Boswellia serrata extract in treatment of osteoarthritis of knee—A randomized double blind placebo controlled trial. Phytomedicine 2003, 10, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Cavaillon, J.M. Pro- versus anti-inflammatory cytokines: Myth or reality. Cell. Mol. Biol. 2001, 47, 695–702. [Google Scholar] [PubMed]
- Bachurin, S.O.; Bovina, E.V.; Ustyugov, A.A. Drugs in clinical trials for Alzheimer’s disease: The major trends. Med. Res. Rev. 2017, 37, 1186–1225. [Google Scholar] [CrossRef] [PubMed]
- Brady, N.; Poljak, A.; Jayasena, T.; Sachdev, P. Natural plant-derived acetylcholinesterase inhibitors: Relevance for Alzheimer’s disease. In Natural Products Targeting Clinically Relelvant Enzymes; Andrade, P.B., Valentão, P., Pereira, D.M., Eds.; Wiley-VCH: Weinheim, Germany, 2017; pp. 297–318. [Google Scholar]
- Bürkle, A.; Brabeck, C.; Diefenbach, J.; Beneke, S. The emerging role of poly(ADP-ribose) polymerase-1 in longevity. Int. J. Biochem. Cell Biol. 2005, 37, 1043–1053. [Google Scholar] [CrossRef] [PubMed]
- Bryant, H.E.; Schultz, N.; Thomas, H.D.; Parker, K.M.; Flower, D.; Lopez, E.; Kyle, S.; Meuth, M.; Curtin, N.J.; Helleday, T. Specific killing of BRCA2-deficient tumours with inhibitors of poly(ADP-ribose) polymerase. Nature 2007, 434, 913–917. [Google Scholar] [CrossRef] [PubMed]
- Ratnam, K.; Low, J.A. Current development of clinical inhibitors of poly(ADP-ribose) polymerase in oncology. Clin. Cancer Res. 2007, 13, 1383–1388. [Google Scholar] [CrossRef] [PubMed]
- Xia, W.; Low, P.S. Folate-targeted therapies for cancer. J. Med. Chem. 2010, 53, 6811–6824. [Google Scholar] [CrossRef] [PubMed]
- Low, P.S.; Henne, W.A.; Doorneweerd, D.D. Discovery and development of folic-acid-based receptor targeting for imaging and therapy of cancer and inflammatory diseases. Acc. Chem. Res. 2008, 41, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Miller, T.A.; Witter, D.J.; Belvedere, S. Histone deacetylase inhibitors. J. Med. Chem. 2003, 46, 5097–5116. [Google Scholar] [CrossRef] [PubMed]
- Mottamal, M.; Zheng, S.; Huang, T.L.; Wang, G. Histone deacetylase inhibitors in clinical studies as templates for new anticancer agents. Molecules 2015, 20, 3898–3941. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Furumai, R.; Nishiyama, M.; Komatsu, Y.; Nishino, N.; Horinouchi, S. Histone deacetylase as a new target for cancer chemotherapy. Cancer Chemother. Pharmacol. 2001, 48, S20–S26. [Google Scholar] [CrossRef] [PubMed]
- Lane, A.A.; Chabner, B.A. Histone deacetylase inhibitors in cancer therapy. J. Clin. Oncol. 2009, 27, 5459–5468. [Google Scholar] [CrossRef] [PubMed]
- Karpozilos, A.; Pavlidis, N. The treatment of cancer in Greek antiquity. Eur. J. Cancer 2004, 40, 2033–2040. [Google Scholar] [CrossRef] [PubMed]
- Hamidpour, R.; Hamidpour, S.; Hamidpour, M.; Shahlari, M. Frankincense (乳香 Rǔ Xiāng; Boswellia species): From the selection of traditional applications to the novel phytotherapy for the prevention and treatment of serious diseases. J. Tradit. Complement. Med. 2013, 3, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Frank, M.B.; Yang, Q.; Osban, J.; Azzarello, J.T.; Saban, M.R.; Saban, R.; Ashley, R.A.; Welter, J.C.; Fung, K.M.; Lin, H.K. Frankincense oil derived from Boswellia carteri induces tumor cell specific cytotoxicity. BMC Complement. Altern. Med. 2009, 9, 6. [Google Scholar] [CrossRef] [PubMed]
- Forouzandeh, S.; Naghsh, N.; Salimi, S.; Jahantigh, D. Cytotoxic effect of Boswellia serrata hydroalcoholic extract on human cervical carcinoma epithelial cell line. Med. Lab. J. 2014, 8, 7–13. [Google Scholar]
- Hakkim, F.L.; Al-Buloshi, M.; Al-Sabahi, J. Frankincense derived heavy terpene cocktail boosting breast cancer cell (MDA-MB-231) death in vitro. Asian Pac. J. Trop. Biomed. 2015, 5, 824–828. [Google Scholar] [CrossRef]
- Eichhorn, T.; Greten, H.J.; Efferth, T. Molecular determinants of the response of tumor cells to boswellic acids. Pharmaceuticals 2011, 4, 1171–1182. [Google Scholar] [CrossRef]
- Agrawal, S.S.; Saraswati, S.; Mathur, R.; Pandey, M. Antitumor properties of Boswellic acid against Ehrlich ascites cells bearing mouse. Food Chem. Toxicol. 2011, 49, 1924–1934. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Sung, B.; Shen, Y.; Hur, K.; Link, A.; Boland, C.R.; Aggarwal, B.B.; Goel, A. Boswellic acid exerts antitumor effects in colorectal cancer cells by modulating expression of the let-7 and miR-200 microRNA family. Carcinogenesis 2012, 33, 2441–2449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, V.R.; Prasad, S.; Sung, B.; Kannappan, R.; Aggarwal, B.B. Targeting inflammatory pathways by triterpenoids for prevention and treatment of cancer. Toxins 2010, 2, 2428–2466. [Google Scholar] [CrossRef] [PubMed]
- Ranzato, E.; Martinotti, S.; Volante, A.; Tava, A.; Masini, M.A.; Burlando, B. The major Boswellia serrata active 3-acetyl-11-keto-β-boswellic acid strengthens interleukin-1α upregulation of matrix metalloproteinase-9 via JNK MAP kinase activation. Phytomedicine 2017, 36, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Seckl, J.R.; Walker, B.R. Minireview: 11β-Hydroxysteroid dehydrogenase type 1—A tissue-specific amplifier of glucocorticoid action. Endocrinology 2001, 142, 1371–1376. [Google Scholar] [CrossRef] [PubMed]
- Morton, N.M.; Paterson, J.M.; Masuzaki, H.; Holmes, M.C.; Staels, B.; Fievet, C.; Walker, B.R.; Flier, J.S.; Mullins, J.J.; Seckl, J.R. Novel adipose tissue-mediated resistance to diet-induced visceral obesity in 11β-hydroxysteroid dehydrogenase type 1-deficient mice. Diabetes 2004, 53, 931–938. [Google Scholar] [CrossRef] [PubMed]
- Hosfield, D.J.; Wu, Y.; Skene, E.J.; Hilgers, M.; Jennings, A.; Snell, G.P.; Aertgeerts, K. Conformational flexibility in crystal structures of human 11β-hydroxysteroid dehydrogenase type I provide insights into glucocorticoid interconversion and enzyme regulation. J. Biol. Chem. 2005, 280, 4639–4648. [Google Scholar] [CrossRef] [PubMed]
- Wan, Z.K.; Chenail, E.; Xiang, J.; Li, H.Q.; Ipek, M.; Bard, J.; Svenson, K.; Mansour, T.S.; Xu, X.; Tian, X.; et al. Efficacious 11β-hydroxysteroid dehydrogenase type I inhibitors in the diet-induced obesity mouse model. J. Med. Chem. 2009, 52, 5449–5461. [Google Scholar] [CrossRef] [PubMed]
- Ahangarpour, A.; Heidari, H.; Fatemeh, R.A.A.; Pakmehr, M.; Shahbazian, H.; Ahmadi, I.; Mombeini, Z.; Mehrangiz, B.H. Effect of Boswellia serrata supplementation on blood lipid, hepatic enzymes and fructosamine levels in type2 diabetic patients. J. Diabetes Metab. Disord. 2014, 13, 29. [Google Scholar] [CrossRef] [PubMed]
- Khalili, N.; Fereydoonzadeh, R.; Mohtashami, R.; Mehrzadi, S.; Heydari, M.; Huseini, H.F. Silymarin, olibanum, and nettle, a mixed herbal formulation in the treatment of type II diabetes: A randomized, double-blind, placebo-controlled, clinical trial. J. Evid. Based Complement. Altern. Med. 2017, 22, 603–608. [Google Scholar] [CrossRef] [PubMed]
- Shehata, A.M.; Quintanilla-Fend, L.; Bettio, S.; Singh, C.B.; Ammon, H.P.T. Prevention of multiple low-dose streptozotocin (MLD-STZ) diabetes in mice by an extract from gum resin of Boswellia serrata (BE). Phytomedicine 2011, 18, 1037–1044. [Google Scholar] [CrossRef] [PubMed]
- Kavitha, J.V.; Rosario, J.F.; Chandran, J.; Anbu, P.; Bakkiyanathan. Hypoglycemic and other related effects of Boswellia glabra in alloxan-induced diabetic rats. Indian J. Physiol. Pharmacol. 2007, 51, 29–39. [Google Scholar] [PubMed]
- Abe, I.; Rohmer, M.; Prestwich, G.D. Enzymatic cyclization of squalene and oxidosqualene to sterols and triterpenes. Chem. Rev. 1993, 93, 2189–2206. [Google Scholar] [CrossRef]
- Nes, W.D. Biosynthesis of cholesterol and other sterols. Chem. Rev. 2011, 111, 6423–6451. [Google Scholar] [CrossRef] [PubMed]
- Huff, M.W.; Telford, D.E. Lord of the rings—The mechanism for oxidosqualene:lanosterol cyclase becomes crystal clear. Trends Pharmacol. Sci. 2005, 26, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Rabelo, V.W.H.; Romeiro, N.C.; Abreu, P.A. Design strategies of oxidosqualene cyclase inhibitors: Targeting the sterol biosynthetic pathway. J. Steroid Biochem. Mol. Biol. 2017, 171, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Lenhart, A.; Reinert, D.J.; Aebi, J.D.; Dehmlow, H.; Morand, O.H.; Schulz, G.E. Binding structures and potencies of oxidosqualene cyclase inhibitors with the homologous squalene-hopene cyclase. J. Med. Chem. 2003, 46, 2083–2092. [Google Scholar] [CrossRef] [PubMed]





















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ligand | PDB a | Edock | Target Protein |
|---|---|---|---|
| Boscartin A | 1e66 | −119.7 | Torpedo californica acetylcholinesterase (TcAChE) |
| 2cek | −123.7 | Torpedo californica acetylcholinesterase (TcAChE) | |
| 4bdt | −113.5 | human acetylcholinesterase (HsAChE) | |
| −79.2 | Median docking energy | ||
| Boscartin B | 1h0g | −107.8 | Serratia marcescens chitinase B (SmChiB) |
| 2ew5 | −104.6 | Helicobacter pylori peptide deformylase (HpPDF) | |
| 3tfu | −107.5 | Mycobacterium tuberculosis 7,8-diaminopelargonic acid synthase (MtBioA) | |
| 3v0o | −110.4 | human fucosylgalactoside α N-acetylgalactosaminyltransferase (HsGTA) | |
| −80.5 | Median docking energy | ||
| Boscartin C | 1ahg | −112.5 | Escherichia coli aspartate aminotransferase (EcAspTA) |
| 1fv0 | −108.2 | Daboia russelii (Russell’s viper) phospholipase A2 (DrPLA2) | |
| 1q83 | −110.4 | murine acetylcholinesterase (MmAChE) | |
| 4g1n | −106.5 | human pyruvate kinase isozyme M2 (HsPKM2) | |
| −91.2 | Median docking energy | ||
| Boscartin D | 1ahg | −114.4 | Escherichia coli aspartate aminotransferase (EcAspTA) |
| 1xzq | −106.3 | Thermotoga maritima GTP-binding protein TrmE (TmTrmE) | |
| 2cek | −115.3 | Torpedo californica acetylcholinesterase (TcAChE) | |
| −82.0 | Median docking energy | ||
| Boscartin E | 1ahg | −110.4 | Escherichia coli aspartate aminotransferase (EcAspTA) |
| 1e66 | −111.7 | Torpedo californica acetylcholinesterase (TcAChE) | |
| 2ew5 | −107.5 | Helicobacter pylori peptide deformylase (HpPDF) | |
| −68.4 | Median docking energy | ||
| Boscartin F | 2ew5 | −109.5 | Helicobacter pylori peptide deformylase (HpPDF) |
| 3i6m | −107.3 | Torpedo californica acetylcholinesterase (TcAChE) | |
| 4g1n | −108.5 | human pyruvate kinase isozyme M2 (HsPKM2) | |
| −82.9 | Median docking energy | ||
| Boscartin G | 1e66 | −126.4 | Torpedo californica acetylcholinesterase (TcAChE) |
| 2cek | −116.8 | Torpedo californica acetylcholinesterase (TcAChE) | |
| 4bdt | −118.7 | human acetylcholinesterase (HsAChE) | |
| −89.9 | Median docking energy | ||
| Boscartin H | 1fv0 | −106.9 | Daboia russelii (Russell’s viper) phospholipase A2 (DrPLA2) |
| 1w1t | −106.0 | Serratia marcescens chitinase B (SmChiB) | |
| 2gyw | −105.9 | murine acetylcholinesterase (MmAChE) | |
| 3v0o | −109.3 | human fucosylgalactoside α N-acetylgalactosaminyltransferase (HsGTA) | |
| −88.4 | Median docking energy | ||
| Incensole | 2cek | −111.9 | Torpedo californica acetylcholinesterase (TcAChE) |
| 3ugr | −109.7 | human aldo-keto reductase 1C3 (HsAKR1C3) | |
| 1h0g | −102.4 | Serratia marcescens chitinase B (SmChiB) | |
| −78.1 | Median docking energy | ||
| Incensole acetate | 1ahg | −106.9 | Escherichia coli aspartate aminotransferase (EcAspTA) |
| 3jun | −103.7 | Burkholderia cepacia phenazine biosynthesis protein A/B (BcPhzA/B) | |
| −69.5 | Median docking energy | ||
| Incensole oxide | 1ahg | −108.6 | Escherichia coli aspartate aminotransferase (EcAspTA) |
| 2cek | −103.1 | Torpedo californica acetylcholinesterase (TcAChE) | |
| 3jup | −103.5 | Burkholderia cepacia phenazine biosynthesis protein A/B (BcPhzA/B) | |
| 3mee | −106.7 | HIV-1 reverse transcriptase (HIV-1 RT) | |
| −74.6 | Median docking energy | ||
| Incensole oxide acetate | 1ahg | −119.0 | Escherichia coli aspartate aminotransferase (EcAspTA) |
| 1q83 | −109.6 | murine acetylcholinesterase (MmAChE) | |
| 2cek | −110.4 | Torpedo californica acetylcholinesterase (TcAChE) | |
| 3i6m | −114.3 | Torpedo californica acetylcholinesterase (TcAChE) | |
| 3mee | −113.5 | HIV-1 reverse transcriptase (HIV-1 RT) | |
| −91.9 | Median docking energy | ||
| Isoincensole oxide | 3jup | −103.3 | Burkholderia cepacia phenazine biosynthesis protein A/B (BcPhzA/B) |
| −80.5 | Median docking energy | ||
| Isoincensolol | 1fv0 | −108.2 | Daboia russelii (Russell’s viper) phospholipase A2 (DrPLA2) |
| 1jus | −107.9 | Staphylococcus aureus multidrug binding protein (SaQacR) | |
| 2qp4 | −102.2 | human dehydroepiandrosterone sulfotransferase (HsSULT2A1) | |
| −74.0 | Median docking energy | ||
| Serratol | 1e66 | −104.1 | Torpedo californica acetylcholinesterase (TcAChE) |
| 2cek | −106.1 | Torpedo californica acetylcholinesterase (TcAChE) | |
| 2ew5 | −103.2 | Helicobacter pylori peptide deformylase (HpPDF) | |
| 4bdt | −103.0 | human acetylcholinesterase (HsAChE) | |
| −78.1 | Median docking energy | ||
| Verticillatriene | 1w4l | −104.3 | Torpedo californica acetylcholinesterase (TcAChE) |
| 3i6m | −100.8 | Torpedo californica acetylcholinesterase (TcAChE) | |
| 3i6z | −103.2 | Torpedo californica acetylcholinesterase (TcAChE) | |
| −63.6 | Median docking energy |
| Ligand | TcAChE | TcAChE | TcAChE | TcAChE | TcAChE | TcAChE | MmAChE | MmAChE | HsAChE |
|---|---|---|---|---|---|---|---|---|---|
| 1e66 | 1h22 | 1w4l | 2cek | 3i6m | 3i6z | 1q83 | 2gyw | 4bdt | |
| Boscartin A | −119.7 | −98.4 | −99.5 | −123.7 | −98.2 | −100.6 | −95.9 | −101.4 | −113.5 |
| Boscartin B | −95.1 | −98.8 | −92.5 | −106.3 | −99.2 | −94.5 | −94.3 | −85.8 | −106.3 |
| Boscartin C | −87.7 | −90.8 | −94.7 | −96.9 | −104.1 | −94.2 | −110.4 | −94.0 | −86.6 |
| Boscartin D | −99.7 | −102.8 | −93.1 | −115.3 | −103.7 | −93.9 | −92.5 | −93.6 | −90.2 |
| Boscartin E | −111.7 | −96.5 | −94.5 | −108.7 | −95.5 | −92.9 | −86.0 | −94.8 | −90.6 |
| Boscartin F | −93.2 | −97.0 | −98.5 | −104.8 | −107.3 | −95.8 | −101.1 | −82.4 | −77.3 |
| Boscartin G | −126.4 | −99.4 | −94.5 | −116.8 | −101.5 | −95.6 | −111.1 | −94.1 | −118.7 |
| Boscartin H | −100.1 | −106.3 | −103.6 | −105.6 | −103.0 | −104.2 | −98.8 | −105.9 | −97.8 |
| Incensole | −89.7 | −92.1 | −89.5 | −111.9 | −94.1 | −89.6 | −90.3 | −87.7 | −53.4 |
| Incensole acetate | −88.6 | −97.2 | −102.8 | −102.5 | −102.3 | −101.7 | −87.5 | −85.0 | −42.3 |
| Incensole oxide | −89.6 | −93.2 | −92.7 | −103.1 | −96.9 | −89.3 | −101.9 | −95.5 | −93.5 |
| Incensole oxide acetate | −109.8 | −98.7 | −96.8 | −110.4 | −114.3 | −95.8 | −109.6 | −97.7 | −102.6 |
| Isoincensole oxide | −92.9 | −98.9 | −96.4 | −103.6 | −98.1 | −89.9 | −89.9 | −90.3 | −94.1 |
| Isoincensolol | −88.5 | −99.5 | −92.4 | −101.5 | −98.3 | −91.2 | −77.0 | −80.4 | −73.6 |
| Serratol | −104.1 | −95.7 | −92.2 | −106.1 | −94.5 | −88.1 | −96.4 | −89.3 | −104.0 |
| Verticillatriene | −82.5 | −91.2 | −104.3 | −85.3 | −100.8 | −103.2 | −73.9 | −98.1 | −61.2 |
| Ligand | EcAspTA | SmChiB | SmChiB | SmChiB | SaQacR | SaQacR | SaQacR | SaQacR | HpPDF | BcPhzA/B | BcPhzA/B | MtBioA |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1ahg | 1h0g | 1w1t | 3wd2 | 1jus | 1rpw | 3br2 | 3bti | 2ew5 | 3jun | 3jup | 3tfu | |
| Boscartin A | −116.9 | −93.6 | −99.4 | −96.3 | −99.4 | −104.1 | −94.9 | −95.7 | −101.8 | −100.3 | −107.8 | −104.9 |
| Boscartin B | −98.2 | −107.8 | −103.7 | −91.5 | −98.1 | −106.5 | −94.7 | −96.5 | −104.6 | −98.4 | −93.9 | −107.5 |
| Boscartin C | −112.5 | −96.3 | −97.3 | −90.6 | −100.0 | −98.5 | −98.4 | −102.3 | −106.2 | −99.3 | −106.4 | −100.4 |
| Boscartin D | −114.4 | −99.7 | −105.3 | −91.5 | −95.2 | −106.2 | −100.1 | −96.3 | −104.9 | −98.0 | −95.8 | −99.3 |
| Boscartin E | −110.4 | −101.0 | −105.9 | −105.4 | −99.2 | −101.3 | −95.3 | −95.3 | −107.5 | −84.7 | −96.7 | −107.7 |
| Boscartin F | −99.7 | −97.3 | −103.7 | −89.9 | −101.0 | −95.7 | −94.4 | −85.2 | −109.5 | −87.7 | −98.0 | −95.1 |
| Boscartin G | −105.5 | −90.8 | −93.0 | −93.8 | −95.1 | −98.2 | −92.2 | −88.7 | −112.7 | −95.4 | −92.6 | −108.5 |
| Boscartin H | −104.1 | −103.6 | −106.0 | −104.4 | −97.2 | −98.8 | −97.4 | −94.5 | −100.9 | −93.4 | −101.5 | −99.5 |
| Incensole | −104.3 | −102.4 | −101.1 | −96.2 | −85.0 | −96.3 | −90.7 | −95.0 | −94.0 | −92.9 | −93.0 | −95.7 |
| Incensole acetate | −109.9 | −102.3 | −96.2 | −98.1 | −96.6 | −104.5 | −96.1 | −98.8 | −96.2 | −103.7 | −91.5 | −96.2 |
| Incensole oxide | −108.6 | −92.0 | −93.5 | −90.3 | −97.4 | −99.8 | −94.2 | −95.6 | −101.4 | −97.1 | −103.5 | −103.5 |
| Incensole oxide acetate | −119.0 | −101.1 | −98.9 | −96.4 | −96.7 | −98.7 | −99.2 | −100.0 | −106.9 | −105.5 | −99.4 | −100.0 |
| Isoincensole oxide | −96.2 | −87.6 | −93.5 | −89.9 | −98.5 | −97.5 | −98.5 | −92.7 | −88.9 | −87.5 | −103.3 | −91.2 |
| Isoincensolol | −87.3 | −91.2 | −88.9 | −87.9 | −107.9 | −90.0 | −99.6 | −87.8 | −99.7 | −91.5 | −100.0 | −95.2 |
| Serratol | −90.2 | −90.1 | −89.9 | −82.2 | −90.1 | −91.4 | −97.6 | −89.1 | −103.2 | −87.4 | −92.7 | −93.4 |
| Verticillatriene | −74.0 | −69.5 | −69.0 | −69.5 | −81.2 | −87.2 | −85.5 | −78.5 | −77.3 | −76.5 | −89.9 | −84.4 |
| Ligand | 1eet | 2hnz | 3irx | 3is9 | 3mee | 3lal | 3t19 |
|---|---|---|---|---|---|---|---|
| Boscartin A | −99.1 | −101.7 | −96.8 | −93.8 | −96.0 | −69.4 | −75.4 |
| Boscartin B | −94.1 | −98.5 | −100.2 | −94.7 | −95.6 | −72.2 | −97.8 |
| Boscartin C | −104.2 | −91.5 | −97.0 | −101.3 | −100.5 | −87.8 | −84.0 |
| Boscartin D | −89.2 | −100.5 | −97.1 | −97.1 | −96.4 | −78.7 | −83.2 |
| Boscartin E | −97.2 | −100.6 | −91.7 | −97.0 | −89.4 | −60.8 | −72.0 |
| Boscartin F | −88.9 | −93.8 | −98.1 | −98.1 | −95.1 | −88.1 | −69.4 |
| Boscartin G | −92.8 | −88.7 | −97.6 | −95.4 | −113.8 | +9.3 | −82.3 |
| Boscartin H | −98.2 | −90.4 | −96.9 | −97.1 | −98.2 | −86.9 | −56.2 |
| Incensole | −83.8 | −101.5 | −88.8 | −88.3 | −86.8 | −84.0 | −94.7 |
| Incensole acetate | −95.6 | −92.6 | −97.9 | −97.0 | −84.6 | −95.9 | −91.6 |
| Incensole oxide | −105.1 | −93.2 | −94.9 | −98.7 | −106.7 | −81.7 | −79.0 |
| Incensole oxide acetate | −107.2 | −84.5 | −106.8 | −106.2 | −113.5 | −87.8 | −87.1 |
| Isoincensole oxide | −97.8 | −86.5 | −89.4 | −90.2 | −98.7 | −77.5 | −57.7 |
| Isoincensolol | −95.2 | −85.6 | −98.2 | −85.4 | −85.9 | −70.6 | −72.5 |
| Serratol | −90.4 | −87.9 | −86.8 | −89.3 | −90.1 | −56.0 | −48.9 |
| Verticillatriene | −81.6 | −67.1 | −78.4 | −81.0 | −82.5 | −76.9 | −25.5 |
| Ligand | PDB a | Edock | Target Protein |
|---|---|---|---|
| Boscartol A | 3hfj | −124.2 | Bacillus anthracis nucleotide adenyltransferase (BaNadD) |
| 4kn0 | −125.6 | human folate receptor β (HsFRβ) | |
| 4kn1 | −127.5 | human folate receptor β (HsFRβ) | |
| −89.7 | Median docking energy | ||
| Boscartol B | 3hfj | −124.2 | Bacillus anthracis nucleotide adenyltransferase (BaNadD) |
| 4kn0 | −127.8 | human folate receptor β (HsFRβ) | |
| 4kn1 | −126.8 | human folate receptor β (HsFRβ) | |
| −91.6 | Median docking energy | ||
| Boscartol C | 3bt9 | −120.2 | Staphylococcus aureus multidrug binding protein (SaQacR) |
| −85.8 | Median docking energy | ||
| (15R)-Boscartol D | 2cek | −120.9 | Torpedo californica acetylcholine esterase (TcAChE) |
| 3bt9 | −122.7 | Staphylococcus aureus multidrug binding protein (SaQacR) | |
| 3lal | −125.5 | HIV-1 reverse transcriptase | |
| 3t19 | −123.1 | HIV-1 reverse transcriptase | |
| −87.6 | Median docking energy | ||
| (15S)-Boscartol D | 3lal | −120.6 | HIV-1 reverse transcriptase |
| −81.8 | Median docking energy | ||
| Boscartol E | 1s9d | −125.9 | bovine guanine nucleotide exchange factor (BtGEF) |
| 3hjf | −135.1 | Bacillus anthracis nucleotide adenyltransferase (BaNadD) | |
| 4b80 | −124.1 | murine acetylcholinesterase (MmAChE) | |
| 4kn0 | −124.2 | human folate receptor β (HsFRβ) | |
| −85.0 | Median docking energy | ||
| Boscartol F | 1s9d | −124.5 | bovine guanine nucleotide exchange factor (BtGEF) |
| 3hfj | −123.1 | Bacillus anthracis nucleotide adenyltransferase (BaNadD) | |
| 4kn1 | −122.3 | human folate receptor β (HsFRβ) | |
| −84.6 | Median docking energy | ||
| Boscartol G | 3hfj | −125.8 | Bacillus anthracis nucleotide adenylyltransferase (BaNadD) |
| −80.3 | Median docking energy | ||
| Boscartol H | 1dx4 | −123.6 | Drosophila melanogaster acetylcholine esterase (DmAChE) |
| 3bt9 | −122.0 | Staphylococcus aureus multidrug binding protein (SaQacR) | |
| 3hfj | −127.9 | Bacillus anthracis nucleotide adenyltransferase (BaNadD) | |
| 4kn0 | −124.6 | human folate receptor β (HsFRβ) | |
| −85.9 | Median docking energy | ||
| Boscartol I | 3hfj | −133.8 | Bacillus anthracis nucleotide adenyltransferase (BaNadD) |
| 3p2v | −121.1 | human aldose reductase (HsAR) | |
| 4kn0 | −127.3 | human folate receptor β (HsFRβ) | |
| −91.4 | Median docking energy | ||
| Olibanumol D | 4kn1 | −120.6 | human folate receptor β (HsFRβ) |
| −78.4 | Median docking energy |
| Ligand | PDB a | Edock | Target |
|---|---|---|---|
| (20S)-3,7-Dioxotirucalla-8,24-dien-21-oic acid | 3l3m | −141.8 | Human poly(ADP-ribose) polymerase-1 (HsPARP-1) (anticancer target) |
| 3ua9 | −140.3 | Human tankyrase-2 (HsTANK2) = human poly(ADP-ribose) polymerase-5b (HsPARP-5b) (antitumor target) | |
| 3g49 | −132.4 | Cavia porcellus 11β-hydroxysteroid dehydrogenase type 1 (Cp11βHSD1) (diabetes target) | |
| 2b03 | −127.4 | Porcine pancreatic phospholipase A2 (SsPLA2) (anti-inflammatory target) | |
| −91.3 | Median docking energy | ||
| 11-Ethoxy-β-boswellic acid | 3i6m | −124.6 | Torpedo californica acetylcholinesterase (TcAChE) (Alzheimer’s target) |
| −63.0 | Median docking energy | ||
| 11-Keto-β-boswellic acid | 4b84 | −125.3 | Murine acetylcholinesterase (MmAChE) (Alzheimer’s target) |
| 3i6m | −120.8 | Torpedo californica acetylcholinesterase (TcAChE) (Alzheimer’s target) | |
| −59.9 | Median docking energy | ||
| 12-Ursen-3,11-dione | 1h22 | −118.8 | Torpedo californica acetylcholinesterase (TcAChE) (Alzheimer’s target) |
| −64.2 | Median docking energy | ||
| 12-Ursen-3,24-diol | 1h22 | −123.8 | Torpedo californica acetylcholinesterase (TcAChE) (Alzheimer’s target) |
| +17.4 | Median docking energy | ||
| 2,3-Dihydroxy-12-ursen-24-oic acid | 4b84 | −128.0 | Murine acetylcholinesterase (MmAChE) (Alzheimer’s target) |
| +16.2 | Median docking energy | ||
| 20,22-Epoxytirucall-24-en-3-one | 3l3m | −133.5 | Human poly(ADP-ribose) polymerase-1 (HsPARP-1) (anticancer target) |
| 3g49 | −125.0 | Cavia porcellus 11β-hydroxysteroid dehydrogenase type 1 (Cp11βHSD1) (diabetes target) | |
| −88.9 | Median docking energy | ||
| 24-Nor-3,12-ursadien-11-one | 1h22 | −121.0 | Torpedo californica acetylcholinesterase (TcAChE) (Alzheimer’s target) |
| 4b84 | −120.9 | Murine acetylcholinesterase (MmAChE) (Alzheimer’s target) | |
| +30.9 | Median docking energy | ||
| 24-Nor-3,9(11),12-oleanatriene | 3lz6 | −112.6 | Cavia porcellus 11β-hydroxysteroid dehydrogenase type 1 (Cp11βHSD1) (diabetes target) |
| −31.2 | Median docking energy | ||
| 24-Nor-3,9(11),12-ursatriene | 1h22 | −118.4 | Torpedo californica acetylcholinesterase (TcAChE) (Alzheimer’s target) |
| +15.1 | Median docking energy | ||
| 3-Acetoxy-12,20(29)-lupadien-24-oic acid | 1uk1 | −118.4 | Human poly(ADP-ribose) polymerase-1 (HsPARP-1) (anticancer target) |
| 3bti | −117.9 | Staphylococcus aureus multidrug binding protein (SaQacR) | |
| +16.4 | Median docking energy | ||
| 3-Acetoxy-20(29)-lupen-24-oic acid (= 3-Acetyl lupeolic acid) | 1cgl | −117.5 | Human fibroblast collagenase (HsMMP-1) (arthritis target) |
| +8.0 | Median docking energy | ||
| 3β-Acetoxy-20S,24R-dihydroxydammar-25-ene b | 2aba | −144.7 | Enterobacter cloacae pentaerythritol tetranitrate reductase (EcPETNR) (antibacterial target) |
| 3lz6 | −141.8 | Cavia porcellus 11β-hydroxysteroid dehydrogenase type 1 (Cp11βHSD1) (diabetes target) | |
| 1h36 | −136.4 | Alicyclobacillus acidocardarius oxidosqualene cyclase (Aa OSC) (cholesterol-lowering) | |
| −64.6 | Median docking energy | ||
| 3β-Acetoxy-20S,24S-dihydroxydammar-25-ene b | 2aba | −149.9 | Enterobacter cloacae pentaerythritol tetranitrate reductase (EcPETNR) (antibacterial target) |
| 3g49 | −139.7 | Cavia porcellus 11β-hydroxysteroid dehydrogenase type 1 (Cp11βHSD1) (diabetes target) | |
| 3lz6 | −136.2 | Cavia porcellus 11β-hydroxysteroid dehydrogenase type 1 (Cp11βHSD1) (diabetes target) | |
| 3tfu | −134.5 | Mycobacterium tuberculosis 7,8-diaminopelargonic acid synthase (MtBioA) | |
| −73.7 | Median docking energy | ||
| 3-Acetoxy-5,12-ursadien-24-oic acid | 3i6m | −131.7 | Torpedo californica acetylcholinesterase (TcAChE) (Alzheimer’s target) |
| −66.7 | Median docking energy | ||
| 3-Acetyl-11-keto-β-boswellic acid | 3i6m | −129.7 | Torpedo californica acetylcholinesterase (TcAChE) (Alzheimer’s target) |
| −70.7 | Median docking energy | ||
| 3-Acetyl-11α-methoxy-β-boswellic_acid | 3i6m | −129.0 | Torpedo californica acetylcholinesterase (TcAChE) (Alzheimer’s target) |
| −72.2 | Median docking energy | ||
| 3-Acetyl-9,11-dehydro-β-boswellic_acid | 3i6m | −124.0 | Torpedo californica acetylcholinesterase (TcAChE) (Alzheimer’s target) |
| −64.4 | Median docking energy | ||
| 3-Acetyl-α-boswellic acid | 2b03 | −112.7 | Porcine pancreatic phospholipase A2 (SsPLA2) (anti-inflammatory target) |
| −64.8 | Median docking energy | ||
| 3-Acetyl-β-boswellic acid | 3i6m | −128.3 | Torpedo californica acetylcholinesterase (TcAChE) (Alzheimer’s target) |
| −71.5 | Median docking energy | ||
| 3-Hydroxy-20(29)-lupen-24-oic acid (= Lupeolic acid) | 3bt9 | −119.3 | Staphylococcus aureus multidrug binding protein (SaQacR) |
| −55.8 | Median docking energy | ||
| 3-Oxotirucalla-7,9(11),24-trien-21-oic acid | 3ua9 | −151.0 | Human tankyrase-2 (HsTANK2) = human poly(ADP-ribose) polymerase-5b (HsPARP-5b) (antitumor target) |
| 3l3m | −137.6 | Human poly(ADP-ribose) polymerase-1 (HsPARP-1) (anticancer target) | |
| 3h6k | −130.9 | Human 11β-hydroxysteroid dehydrogenase type 1 (11βHSD1) (diabetes target) | |
| 1w6j | −127.5 | Human oxidosqualene cyclase (HsOSC) (hypercholesterolemia target) | |
| 3bt9 | −127.1 | Staphylococcus aureus multidrug binding protein (SaQacR) | |
| −96.8 | Median docking energy | ||
| 3β-Hydroxytirucalla-8,24-dien-21-oic acid | 3ua9 | −144.5 | Human tankyrase-2 (HsTANK2) = human poly(ADP-ribose) polymerase-5b (HsPARP-5b) (antitumor target) |
| 3l3m | −133.0 | Human poly(ADP-ribose) polymerase-1 (HsPARP-1) (anticancer target) | |
| 3g49 | −132.3 | Cavia porcellus 11β-hydroxysteroid dehydrogenase type 1 (Cp11βHSD1) (diabetes target) | |
| 2ilt | −129.1 | Human 11β-hydroxysteroid-dehydrogenase (Hs11β-HSDH) (diabetes target) | |
| 4krs | −127.7 | Human tankyrase-1 (HsTANK1) (anticancer target) | |
| 4l0i | −127.3 | Human tankyrase-2 (HsTANK2) (anticancer target) | |
| −85.6 | Median docking energy | ||
| 3α-Acetoxytirucalla-7,24-dien-21-oic acid | 3ua9 | −147.1 | Human tankyrase-2 (HsTANK2) = human poly(ADP-ribose) polymerase-5b (HsPARP-5b) (antitumor target) |
| 4krs | −134.1 | Human tankyrase-1 (HsTANK1) (anticancer target) | |
| 1cgl | −129.9 | Human fibroblast collagenase (HsMMP-1) (arthritis target) | |
| 4gv0 | −128.4 | Human poly(ADP-ribose) polymerase-1 (HsPARP-1) (anticancer target) | |
| 4l0i | −128.4 | Human tankyrase-2 (HsTANK2) (anticancer target) | |
| 3lep | −126.8 | Human aldose reductase (HsAR) (diabetes target) | |
| −102.1 | Median docking energy | ||
| 3α-Hydroxytirucalla-7,24-dien-21-oic acid | 3ua9 | −154.3 | Human tankyrase-2 (HsTANK2) = human poly(ADP-ribose) polymerase-5b (HsPARP-5b) (antitumor target) |
| 3g49 | −128.9 | Cavia porcellus 11β-hydroxysteroid dehydrogenase type 1 (Cp11βHSD1) (diabetes target) | |
| 3l3m | −126.7 | Human poly(ADP-ribose) polymerase-1 (HsPARP-1) (anticancer target) | |
| 3tfu | −126.4 | Mycobacterium tuberculosis 7,8-diaminopelargonic acid synthase (MtBioA) | |
| −88.7 | Median docking energy | ||
| 3β-Acetoxydammar-24-ene-16β,20R-diol | 3lz6 | −149.6 | Cavia porcellus 11β-hydroxysteroid dehydrogenase type 1 (Cp11βHSD1) (diabetes target) |
| 2aba | −141.1 | Enterobacter cloacae pentaerythritol tetranitrate reductase (EcPETNR) (antibacterial target) | |
| 1h36 | −139.2 | Alicyclobacillus acidocardarius oxidosqualene cyclase (AaOSC) (cholesterol-lowering) | |
| 1s0x | −131.2 | Human retinoic acid-related orphan receptor α (HsRORα) (may regulate lipid metabolism) | |
| −54.0 | Median docking energy | ||
| 3β-Acetoxylup-20(29)-en-11β-ol | 1uk1 | −121.9 | Human poly(ADP-ribose) polymerase-1 (HsPARP-1) (anticancer target) |
| −50.4 | Median docking energy | ||
| 4,23-Dihydroburic acid | 1h22 | −135.8 | Torpedo californica acetylcholinesterase (TcAChE) (Alzheimer’s target) |
| 4b84 | −127.8 | Murine acetylcholinesterase (MmAChE) (Alzheimer’s target) | |
| 1jtx | −125.1 | Staphylococcus aureus multidrug binding protein (SaQacR) | |
| +11.7 | Median docking energy | ||
| 6,7-Epoxy-9(11)-oleanen-3-ol | 4hai | −111.6 | Human soluble epoxide hydrolase (HsEPHX2) (anti-inflammatory target) |
| −60.7 | Median docking energy | ||
| 6,7-Epoxy-9(11)-oleanen-3-one | 1ry0 | −113.7 | Human prostaglandin F synthase (HsPGFS) (hypertension target) |
| −49.1 | Median docking energy | ||
| 9,11-Dehydro-β-boswellic acid | 1qvu | −114.3 | Staphylococcus aureus multidrug binding protein (SaQacR) |
| −60.8 | Median docking energy | ||
| Boscartene L | 3l3m | −131.0 | Human poly(ADP-ribose) polymerase-1 (HsPARP-1) (anticancer target) |
| −87.6 | Median docking energy | ||
| Boscartene M | 3l3m | −133.7 | Human poly(ADP-ribose) polymerase-1 (HsPARP-1) (anticancer target) |
| −89.4 | Median docking energy | ||
| Boscartene N | 3ua9 | −157.2 | Human tankyrase-2 (HsTANK2) = human poly(ADP-ribose) polymerase-5b (HsPARP-5b) (antitumor target) |
| 3bt9 | −136.7 | Staphylococcus aureus multidrug binding protein (SaQacR) | |
| 3l3m | −133.3 | Human poly(ADP-ribose) polymerase-1 (HsPARP-1) (anticancer target) | |
| 1xl5 | −125.2 | HIV-1 protease | |
| −95.1 | Median docking energy | ||
| Dammarenediol II | 1w6k | −135.1 | Human oxidosqualene cyclase (HsOSC) (hypercholesterolemia target) |
| 3lz6 | −134.6 | Cavia porcellus 11β-hydroxysteroid dehydrogenase type 1 (Cp11βHSD1) (diabetes target) | |
| 1h36 | −129.2 | Alicyclobacillus acidocardarius oxidosqualene cyclase (AaOSC) (cholesterol-lowering) | |
| −72.1 | Median docking energy | ||
| Dammarenediol II acetate | 3lz6 | −142.8 | Cavia porcellus 11β-hydroxysteroid dehydrogenase type 1 (Cp11βHSD1) (diabetes target) |
| 1h36 | −138.9 | Alicyclobacillus acidocardarius oxidosqualene cyclase (AaOSC) (cholesterol-lowering) | |
| 2aba | −136.4 | Enterobacter cloacae pentaerythritol tetranitrate reductase (EcNETNR) (antibacterial target) | |
| 4kn0 | −132.3 | Human folate receptor β (HsFRβ) (anticancer target) | |
| −77.7 | Median docking energy | ||
| Eupha-2,8,22-triene-20,24R-diol b | 3l3m | −139.8 | Human poly(ADP-ribose) polymerase-1 (HsPARP-1) (anticancer target) |
| 3ua9 | −135.3 | Human tankyrase-2 (HsTANK2) = human poly(ADP-ribose) polymerase-5b (HsPARP-5b) (antitumor target) | |
| 4dbs | −124.0 | Human estrogenic 17β-hydroxysteroid dehydrogenase (17β-HSD1) | |
| −94.9 | Median docking energy | ||
| Eupha-2,8,22-triene-20,24S-diol b | 1uk1 | −137.6 | Human poly(ADP-ribose) polymerase-1 (HsPARP-1) (anticancer target) |
| 3ua9 | −135.8 | Human tankyrase-2 (HsTANK2) = human poly(ADP-ribose) polymerase-5b (HsPARP-5b) (antitumor target) | |
| 4dbs | −127.4 | Human estrogenic 17β-hydroxysteroid dehydrogenase (17β-HSD1) | |
| −85.1 | Median docking energy | ||
| Isofouquierol | 3lz6 | −134.1 | Cavia porcellus 11β-hydroxysteroid dehydrogenase type 1 (Cp11βHSD1) (diabetes target) |
| 2aba | −131.3 | Enterobacter cloacae pentaerythritol tetranitrate reductase (EcPETNR) (antibacterial target) | |
| 4kn0 | −130.4 | Human folate receptor β (HsFRβ) (anticancer target) | |
| −83.2 | Median docking energy | ||
| Isofouquieryl acetate | 3lz6 | −150.1 | Cavia porcellus 11β-hydroxysteroid dehydrogenase type 1 (Cp11βHSD1) (diabetes target) |
| 2aba | −137.9 | Enterobacter cloacae pentaerythritol tetranitrate reductase (EcPETNR) (antibacterial target) | |
| 4kn0 | −137.0 | Human folate receptor β (HsFRβ) (anticancer target) | |
| 1s0x | −135.7 | Human retinoic acid-related orphan receptor α (HsRORα) (may regulate lipid metabolism) | |
| 1h35 | −135.5 | Alicyclobacillus acidocardarius oxidosqualene cyclase (AaOSC) (cholesterol-lowering) | |
| 2w4q | −132.7 | Human zinc-binding alcohol dehydrogenase 1 (HsZADH1) | |
| 3g49 | −132.4 | Cavia porcellus 11β-hydroxysteroid dehydrogenase type 1 (Cp11βHSD1) (diabetes target) | |
| 2zxm | −131.0 | Rat vitamin D receptor (RnVDR) (target for psoriasis) | |
| −79.3 | Median docking energy | ||
| Lup-20(29)-ene-2α,3β-diol | 1xu9 | −112.5 | Human 11β-hydroxysteroid dehydrogenase type 1 (11βHSD1) (diabetes target) |
| 3wd2 | −112.4 | Serratia marcescens chitinase B (SmChiB) | |
| 1xl5 | −112.2 | HIV-1 protease | |
| −62.2 | Median docking energy | ||
| Neoilexonol | 1h22 | −118.2 | Torpedo californica acetylcholinesterase (TcAChE) (Alzheimer’s target) |
| −66.7 | Median docking energy | ||
| Neoilexonyl acetate | 2ilt | −118.8 | Human 11β-hydroxysteroid-dehydrogenase (Hs11β-HSDH) (diabetes target) |
| 4b84 | −118.5 | Murine acetylcholinesterase (MmAChE) (Alzheimer’s target) | |
| −74.8 | Median docking energy | ||
| Nizwanone | 4b84 | −123.2 | Murine acetylcholinesterase (MmAChE) (Alzheimer’s target) |
| −63.3 | Median docking energy | ||
| Ocotillyl acetate | 1gsz | −144.6 | Alicyclobacillus acidocardarius oxidosqualene cyclase (AaOSC) (cholesterol-lowering) |
| 2aba | −130.1 | Enterobacter cloacae pentaerythritol tetranitrate reductase (EcPETNR) (antibacterial target) | |
| 4jbs | −127.7 | Human endoplasmic reticulum aminopeptidase 2 (HsERAP2) (immune response target) | |
| −70.7 | Median docking energy | ||
| Olibanumol E | 2ilt | −109.5 | Human 11β-hydroxysteroid-dehydrogenase (Hs11β-HSDH) (diabetes target) |
| −63.5 | Median docking energy | ||
| Olibanumol F | 1cgl | −110.1 | Human fibroblast collagenase (HsMMP-1) (arthritis target) |
| 3bt9 | −108.8 | Staphylococcus aureus multidrug binding protein (SaQacR) | |
| −36.7 | Median docking energy | ||
| Olibanumol G | 3bt9 | −117.4 | Staphylococcus aureus multidrug binding protein (SaQacR) |
| −53.0 | Median docking energy | ||
| Olibanumol H | 3bt9 | −121.6 | Staphylococcus aureus multidrug binding protein (SaQacR) |
| −60.9 | Median docking energy | ||
| Olibanumol I | 3bt9 | −115.7 | Staphylococcus aureus multidrug binding protein (SaQacR) |
| −63.2 | Median docking energy | ||
| Olibanumol J | 1rpw | −135.5 | Staphylococcus aureus multidrug binding protein (SaQacR) |
| 1cr6 | −133.3 | Murine soluble epoxide hydrolase (MmEPHX2) (anti-inflammatory target) | |
| 3bti | −132.6 | Staphylococcus aureus multidrug binding protein (SaQacR) | |
| 3gyt | −132.1 | Strongyloides stercoralis nuclear receptor DAF-12 (SsDAF12) (antiparasitic target) | |
| −55.5 | Median docking energy | ||
| Olibanumol J’ | 3l3m | −132.3 | Human poly(ADP-ribose) polymerase-1 (HsPARP-1) (anticancer target) |
| 3g49 | −129.0 | Cavia porcellus 11β-hydroxysteroid dehydrogenase type 1 (Cp11βHSD1) (diabetes target) | |
| 3w5e | −128.8 | Human phosphodiesterase 4B (HsPDE4B) (anti-inflammatory target) | |
| 3bt9 | −123.4 | Staphylococcus aureus multidrug binding protein (SaQacR) | |
| −74.7 | Median docking energy | ||
| Olibanumol K | 4b84 | −122.9 | Murine acetylcholinesterase (MmAChE) (Alzheimer’s target) |
| 3i6m | −122.1 | Torpedo californica acetylcholinesterase (TcAChE) (Alzheimer’s target) | |
| −64.8 | Median docking energy | ||
| Olibanumol L | 1ry0 | −116.2 | Human prostaglandin F synthase (HsPGFS) (hypertension target) |
| 3g49 | −111.1 | Cavia porcellus 11β-hydroxysteroid dehydrogenase type 1 (Cp11βHSD1) (diabetes target) | |
| −27.6 | Median docking energy | ||
| Olibanumol L’ | 4b84 | −130.1 | Murine acetylcholinesterase (MmAChE) (Alzheimer’s target) |
| −70.7 | Median docking energy | ||
| Olibanumol M | 1h22 | −128.4 | Torpedo californica acetylcholinesterase (TcAChE) (Alzheimer’s target) |
| −62.7 | Median docking energy | ||
| Olibanumol N | 4b84 | −133.0 | Murine acetylcholinesterase (MmAChE) (Alzheimer’s target) |
| −70.9 | Median docking energy | ||
| Trametenolic acid B | 3ua9 | −131.6 | Human tankyrase-2 (HsTANK2) = human poly(ADP-ribose) polymerase-5b (HsPARP-5b) (antitumor target) |
| 3g49 | −127.1 | Cavia porcellus 11β-hydroxysteroid dehydrogenase type 1 (11βHSD1) (diabetes target) | |
| 3hfb | −126.6 | Human tryoptophan hydroxylase type 1 (HsTPH1) (biosynthesis of serotonin) | |
| 1eve | −126.5 | Torpedo californica acetylcholinesterase (TcAChE) (Alzheimer’s target) | |
| −74.2 | Median docking energy | ||
| Urs-12-ene-3α,11α-diol | 1h22 | −120.9 | Torpedo californica acetylcholinesterase (TcAChE) (Alzheimer’s target) |
| −61.1 | Median docking energy | ||
| Urs-12-ene-3β,11α-diol | 4b84 | −123.7 | Murine acetylcholinesterase (MmAChE) (Alzheimer’s target) |
| −59.8 | Median docking energy | ||
| α-Boswellic acid | 3g49 | −110.0 | Cavia porcellus 11β-hydroxysteroid dehydrogenase type 1 (11βHSD1) (diabetes target) |
| −65.7 | Median docking energy | ||
| α-Elemolic acid | 3ua9 | −152.7 | Human tankyrase-2 (HsTANK2) = human poly(ADP-ribose) polymerase-5b (HsPARP-5b) (antitumor target) |
| 1c3s | −136.9 | Aquifex aeolicus histone deacetylase (AaHDAC) (anticancer target) | |
| 3g49 | −128.9 | Cavia porcellus 11β-hydroxysteroid dehydrogenase type 1 (11βHSD1) (diabetes target) | |
| −87.8 | Median docking energy | ||
| β-Boswellic acid | 3i6m | −118.1 | Torpedo californica acetylcholinesterase (TcAChE) (Alzheimer’s target) |
| +20.7 | Median docking energy | ||
| β-Elemonic acid | 3ua9 | −147.3 | Human tankyrase-2 (HsTANK2) = human poly(ADP-ribose) polymerase-5b (HsPARP-5b) (antitumor target) |
| 3l3m | −133.7 | Human poly(ADP-ribose) polymerase-1 (HsPARP-1) (anticancer target) | |
| 3g49 | −131.9 | Cavia porcellus 11β-hydroxysteroid dehydrogenase type 1 (11βHSD1) (diabetes target) | |
| −89.0 | Median docking energy | ||
| δ-Boswellic acid | 3g49 | −106.7 | Cavia porcellus 11β-hydroxysteroid dehydrogenase type 1 (11βHSD1) (diabetes target) |
| −60.7 | Median docking energy |
| Ligand | MmEPHX2 | HsEPHX2 | HsMMP-1 | SsPLA2 | HsPLA2 | Hs5-LOX | Mm iNOS | HsPI3Kγ | HsIRAK4 | HsERAP2 | HsGSTO1 | HsPDE4B |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1cr6 | 4hai | 1cgl | 2b03 | 1j1a | 3v99 | 1m8d | 2a5u | 5t1s | 4jbs | 5v3q | 3w5e | |
| (20S)-3,7-Dioxotirucalla-8,24-dien-21-oic acid | −113.3 | −114.1 | −121.8 | −127.4 | −106.9 | −103.1 | −88.0 | −87.7 | −112.7 | −107.3 | −106.0 | −114.0 |
| 11-Ethoxy-β-boswellic acid | −80.3 | −96.3 | −107.9 | −100.9 | −87.3 | −85.8 | −71.7 | −82.5 | −99.7 | −104.5 | −66.8 | −101.5 |
| 11-Keto-β-boswellic acid | −75.7 | −91.8 | −100.7 | −94.9 | −85.3 | −83.3 | −64.2 | −77.2 | −89.7 | −99.4 | −70.7 | 2.8 |
| 12-Ursen-3,11-dione | −73.9 | −88.8 | −102.0 | −102.4 | −71.2 | −75.8 | −63.0 | −74.9 | −82.0 | −98.6 | −71.4 | −99.9 |
| 12-Ursen-3,24-diol | −81.9 | −84.5 | −105.5 | −88.7 | −93.7 | −77.4 | −60.4 | −76.9 | −89.8 | −96.4 | −87.8 | −101.1 |
| 2,3-Dihydroxy-12-ursen-24-oic acid | −78.8 | −83.9 | −98.9 | −100.0 | −91.1 | −81.7 | −30.0 | −78.6 | −91.7 | −96.5 | −74.8 | −102.4 |
| 20,22-Epoxytirucall-24-en-3-one | −98.2 | −102.5 | −113.1 | −111.0 | −95.7 | −90.2 | −89.9 | −82.5 | −94.7 | −105.1 | −80.9 | −99.3 |
| 24-Nor-3,12-ursadien-11-one | −84.1 | −89.6 | −99.9 | −103.4 | −95.6 | −77.5 | −71.8 | −75.1 | −64.1 | −91.7 | −50.1 | −83.4 |
| 24-Nor-3,9(11),12-oleanatriene | −83.4 | −94.7 | −99.5 | −99.9 | −93.3 | −70.8 | −61.0 | −75.2 | −59.9 | −95.3 | −47.3 | −90.8 |
| 24-Nor-3,9(11),12-ursatriene | −83.3 | −89.4 | −100.6 | −99.3 | −85.5 | −81.4 | −65.0 | −75.3 | −77.0 | −86.3 | −62.2 | −91.2 |
| 3-Acetoxy-12,20(29)-lupadien-24-oic acid | −77.8 | −85.2 | −115.4 | −87.5 | −82.7 | −83.7 | −68.4 | −93.6 | −59.1 | −105.0 | −78.9 | −97.8 |
| 3-Acetoxy-20(29)-lupen-24-oic acid (=3-Acetyl lupeolic acid) | −84.1 | −83.5 | −117.5 | −86.7 | −87.1 | −82.7 | −71.8 | −92.9 | −79.7 | −104.5 | −79.0 | −92.8 |
| 3β-Acetoxy-20S,24R-dihydroxydammar-25-ene | −109.7 | −109.3 | −124.7 | −118.5 | −120.3 | −100.8 | −96.7 | −94.4 | −106.4 | −119.1 | −85.8 | −116.3 |
| 3β-Acetoxy-20S,24S-dihydroxydammar-25-ene | −106.2 | −109.2 | −121.7 | −115.6 | −122.0 | −100.3 | −81.5 | −93.9 | −109.5 | −126.1 | −76.5 | −111.0 |
| 3-Acetoxy-5,12-ursadien-24-oic acid | −92.6 | −78.1 | −121.1 | −97.5 | −81.7 | −81.0 | −73.3 | −83.6 | −70.4 | −105.0 | −70.2 | −47.3 |
| 3-Acetyl-11-keto-β-boswellic acid | −87.3 | −75.6 | −107.6 | −102.4 | −74.0 | −84.9 | −67.7 | −79.7 | −71.9 | −93.3 | −71.6 | −49.2 |
| 3-Acetyl-11α-methoxy-β-boswellic_acid | −86.7 | −91.1 | −109.3 | −91.0 | −75.4 | −87.3 | −76.1 | −79.5 | −90.0 | −101.9 | −69.5 | −52.0 |
| 3-Acetyl-9,11-dehydro-β-boswellic_acid | −90.4 | −80.3 | −120.6 | −98.5 | −94.8 | −82.7 | −74.4 | −82.7 | −61.1 | −103.7 | −84.3 | −50.3 |
| 3-Acetyl-α-boswellic acid | −79.0 | −82.4 | −109.7 | −112.7 | −97.2 | −82.0 | −70.5 | −83.0 | −83.8 | −99.7 | −72.7 | −91.4 |
| 3-Acetyl-β-boswellic acid | −91.5 | −83.1 | −119.8 | −99.4 | −96.2 | −85.1 | −69.0 | −79.4 | −70.8 | −90.2 | −80.6 | −91.8 |
| 3-Hydroxy-20(29)-lupen-24-oic acid (=Lupeolic acid) | −103.4 | −105.0 | −106.6 | −82.6 | −76.6 | −79.2 | −72.6 | −81.8 | −76.0 | −103.4 | −69.5 | −87.3 |
| 3-Oxotirucalla-7,9(11),24-trien-21-oic acid | −113.1 | −113.3 | −123.3 | −116.9 | −113.5 | −100.5 | −86.3 | −92.1 | −103.6 | −108.8 | −99.8 | −113.2 |
| 3β-Hydroxytirucalla-8,24-dien-21-oic acid | −110.8 | −112.1 | −118.6 | −123.4 | −106.5 | −103.3 | −97.3 | −80.7 | −107.8 | −109.4 | −94.6 | −111.9 |
| 3α-Acetoxytirucalla-7,24-dien-21-oic acid | −115.5 | −113.3 | −129.9 | −109.8 | −116.2 | −91.2 | −95.4 | −95.3 | −109.7 | −106.1 | −91.4 | −104.9 |
| 3α-Hydroxytirucalla-7,24-dien-21-oic acid | −110.8 | −115.7 | −115.4 | −117.2 | −110.1 | −105.7 | −84.1 | −86.9 | −102.8 | −108.6 | −82.9 | −109.8 |
| 3β-Acetoxydammar-24-ene-16β,20R-diol | −102.8 | −110.3 | −125.7 | −111.2 | −109.3 | −97.7 | −97.0 | −90.7 | −107.9 | −125.2 | −79.8 | −115.2 |
| 3β-Acetoxylup-20(29)-en-11β-ol | −94.6 | −92.9 | −109.8 | −105.6 | −90.8 | −86.1 | −87.1 | −80.9 | −93.2 | −108.2 | −74.5 | −41.2 |
| 4,23-Dihydroburic acid | −101.2 | −101.6 | −109.8 | −120.9 | −92.0 | −88.3 | −72.0 | −84.3 | −102.9 | −110.2 | −85.3 | −107.7 |
| 6,7-Epoxy-9(11)-oleanen-3-ol | −53.0 | −111.6 | −100.2 | −106.3 | −89.8 | −79.6 | −64.5 | −67.7 | −66.2 | −94.9 | −68.6 | −87.2 |
| 6,7-Epoxy-9(11)-oleanen-3-one | −66.0 | −100.9 | −105.0 | −107.8 | −93.3 | −75.1 | −68.5 | −70.3 | −40.3 | −100.6 | −66.1 | −106.2 |
| 9,11-Dehydro-β-boswellic acid | −75.8 | −85.7 | −109.8 | −96.8 | −93.1 | −79.8 | −27.4 | −73.6 | −89.5 | −91.2 | −83.4 | −62.6 |
| Boscartene L | −83.2 | −110.2 | −104.4 | −115.7 | −77.4 | −100.1 | −81.6 | −82.0 | −100.0 | −108.6 | −62.6 | −97.7 |
| Boscartene M | −73.8 | −110.8 | −114.0 | −107.0 | −96.7 | −102.9 | −86.8 | −83.4 | −76.8 | −110.4 | −83.5 | −91.5 |
| Boscartene N | −110.2 | −113.7 | −116.6 | −126.2 | −106.6 | −92.9 | −85.8 | −87.4 | −109.6 | −111.8 | −100.1 | −118.6 |
| Dammarenediol II | −114.1 | −112.3 | −126.2 | −103.7 | −110.8 | −95.8 | −77.3 | −87.2 | −110.3 | −108.4 | −101.4 | −120.5 |
| Dammarenediol II acetate | −109.5 | −109.5 | −113.4 | −101.8 | −115.7 | −105.2 | −92.1 | −97.4 | −115.2 | −107.2 | −77.7 | −104.9 |
| Eupha-2,8,22-triene-20,24R-diol | −111.9 | −106.1 | −112.9 | −121.3 | −98.2 | −97.9 | −80.5 | −85.9 | −101.0 | −106.4 | −92.8 | −104.5 |
| Eupha-2,8,22-triene-20,24S-diol | −117.2 | −104.4 | −113.1 | −115.5 | −100.1 | −97.8 | −82.6 | −87.3 | −99.4 | −103.0 | −94.1 | −105.8 |
| Isofouquierol | −116.4 | −103.7 | −118.4 | −113.5 | −114.9 | −104.0 | −96.4 | −93.2 | −110.4 | −114.1 | −96.8 | −128.3 |
| Isofouquieryl acetate | −107.6 | −119.6 | −123.5 | −122.4 | −115.8 | −96.8 | −103.7 | −86.4 | −113.4 | −124.4 | −85.2 | −106.6 |
| Lup-20(29)-ene-2α,3β-diol | −101.4 | −98.8 | −106.0 | −95.5 | −83.0 | −71.4 | −87.9 | −75.4 | −89.8 | −107.6 | −73.2 | −73.6 |
| Neoilexonol | −73.0 | −87.9 | −101.1 | −104.8 | −71.4 | −74.7 | −62.8 | −69.5 | −52.3 | −92.8 | −73.9 | −98.0 |
| Neoilexonyl acetate | −86.0 | −86.7 | −108.2 | −109.9 | −83.8 | −79.4 | −68.2 | −74.3 | −87.5 | −95.5 | −69.4 | −31.7 |
| Nizwanone | −81.2 | −89.7 | −108.6 | −89.5 | −95.3 | −78.5 | −60.5 | −70.1 | −88.3 | −98.1 | −90.9 | −106.3 |
| Ocotillyl acetate | −94.4 | −96.1 | −121.5 | −105.5 | −107.2 | −96.7 | −72.8 | −84.5 | −108.2 | −127.7 | −77.5 | −97.2 |
| Olibanumol E | −81.7 | −82.4 | −101.7 | −98.7 | −91.8 | −82.2 | −60.6 | −79.4 | −92.4 | −97.9 | −49.2 | −99.2 |
| Olibanumol F | −85.7 | −108.9 | −110.1 | −86.1 | −79.7 | −87.6 | −65.9 | −86.4 | −80.4 | −95.0 | −77.3 | −46.1 |
| Olibanumol G | −81.5 | −95.3 | −99.3 | −82.4 | −70.4 | −80.0 | −61.0 | −77.9 | −80.8 | −93.2 | −54.0 | −72.7 |
| Olibanumol H | −101.1 | −113.9 | −107.7 | −102.3 | −86.8 | −82.6 | −78.1 | −88.4 | −94.2 | −103.8 | −74.1 | −55.0 |
| Olibanumol I | −96.2 | −97.4 | −101.5 | −93.4 | −70.6 | −81.9 | −75.5 | −78.2 | −88.8 | −96.9 | −71.6 | −79.2 |
| Olibanumol J | −133.3 | −119.3 | −114.2 | −112.5 | −117.6 | −106.1 | −93.8 | −88.4 | −108.3 | −123.8 | −99.3 | −112.3 |
| Olibanumol J’ | −108.2 | −119.2 | −112.7 | −112.3 | −118.8 | −107.7 | −84.4 | −84.3 | −109.2 | −124.4 | −84.6 | −128.8 |
| Olibanumol K | −88.8 | −77.8 | −110.3 | −90.5 | −94.9 | −77.0 | −64.9 | −79.8 | −76.2 | −89.9 | −78.8 | −82.4 |
| Olibanumol L | −78.1 | −80.8 | −109.6 | −91.7 | −98.1 | −81.3 | −67.6 | −73.8 | −82.4 | −97.3 | −64.1 | −108.1 |
| Olibanumol L’ | −85.0 | −74.1 | −111.4 | −97.1 | −83.9 | −84.7 | −67.6 | −77.4 | −72.7 | −92.4 | −70.0 | −37.9 |
| Olibanumol M | −77.5 | −89.2 | −100.1 | −95.5 | −57.5 | −83.9 | −69.2 | −77.1 | −96.5 | −93.1 | −63.9 | −100.0 |
| Olibanumol N | −85.0 | −76.5 | −103.3 | −84.5 | −70.8 | −86.3 | −73.6 | −79.7 | −87.9 | −96.8 | −70.4 | −50.1 |
| Trametenolic acid B | −106.3 | −107.3 | −106.9 | −107.3 | −107.9 | −99.7 | −92.6 | −84.0 | −107.6 | −115.0 | −92.9 | −111.6 |
| Urs-12-ene-3α,11α-diol | −77.8 | −78.6 | −104.3 | −94.2 | −85.2 | −84.4 | −68.3 | −77.7 | −92.2 | −90.9 | −64.6 | −100.1 |
| Urs-12-ene-3β,11α-diol | −79.4 | −74.8 | −101.3 | −108.0 | −71.2 | −76.9 | −67.3 | −69.9 | −70.9 | −93.8 | −61.8 | −114.1 |
| α-Boswellic acid | −77.9 | −87.8 | −103.7 | −100.2 | −95.0 | −80.7 | −64.4 | −79.4 | −85.7 | −94.8 | −62.5 | −62.0 |
| α-Elemolic acid | −106.5 | −110.5 | −113.9 | −124.4 | −104.1 | −99.7 | −107.1 | −84.0 | −108.7 | −111.1 | −97.3 | −110.3 |
| β-Boswellic acid | −78.0 | −87.8 | −108.9 | −94.2 | −90.0 | −82.5 | −7.8 | −76.7 | −91.1 | −94.4 | −85.2 | −95.5 |
| β-Elemonic acid | −112.2 | −113.8 | −110.5 | −123.3 | −109.6 | −100.4 | −92.4 | −86.3 | −106.6 | −107.1 | −108.0 | −112.6 |
| β-Elemonic acid | −110.0 | −114.6 | −120.2 | −120.4 | −106.3 | −99.8 | −98.8 | −83.5 | −109.2 | −109.0 | −103.6 | −116.2 |
| δ-Boswellic acid | −76.4 | −90.4 | −101.3 | −89.6 | −82.5 | −74.7 | −69.2 | −80.7 | −87.2 | −96.4 | −64.4 | −76.4 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Byler, K.G.; Setzer, W.N. Protein Targets of Frankincense: A Reverse Docking Analysis of Terpenoids from Boswellia Oleo-Gum Resins. Medicines 2018, 5, 96. https://doi.org/10.3390/medicines5030096
Byler KG, Setzer WN. Protein Targets of Frankincense: A Reverse Docking Analysis of Terpenoids from Boswellia Oleo-Gum Resins. Medicines. 2018; 5(3):96. https://doi.org/10.3390/medicines5030096
Chicago/Turabian StyleByler, Kendall G., and William N. Setzer. 2018. "Protein Targets of Frankincense: A Reverse Docking Analysis of Terpenoids from Boswellia Oleo-Gum Resins" Medicines 5, no. 3: 96. https://doi.org/10.3390/medicines5030096
APA StyleByler, K. G., & Setzer, W. N. (2018). Protein Targets of Frankincense: A Reverse Docking Analysis of Terpenoids from Boswellia Oleo-Gum Resins. Medicines, 5(3), 96. https://doi.org/10.3390/medicines5030096