

A Two-Compartment Fermentation System to Quantify Strain-Specific Interactions in Microbial Co-Cultures

 , , ,
, , , 
(This article belongs to the Section Biochemical Engineering)
Abstract
:1. Introduction
- Computational approaches are being steadily extended to unravel and predict interactions between bacteria [21,22,23]. To improve the simulation results, data from quantitative experiments providing strain-specific information—in particular, strain-specific growth rates, metabolite production, and consumption rates—are essential to validate model qualities, as indicated previously [24,25,26,27,28].
- Adaptive evolution experiments have been used to improve the performance of strains [31,32] and have been adapted for co-culture systems [33,34,35]. However, to select them for the jointly increased growth of co-cultures, individual adjustments may be necessary, such as the implementation of individual dilution rates to prevent overgrowth and washout scenarios.
2. Materials and Methods
2.1. Medium Conditions
2.2. Strain Cultivation
2.3. Biomass Quantification via the Optical Density Method
2.4. Biomass Quantification via Flow Cytometry
2.5. Membrane Unit
2.6. Vessel Bioreactor System
2.7. Tube Bioreactor System
2.8. Continuous Cultivation in the Tube Bioreactor System
2.9. Metabolite Balancing
2.10. Reaction Rate Constant of Metabolite Productivity
2.11. Determination of Amino Acid Transport Coefficients in the Membrane Unit
2.12. Determination of the Bodenstein Number
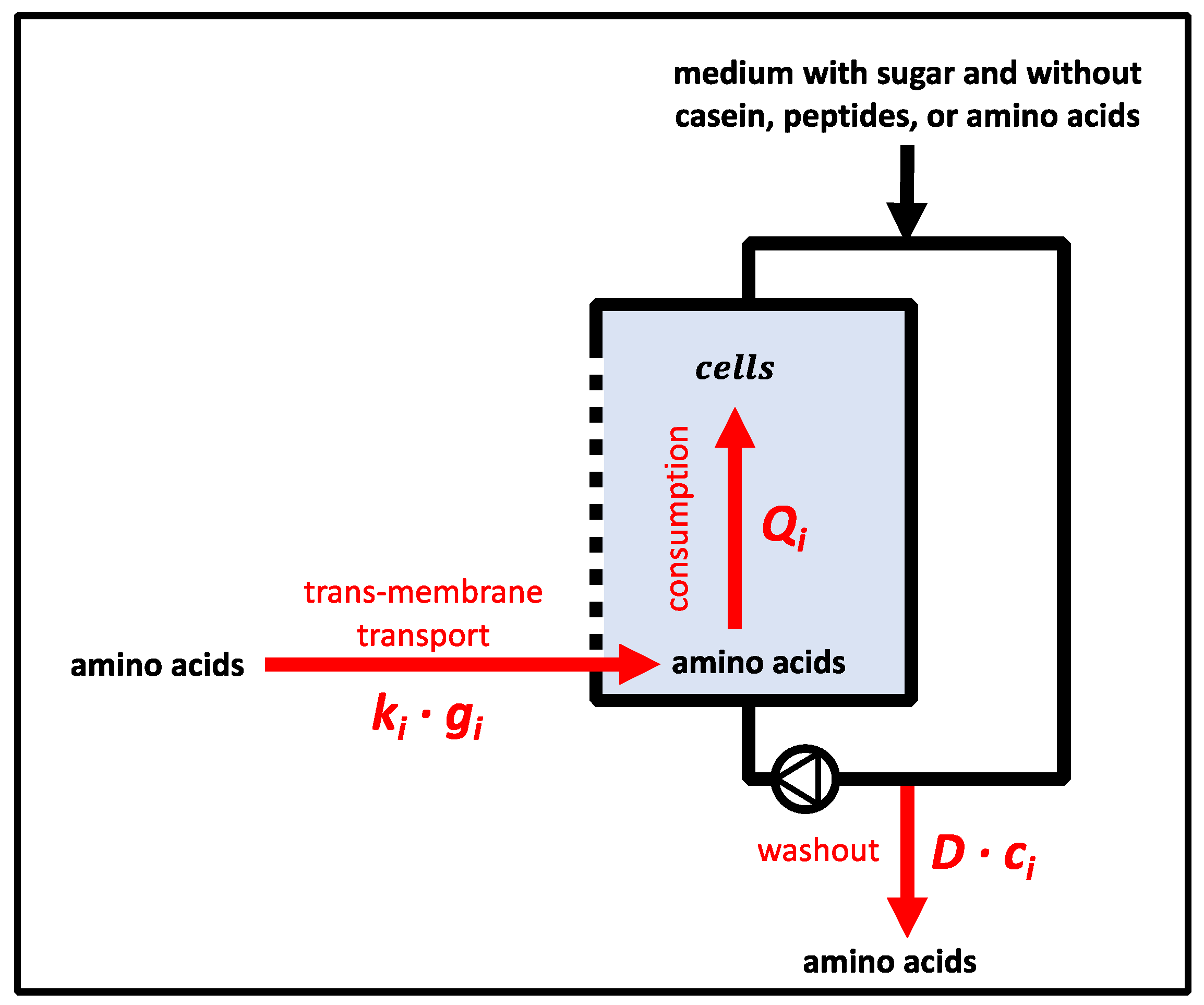
2.13. Calculation of the Damkoehler Number
2.14. Quantification of Extracellular Metabolites
2.15. Quantification of Extracellular and Intracellular Metabolites
2.16. Determination of Amino Acid Composition in Casein
2.17. Uncertainty Analysis
3. Results
3.1. Design of the Membrane Unit
3.1.1. Membrane Unit Characteristics
3.1.2. Amino Acid Transport in the Membrane Unit
3.2. Design of the 2cs
3.3. Vessel Bioreactor System: Set-Up and Growth Experiment
3.4. Tube Bioreactor System
3.5. Comparison between Bacterial Growth in Serum Bottles and in the Tube Bioreactor System
3.6. Determination of Strain-Specific Rates in Co-Culture
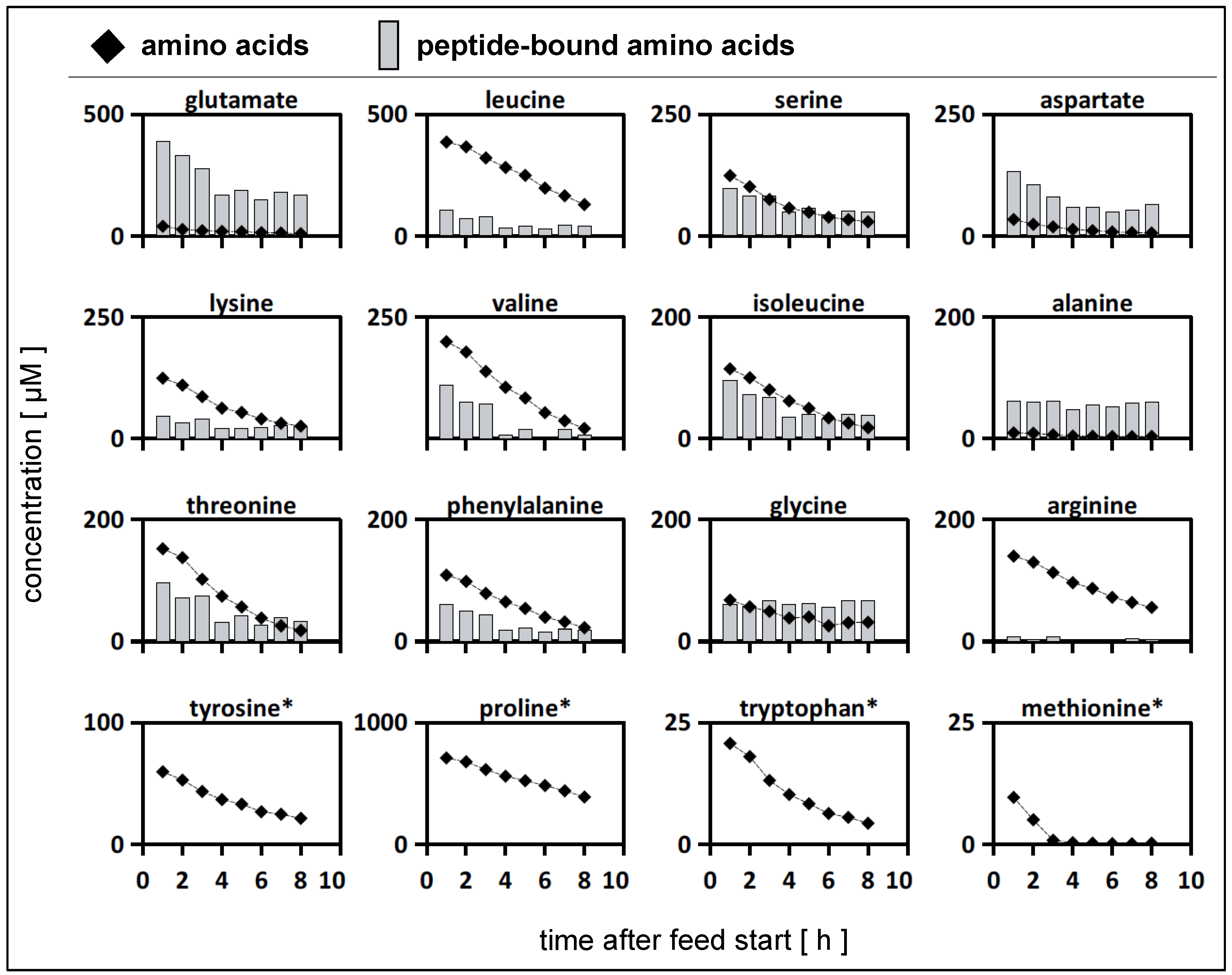
3.6.1. Dynamic Cultivation Tests in the Tube Bioreactor System
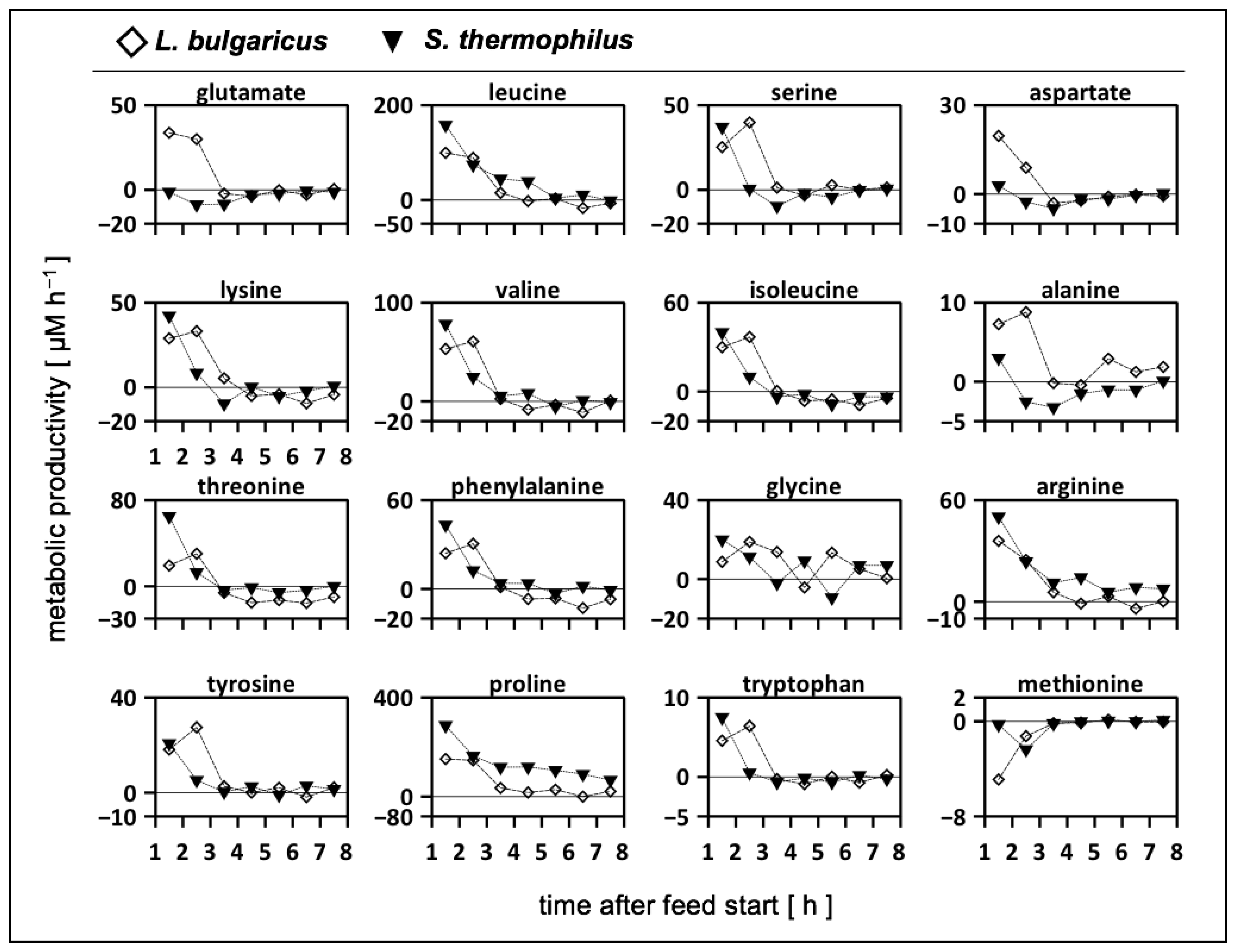
3.6.2. Calculation of Strain-Specific Rates
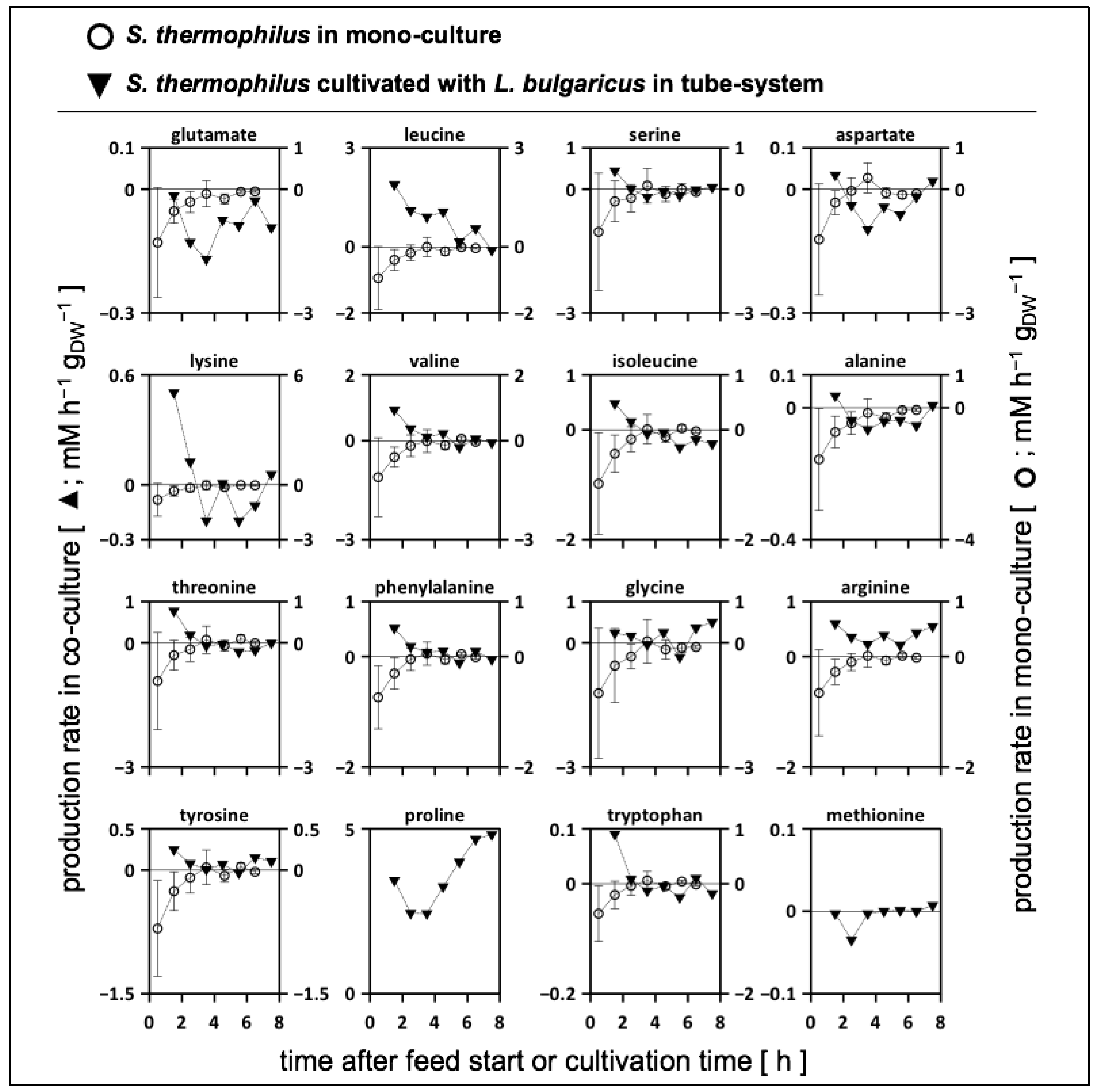
3.6.3. Biomass-Specific Activity of S. thermophilus in Mono- and Co-Cultures
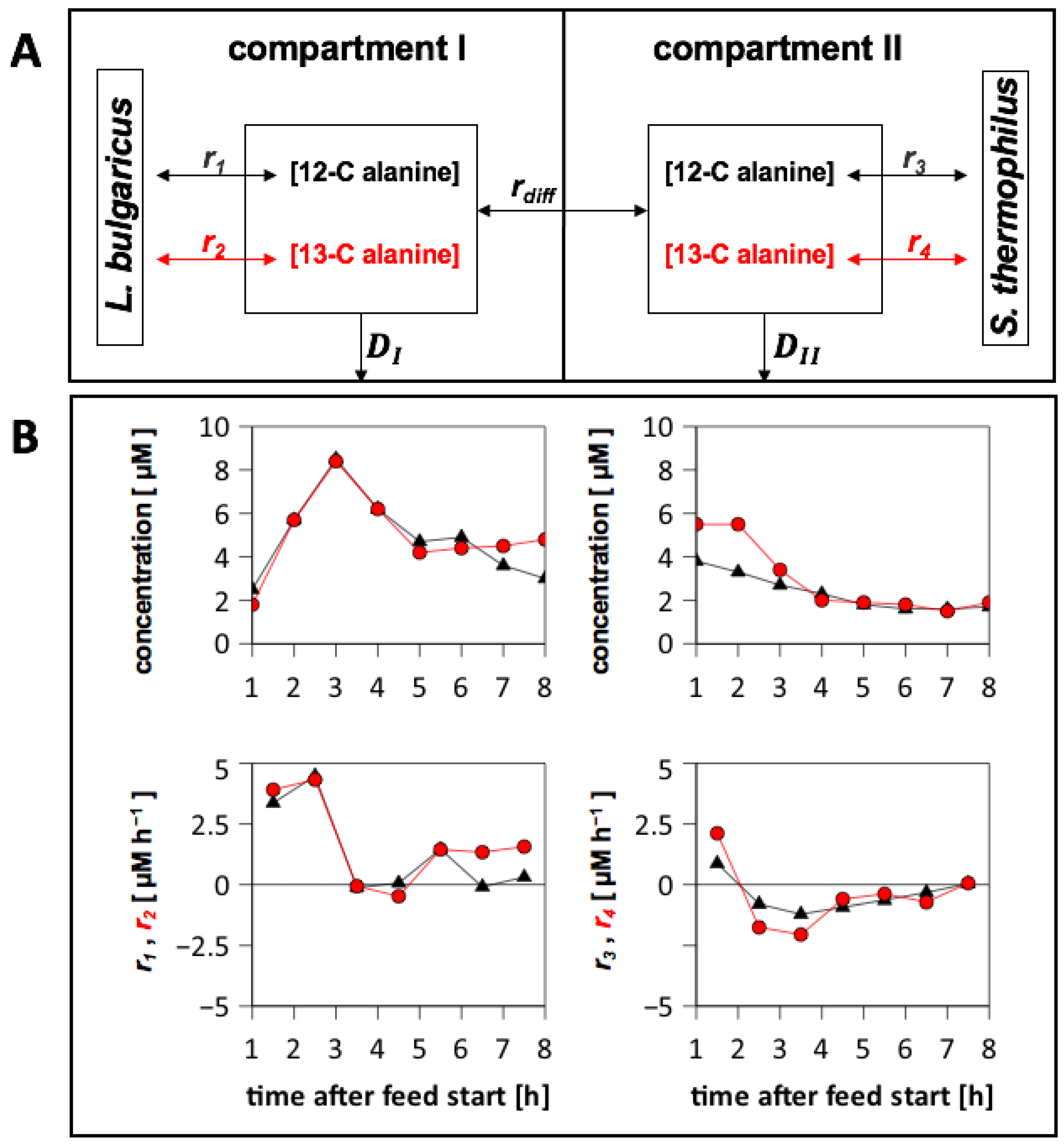
3.6.4. Analysis of Extracellular 13-C Alanine Enrichment
3.6.5. Alanine Exchange between the Compartments
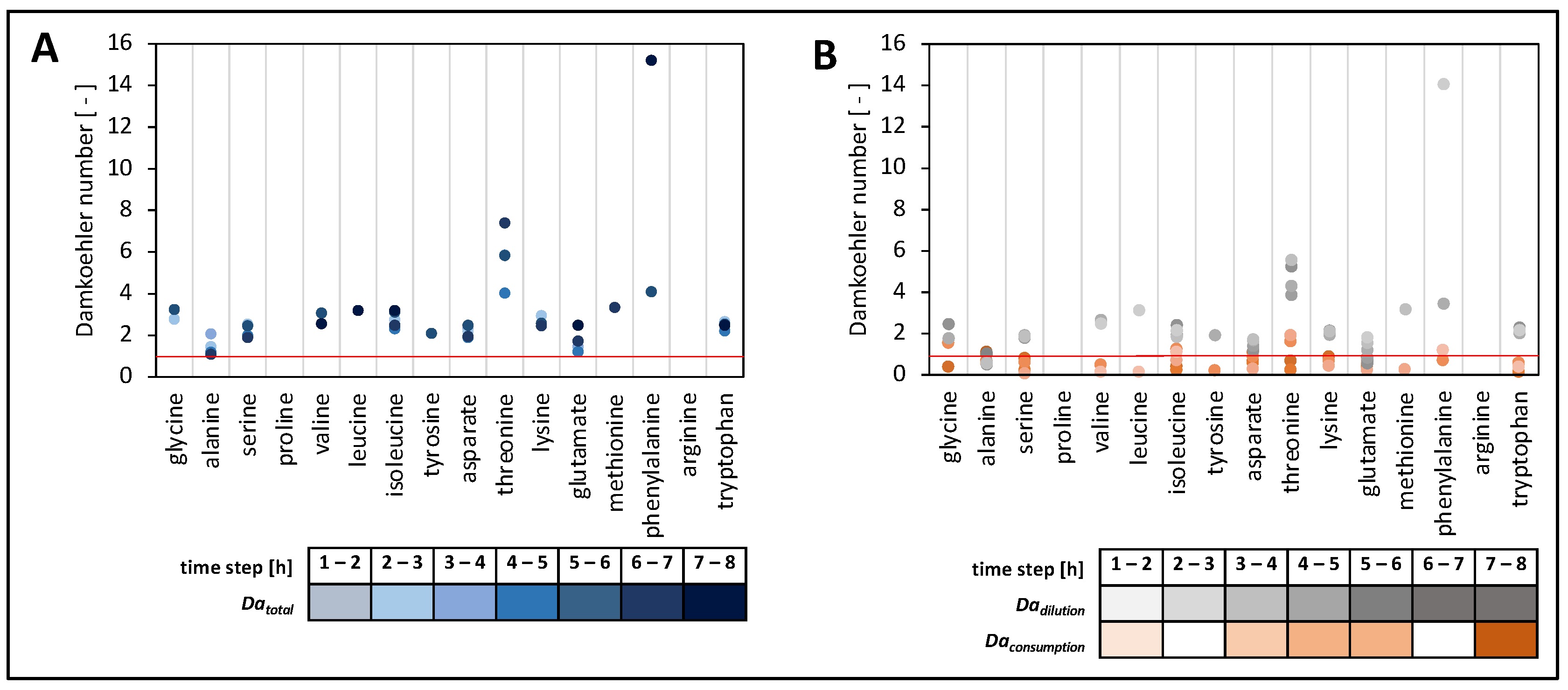
3.6.6. Calculation of Damkoehler Numbers
4. Discussion
4.1. Process Characterisation
4.2. Difference between Cultivation in the Serum Bottle and in the Tube Bioreactor System
4.3. Strain-Specific Amino Acid Release and Consumption in the Tube Bioreactor System
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- West, S.A.; Diggle, S.P.; Buckling, A.; Gardner, A.; Griffin, A.S. The Social Lives of Microbes. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 53–77. [Google Scholar] [CrossRef] [Green Version]
- Lyons, N.A.; Kolter, R. On the Evolution of Bacterial Multicellularity. Curr. Opin. Microbiol. 2015, 24, 21–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Souza, G.; Shitut, S.; Preussger, D.; Yousif, G.; Waschina, S.; Kost, C. Ecology and Evolution of Metabolic Cross-Feeding Interactions in Bacteria. Nat. Prod. Rep. 2018, 35, 455–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bassler, B.L. Small Talk: Cell-to-Cell Communication in Bacteria. Cell 2002, 109, 421–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziesack, M.; Gibson, T.; Oliver, J.K.W.; Shumaker, A.M.; Hsu, B.B.; Riglar, D.T.; Giessen, T.W.; Di Benedetto, N.V.; Bry, L.; Way, J.C.; et al. Engineered Interspecies Amino Acid Cross-Feeding Increases Population Evenness in a Synthetic Bacterial Consortium. mSystems 2019, 4, e00352-19. [Google Scholar] [CrossRef] [Green Version]
- Hennig, S.; Rödel, G.; Ostermann, K. Artificial Cell-Cell Communication as an Emerging Tool in Synthetic Biology Applications. J. Biol. Eng. 2015, 9, 13. [Google Scholar] [CrossRef] [Green Version]
- Giri, S.; Shitut, S.; Kost, C. Harnessing Ecological and Evolutionary Principles to Guide the Design of Microbial Production Consortia. Curr. Opin. Biotechnol. 2020, 62, 228–238. [Google Scholar] [CrossRef]
- Kleerebezem, R.; van Loosdrecht, M.C. Mixed Culture Biotechnology for Bioenergy Production. Curr. Opin. Biotechnol. 2007, 18, 207–212. [Google Scholar] [CrossRef]
- Wintermute, E.H.; Silver, P.A. Emergent Cooperation in Microbial Metabolism. Mol. Syst. Biol. 2010, 6, 407. [Google Scholar] [CrossRef]
- Brenner, K.; You, L.; Arnold, F.H. Engineering Microbial Consortia: A New Frontier in Synthetic Biology. Trends Biotechnol. 2008, 26, 483–489. [Google Scholar] [CrossRef]
- Shong, J.; Jimenez Diaz, M.R.; Collins, C.H. Towards Synthetic Microbial Consortia for Bioprocessing. Curr. Opin. Biotechnol. 2012, 23, 798–802. [Google Scholar] [CrossRef] [PubMed]
- McCarty, N.S.; Ledesma-Amaro, R. Synthetic Biology Tools to Engineer Microbial Communities for Biotechnology. Trends Biotechnol. 2019, 37, 181–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noack, S.; Baumgart, M. Communities of Niche-Optimized Strains: Small-Genome Organism Consortia in Bioproduction. Trends Biotechnol. 2018, 37, 126–139. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.A.; Vernacchio, V.R.; Sinkoe, A.L.; Collins, S.M.; Ibrahim, M.H.A.; Lachance, D.M.; Hahn, J.; Koffas, M.A.G. Experimental and Computational Optimization of an Escherichia Coli Co-Culture for the Efficient Production of Flavonoids. Metab. Eng. 2016, 35, 55–63. [Google Scholar] [CrossRef]
- Zhou, K.; Qiao, K.; Edgar, S.; Stephanopoulos, G. Distributing a Metabolic Pathway among a Microbial Consortium Enhances Production of Natural Products. Nat. Biotechnol. 2015, 33, 377–383. [Google Scholar] [CrossRef] [Green Version]
- Pande, S.; Merker, H.; Bohl, K.; Reichelt, M.; Schuster, S.; De Figueiredo, L.F.; Kaleta, C.; Kost, C. Fitness and Stability of Obligate Cross-Feeding Interactions That Emerge upon Gene Loss in Bacteria. ISME J. 2014, 8, 953–962. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Wang, X. Modular Co-Culture Engineering, a New Approach for Metabolic Engineering. Metab. Eng. 2016, 37, 114–121. [Google Scholar] [CrossRef]
- Tang, Y.J.; Fong, S.S.; Koffas, M.A.; Carr, R.R.; Zha, J.; Roell, G.W. Engineering Microbial Consortia by Division of Labor. Microb. Cell Fact. 2019, 18, 35. [Google Scholar] [CrossRef] [Green Version]
- Giri, S.; Waschina, S.; Kaleta, C.; Kost, C. Defining Division of Labour in Microbial Communities. J. Mol. Biol. 2019, 431, 4712–4731. [Google Scholar] [CrossRef]
- Thommes, M.; Wang, T.; Zhao, Q.; Paschalidis, I.C.; Segrè, D. Designing Metabolic Division of Labor in Microbial Communities. mSystems 2019, 4, e00263-18. [Google Scholar] [CrossRef]
- Xu, P. Dynamics of Microbial Competition, Commensalism, and Cooperation and Its Implications for Coculture and Microbiome Engineering. Biotechnol. Bioeng. 2020, 118, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Germerodt, S.; Bohl, K.; Lück, A.; Pande, S.; Schröter, A.; Kaleta, C.; Schuster, S.; Kost, C. Pervasive Selection for Cooperative Cross-Feeding in Bacterial Communities. PLoS Comput. Biol. 2016, 12, e1004986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lahmann, P.; Reichl, U.; Klamt, S.; Kohrs, F.; Bissinger, T.; Wendschuh, S.; Koch, S.; Benndorf, D. RedCom: A Strategy for Reduced Metabolic Modeling of Complex Microbial Communities and Its Application for Analyzing Experimental Datasets from Anaerobic Digestion. PLoS Comput. Biol. 2019, 15, e00263-18. [Google Scholar] [CrossRef] [Green Version]
- Zengler, K.; Palsson, B.O. A Road Map for the Development of Community Systems (CoSy) Biology. Nat. Rev. Microbiol. 2012, 10, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Branco dos Santos, F.; de Vos, W.M.; Teusink, B. Towards Metagenome-Scale Models for Industrial Applications-the Case of Lactic Acid Bacteria. Curr. Opin. Biotechnol. 2013, 24, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Hanemaaijer, M.; Röling, W.F.M.; Olivier, B.G.; Khandelwal, R.A.; Teusink, B.; Bruggeman, F.J. Systems Modeling Approaches for Microbial Community Studies: From Metagenomics to Inference of the Community Structure. Front. Microbiol. 2015, 6, 213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bajic, D.; Sanchez, A. The Ecology and Evolution of Microbial Metabolic Strategies. Curr. Opin. Biotechnol. 2020, 62, 123–128. [Google Scholar] [CrossRef]
- Somerville, V.; Grigaitis, P.; Battjes, J.; Moro, F.; Teusink, B. Use and Limitations of Genome-Scale Metabolic Models in Food Microbiology. Curr. Opin. Food Sci. 2022, 43, 225–231. [Google Scholar] [CrossRef]
- Wang, X.; Policarpio, L.; Prajapati, D.; Li, Z.; Zhang, H. Developing E. Coli-E. Coli Co-Cultures to Overcome Barriers of Heterologous Tryptamine Biosynthesis. Metab. Eng. Commun. 2020, 10, e00110. [Google Scholar] [CrossRef]
- Ferenci, T. Trade-off Mechanisms Shaping the Diversity of Bacteria. Trends Microbiol. 2016, 24, 209–223. [Google Scholar] [CrossRef]
- Bachmann, H.; Molenaar, D.; Branco Dos Santos, F.; Teusink, B. Experimental Evolution and the Adjustment of Metabolic Strategies in Lactic Acid Bacteria. FEMS Microbiol. Rev. 2017, 41, S201–S219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwentner, A.; Feith, A.; Münch, E.; Busche, T.; Rückert, C.; Kalinowski, J.; Takors, R.; Blombach, B. Metabolic Engineering to Guide Evolution—Creating a Novel Mode for L-Valine Production with Corynebacterium Glutamicum. Metab. Eng. 2018, 47, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, Á.; Vila, J.C.C.; Chang, C.-Y.; Diaz-Colunga, J.; Estrela, S.; Rebolleda-Gomez, M. Directed Evolution of Microbial Communities. Annu. Rev. Biophys. 2021, 50, 323–341. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Reed, J.L. Adaptive Evolution of Synthetic Cooperating Communities Improves Growth Performance. PLoS ONE 2014, 9, e108297. [Google Scholar] [CrossRef]
- Konstantinidis, D.; Pereira, F.; Geissen, E.; Grkovska, K.; Kafkia, E.; Jouhten, P.; Kim, Y.; Devendran, S.; Zimmermann, M.; Patil, K.R. Adaptive Laboratory Evolution of Microbial Co-Cultures for Improved Metabolite Secretion. Mol. Syst. Biol. 2021, 17, e10189. [Google Scholar] [CrossRef]
- Zamboni, N. 13C Metabolic Flux Analysis in Complex Systems. Curr. Opin. Biotechnol. 2011, 22, 103–108. [Google Scholar] [CrossRef]
- Gebreselassie, N.A.; Antoniewicz, M.R. 13C-Metabolic Flux Analysis of Co-Cultures: A Novel Approach. Metab. Eng. 2015, 31, 132–139. [Google Scholar] [CrossRef]
- Ghosh, A.; Nilmeier, J.; Weaver, D.; Adams, P.D.; Keasling, J.D.; Mukhopadhyay, A.; Petzold, C.J.; Martín, H.G. A Peptide-Based Method for 13C Metabolic Flux Analysis in Microbial Communities. PLoS Comput. Biol. 2014, 10, e1003827. [Google Scholar] [CrossRef] [Green Version]
- Wijaya, A.W.; Ulmer, A.; Hundsdorfer, L.; Verhagen, N.; Teleki, A.; Takors, R. Compartment-Specific Metabolome Labeling Enables the Identification of Subcellular Fluxes That May Serve as Promising Metabolic Engineering Targets in CHO Cells. Bioprocess Biosyst. Eng. 2021, 44, 2567–2578. [Google Scholar] [CrossRef]
- Stevens, K.A.; Jaykus, L.A. Bacterial Separation and Concentration from Complex Sample Matrices: A Review. Crit. Rev. Microbiol. 2004, 30, 7–24. [Google Scholar] [CrossRef]
- Even, S.; Lindley, N.D.; Cocaign-Bousquet, M. Transcriptional, Translational and Metabolic Regulation of Glycolysis in Lactococcus Lactis Subsp. Cremoris MG 1363 Grown in Continuous Acidic Cultures. Microbiology 2003, 149, 1935–1944. [Google Scholar] [CrossRef] [PubMed]
- Goers, L.; Freemont, P.; Polizzi, K.M. Co-Culture Systems and Technologies: Taking Synthetic Biology to the next Level. J. R. Soc. Interface 2014, 11, 20140065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burmeister, A.; Hilgers, F.; Langner, A.; Westerwalbesloh, C.; Kerkhoff, Y.; Tenhaef, N.; Drepper, T.; Kohlheyer, D.; von Lieres, E.; Noack, S.; et al. A Microfluidic Co-Cultivation Platform to Investigate Microbial Interactions at Defined Microenvironments. Lab Chip 2019, 19, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Burmeister, A.; Grünberger, A. Microfluidic Cultivation and Analysis Tools for Interaction Studies of Microbial Co-Cultures. Curr. Opin. Biotechnol. 2020, 62, 106–115. [Google Scholar] [CrossRef]
- Hesselman, M.C.; Odoni, D.I.; Ryback, B.M.; de Groot, S.; van Heck, R.G.A.; Keijsers, J.; Kolkman, P.; Nieuwenhuijse, D.; van Nuland, Y.M.; Sebus, E.; et al. A Multi-Platform Flow Device for Microbial (Co-) Cultivation and Microscopic Analysis. PLoS ONE 2012, 7, e36982. [Google Scholar] [CrossRef] [Green Version]
- Hyun, J.K.; Boedicker, J.Q.; Jang, W.C.; Ismagilov, R.F. Defined Spatial Structure Stabilizes a Synthetic Multispecies Bacterial Community. Proc. Natl. Acad. Sci. USA 2008, 105, 18188–18193. [Google Scholar] [CrossRef] [Green Version]
- Pörtner, R.; Märkl, H. Dialysis Cultures. Appl. Microbiol. Biotechnol. 1998, 50, 403–414. [Google Scholar] [CrossRef]
- Courtin, P.; Monnet, V.; Rul, F. Cell-Wall Proteinases PrtS and PrtB Have a Different Role in Streptococcus thermophilus/Lactobacillus bulgaricus Mixed Cultures in Milk. Microbiology 2002, 148, 3413–3421. [Google Scholar] [CrossRef] [Green Version]
- Liu, E.; Zheng, H.; Hao, P.; Konno, T.; Yu, Y.; Kume, H.; Oda, M.; Ji, Z.S. A Model of Proteolysis and Amino Acid Biosynthesis for Lactobacillus Delbrueckii Subsp. Bulgaricus in Whey. Curr. Microbiol. 2012, 65, 742–751. [Google Scholar] [CrossRef]
- Liu, E.; Hao, P.; Konno, T.; Yu, Y.; Oda, M.; Zheng, H.; Ji, Z. Amino Acid Biosynthesis and Proteolysis in Lactobacillus Bulgaricus Revisited: A Genomic Comparison. Sci. Res. 2012, 2012, 61–77. [Google Scholar]
- Sieuwerts, S.; Molenaar, D.; Van Hijum, S.A.F.T.; Beerthuyzen, M.; Stevens, M.J.A.; Janssen, P.W.M.; Ingham, C.J.; De Bok, F.A.M.; De Vos, W.M.; Van Hylckama Vlieg, J.E.T. Mixed-Culture Transcriptome Analysis Reveals the Molecular Basis of Mixed-Culture Growth in Streptococcus thermophilus and Lactobacillus bulgaricus. Appl. Environ. Microbiol. 2010, 76, 7775–7784. [Google Scholar] [CrossRef] [PubMed]
- Pastink, M.I.; Teusink, B.; Hols, P.; Visser, S.; De Vos, W.M.; Hugenholtz, J. Genome-Scale Model of Streptococcus thermophilus LMG18311 for Metabolic Comparison of Lactic Acid Bacteria. Appl. Environ. Microbiol. 2009, 75, 3627–3633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Serrano, G.M.; García-Garibay, J.M.; Cruz-Guerrero, A.E.; Gómez-Ruiz, L.D.C.; Ayala-Niño, A.; Castañeda-Ovando, A.; González-Olivares, L.G. Proteolytic System of Streptococcus thermophilus. J. Microbiol. Biotechnol. 2018, 28, 1581–1588. [Google Scholar] [CrossRef] [PubMed]
- Markakiou, S.; Gaspar, P.; Johansen, E.; Zeidan, A.A.; Neves, A.R. Harnessing the Metabolic Potential of Streptococcus thermophilus for New Biotechnological Applications. Curr. Opin. Biotechnol. 2020, 61, 142–152. [Google Scholar] [CrossRef]
- Alexandraki, V.; Kazou, M.; Blom, J.; Pot, B.; Papadimitriou, K.; Tsakalidou, E. Comparative Genomics of Streptococcus thermophilus Support Important Traits Concerning the Evolution, Biology and Technological Properties of the Species. Front. Microbiol. 2019, 10, 2916. [Google Scholar] [CrossRef]
- Hols, P.; Hancy, F.; Fontaine, L.; Grossiord, B.; Prozzi, D.; Leblond-Bourget, N.; Decaris, B.; Bolotin, A.; Delorme, C.; Ehrlich, S.D.; et al. New Insights in the Molecular Biology and Physiology of Streptococcus thermophilus Revealed by Comparative Genomics. FEMS Microbiol. Rev. 2005, 29, 435–463. [Google Scholar] [CrossRef]
- Li, S.; Tang, S.; He, Q.; Hu, J.; Zheng, J. Changes in Proteolysis in Fermented Milk Produced by Streptococcus thermophilus in Co-Culture with Lactobacillus Plantarum or Bifidobacterium animalis Subsp. Lactis during Refrigerated Storage. Molecules 2019, 24, 3699. [Google Scholar] [CrossRef] [Green Version]
- Liu, E.; Zheng, H.; Shi, T.; Ye, L.; Konno, T.; Oda, M.; Shen, H.; Ji, Z.S. Relationship between Lactobacillus bulgaricus and Streptococcus thermophilus under Whey Conditions: Focus on Amino Acid Formation. Int. Dairy J. 2016, 56, 141–150. [Google Scholar] [CrossRef]
- Letort, C.; Juillard, V. Development of a Minimal Chemically-Defined Medium for the Exponential Growth of Streptococcus thermophilus. J. Appl. Microbiol. 2001, 91, 1023–1029. [Google Scholar] [CrossRef]
- Mar, M.J.; Andersen, J.M.; Kandasamy, V.; Liu, J.; Solem, C.; Jensen, P.R. Biotechnology for Biofuels Synergy at Work: Linking the Metabolism of Two Lactic Acid Bacteria to Achieve Superior Production of 2—Butanol. Biotechnol. Biofuels 2020, 13, 45. [Google Scholar] [CrossRef] [Green Version]
- Teusink, B.; Molenaar, D. Systems Biology of Lactic Acid Bacteria: For Food and Thought. Curr. Opin. Syst. Biol. 2017, 6, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Ulmer, A.; Erdemann, F.; Mueller, S.; Loesch, M.; Wildt, S.; Jensen, M.L.; Gaspar, P.; Zeidan, A.A.; Takors, R. Differential Amino Acid Uptake and Depletion in Mono-Cultures and Co-Cultures of Streptococcus thermophilus and Lactobacillus delbrueckii Subsp. Bulgaricus in a Novel Semi-Synthetic Medium. Microorganisms 2022, 10, 1771. [Google Scholar] [CrossRef] [PubMed]
- Levenspiel, O. Tracer Technology. J. Chem. Inf. Model. 2019, 53, 1689–1699. [Google Scholar]
- Damköhler, G. Einflüsse Der Strömung, Diffusion Und Des Wärmeüberganges Auf Die Leistung von Reaktionsöfen. Z. Elektroch. Bd. 1937, 43, 1–13. [Google Scholar]
- Henderson, J.W.; Brooks, A. Improved Amino Acid Methods Using Agilent ZORBAX Eclipse Plus C18 Columns for a Variety of Agilent LC Instrumentation and Separation Goals. Agil. Technol. 2010, 5990-4547EN, 1–16. [Google Scholar]
- Teleki, A.; Sánchez-Kopper, A.; Takors, R. Alkaline Conditions in Hydrophilic Interaction Liquid Chromatography for Intracellular Metabolite Quantification Using Tandem Mass Spectrometry. Anal. Biochem. 2015, 475, 4–13. [Google Scholar] [CrossRef]
- Rau, M.H.; Gaspar, P.; Jensen, M.L.; Geppel, A.; Neves, A.R.; Zeidan, A.A. Genome-Scale Metabolic Modeling Combined with Transcriptome Profiling Provides Mechanistic Understanding of Streptococcus thermophilus CH8 Metabolism. Appl. Environ. Microbiol. 2022, 88, e00780-22. [Google Scholar] [CrossRef]
- Rau, M.H.; Zeidan, A.A. Constraint-Based Modeling in Microbial Food Biotechnology. In Biochemical Society Transactions; Portland Press Ltd.: London, UK, 2018; pp. 249–260. [Google Scholar] [CrossRef] [Green Version]
- Gottstein, W.; Olivier, B.G.; Bruggeman, F.J.; Teusink, B. Constraint-Based Stoichiometric Modelling from Single Organisms to Microbial Communities. J. R. Soc. Interface 2016, 13, 20160627. [Google Scholar] [CrossRef] [Green Version]
- Arioli, S.; Roncada, P.; Salzano, A.M.; Deriu, F.; Corona, S.; Guglielmetti, S.; Bonizzi, L.; Scaloni, A.; Mora, D. The Relevance of Carbon Dioxide Metabolism in Streptococcus thermophilus. Microbiology 2009, 155, 1953–1965. [Google Scholar] [CrossRef] [Green Version]
- Arioli, S.; Monnet, C.; Guglielmetti, S.; Parini, C.; de Noni, I.; Hogenboom, J.; Halami, P.M.; Mora, D. Aspartate Biosynthesis Is Essential for the Growth of Streptococcus thermophilus in Milk, and Aspartate Availability Modulates the Level of Urease Activity. Appl. Environ. Microbiol. 2007, 73, 5789–5796. [Google Scholar] [CrossRef] [Green Version]
- Bolotin, A.; Quinquis, B.; Renault, P.; Sorokin, A.; Ehrlich, S.D.; Kulakauskas, S.; Lapidus, A.; Goltsman, E.; Mazur, M.; Pusch, G.D.; et al. Complete Sequence and Comparative Genome Analysis of the Dairy Bacterium Streptococcus thermophilus. Nat. Biotechnol. 2004, 22, 1554–1558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loghmani, S.B.; Zitzow, E.; Koh, G.C.C.; Ulmer, A.; Veith, N.; Großeholz, R.; Rossnagel, M.; Loesch, M.; Aebersold, R.; Kreikemeyer, B.; et al. All Driven by Energy Demand? Integrative Comparison of Metabolism of Enterococcus Faecalis Wildtype and a Glutamine Synthase Mutant. Microbiol. Spectr. 2022, 10, e02400-21. [Google Scholar] [CrossRef] [PubMed]
- Khandelwal, R.A.; Olivier, B.G.; Röling, W.F.M.; Teusink, B.; Bruggeman, F.J. Community Flux Balance Analysis for Microbial Consortia at Balanced Growth. PLoS ONE 2013, 8, e64567. [Google Scholar] [CrossRef] [PubMed]
- Herve-Jimenez, L.; Guillouard, I.; Guedon, E.; Boudebbouze, S.; Hols, P.; Monnet, V.; Maguin, E.; Rul, F. Postgenomic Analysis of Streptococcus thermophilus Cocultivated in Milk with Lactobacillus delbrueckii Subsp. Bulgaricus: Involvement of Nitrogen, Purine, and Iron Metabolism. Appl. Environ. Microbiol. 2009, 75, 2062–2073. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| mean amino acid consumption | −Qi | 3.0 ± 2.8 µM × h−1 |
| mean amino acid dilution | D× ci | 11.4 ± 10.1 µM × h−1 |
| mean trans-membrane amino acid influx | ki × gi | 5.5 ± 3.8 µM × h−1 |
| mean change in amino acid concentration | dci/dt | 13.5 ± 13.6 µM × h−1 |
| amino acid feed | D × ci,feed | 0 µM × h−1 (feed medium without amino acids) |
| Damkoehler term for consumption | Daconsumption | 0.6 ± 0.4 |
| Damkoehler term for dilution | Dadilution | 2.3 ± 2.1 |
| Damkoehler number | Datotal | 2.9 ± 2.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ulmer, A.; Veit, S.; Erdemann, F.; Freund, A.; Loesch, M.; Teleki, A.; Zeidan, A.A.; Takors, R. A Two-Compartment Fermentation System to Quantify Strain-Specific Interactions in Microbial Co-Cultures. Bioengineering 2023, 10, 103. https://doi.org/10.3390/bioengineering10010103
Ulmer A, Veit S, Erdemann F, Freund A, Loesch M, Teleki A, Zeidan AA, Takors R. A Two-Compartment Fermentation System to Quantify Strain-Specific Interactions in Microbial Co-Cultures. Bioengineering. 2023; 10(1):103. https://doi.org/10.3390/bioengineering10010103
Chicago/Turabian StyleUlmer, Andreas, Stefan Veit, Florian Erdemann, Andreas Freund, Maren Loesch, Attila Teleki, Ahmad A. Zeidan, and Ralf Takors. 2023. "A Two-Compartment Fermentation System to Quantify Strain-Specific Interactions in Microbial Co-Cultures" Bioengineering 10, no. 1: 103. https://doi.org/10.3390/bioengineering10010103
APA StyleUlmer, A., Veit, S., Erdemann, F., Freund, A., Loesch, M., Teleki, A., Zeidan, A. A., & Takors, R. (2023). A Two-Compartment Fermentation System to Quantify Strain-Specific Interactions in Microbial Co-Cultures. Bioengineering, 10(1), 103. https://doi.org/10.3390/bioengineering10010103