


Bioinformatics Analysis, Expression Profiling, and Functional Characterization of Heat Shock Proteins in Wolfi-poria cocos

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials and Heat Treatment
2.2. W. cocos Sequence Analysis and Phylogenetic Tree Construction
2.3. Physiochemical Properties of Proteins and in Silico Protein-Protein Analysis
2.4. RNA Extraction and cDNA Synthesis
2.5. Expression Analysis by Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.6. Cloning of Heat Shock Protein Target Gene
2.7. Construction of Recombinant Plasmid Prokaryotic Expression Vector
2.8. Prokaryotic Expression and Heat Resistance
3. Results
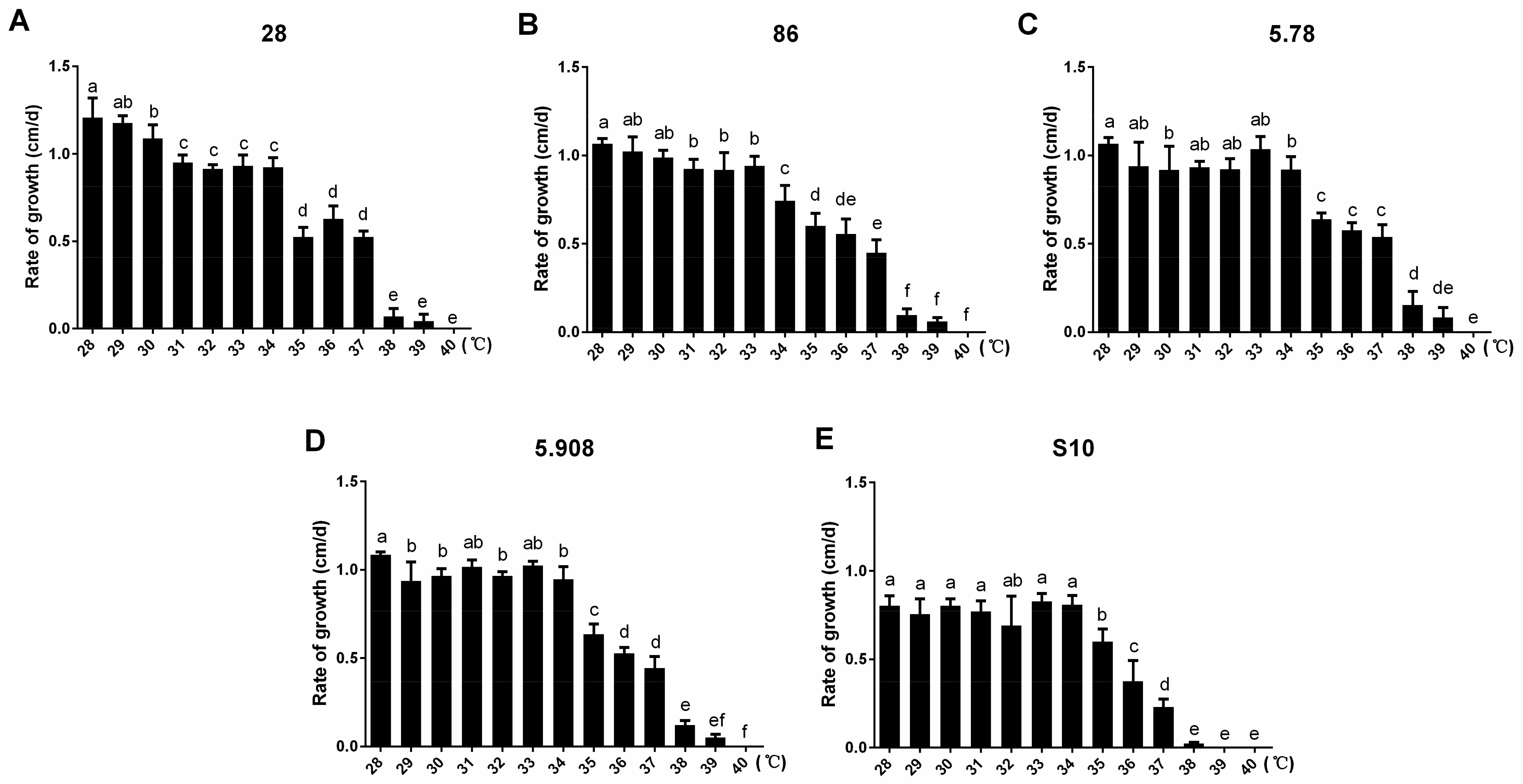
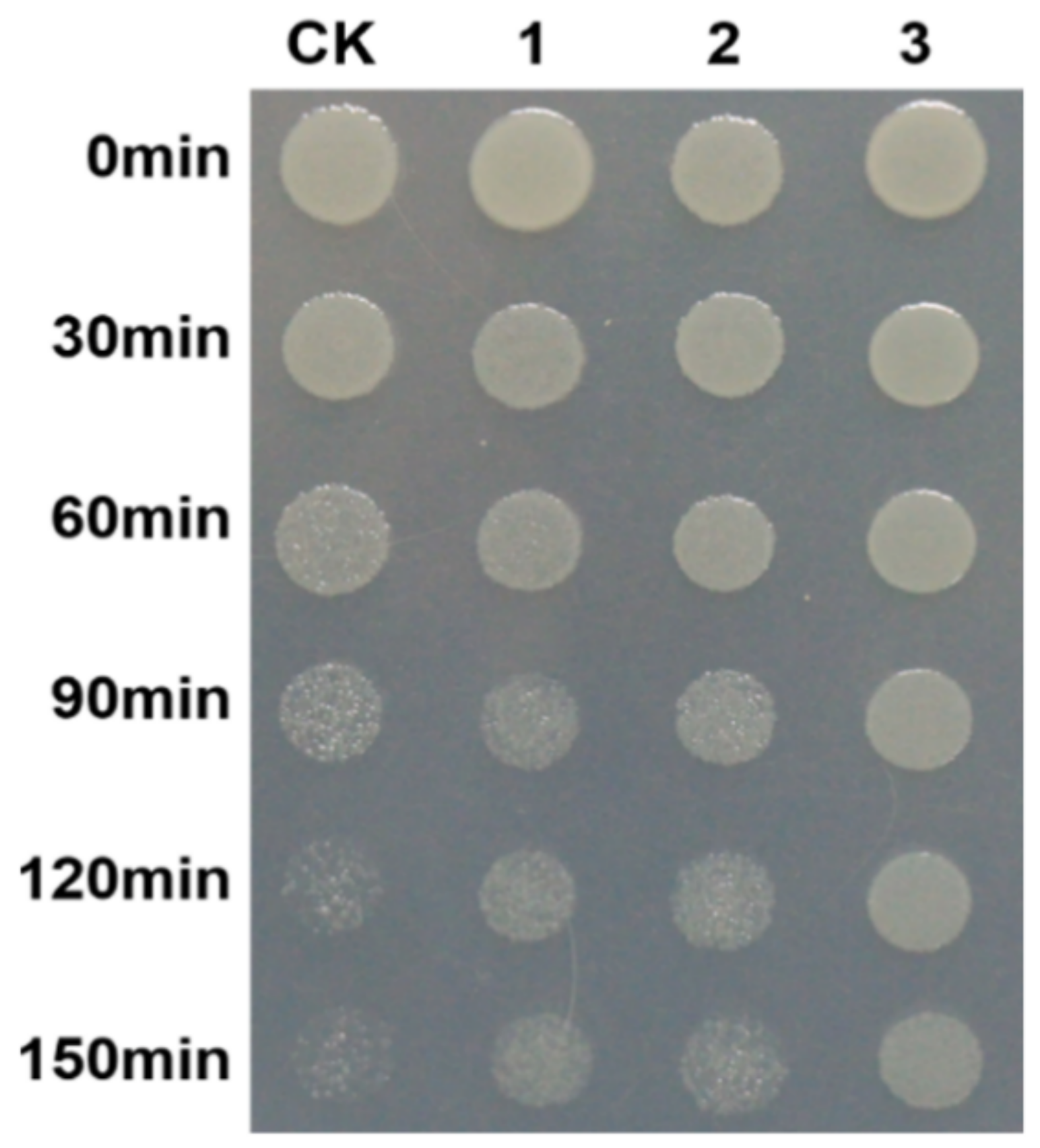
3.1. The Heat-Resistant Analysis of Different W. cocos Strains
3.2. Identification and Characterization of HSP Family Genes in W. cocos
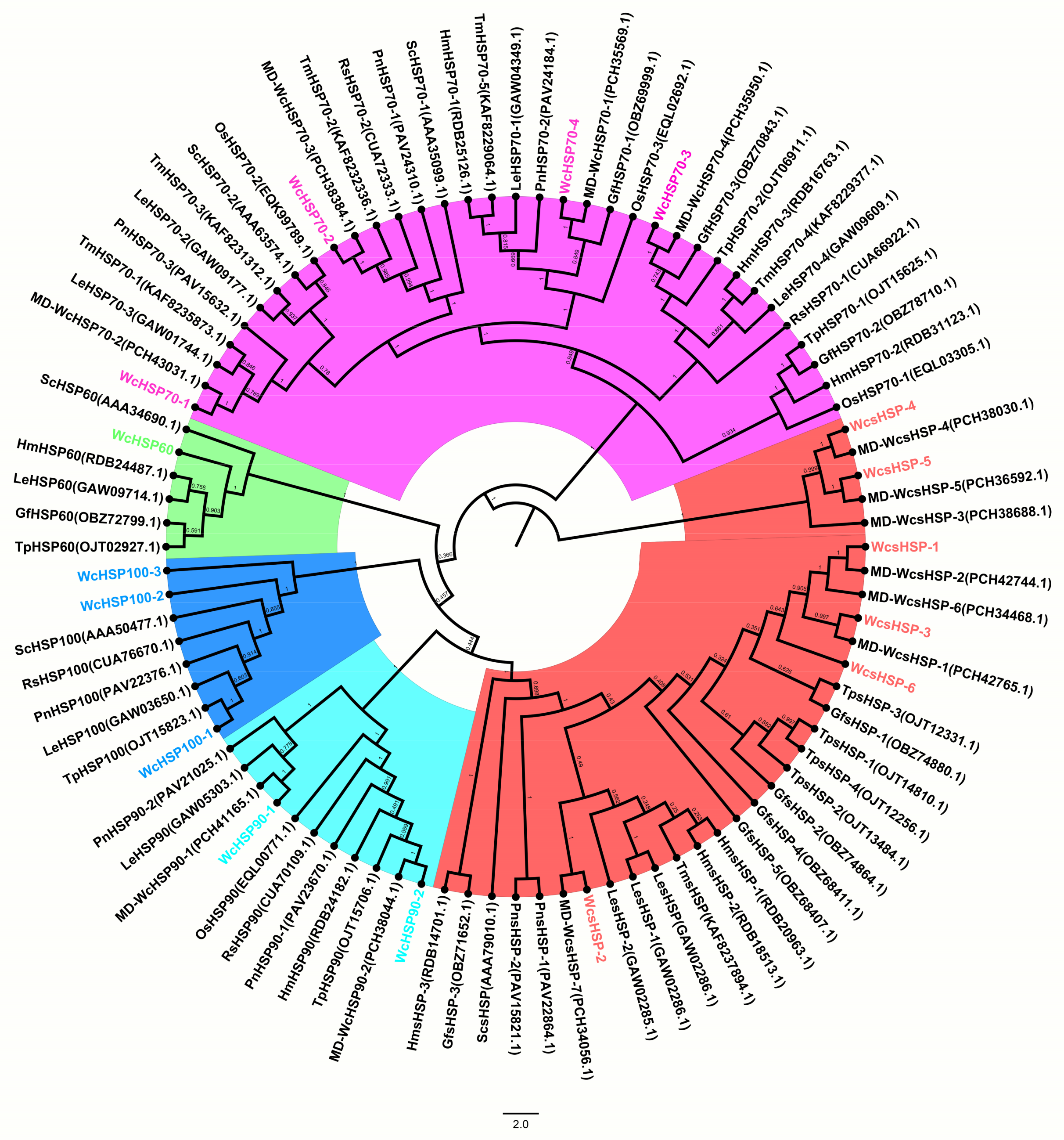
3.3. Phylogenetic Analysis of Heat Shock Protein Family Genes in W. cocos
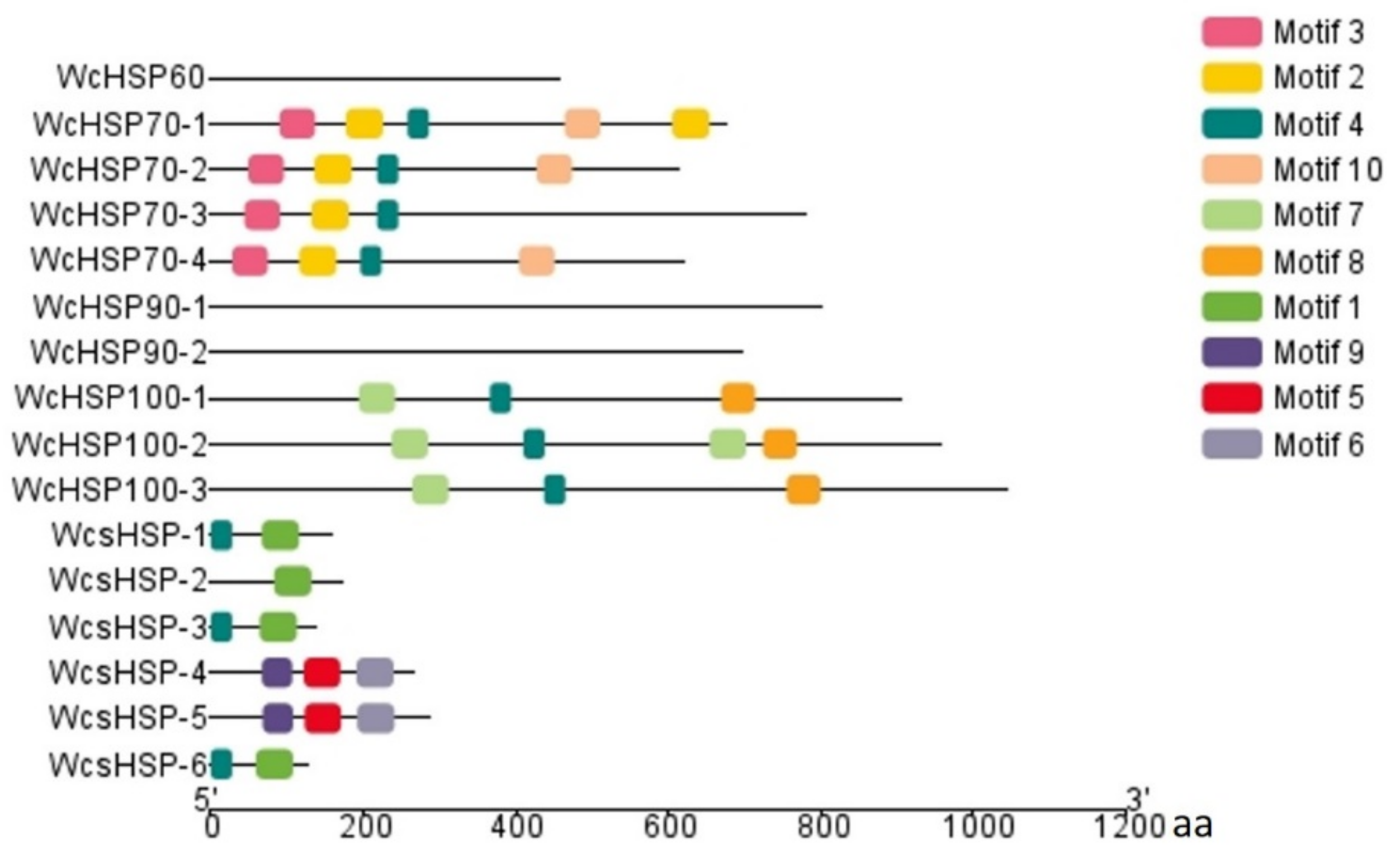
3.4. Conserved Motif Analysis of Heat Shock Protein in W. cocos
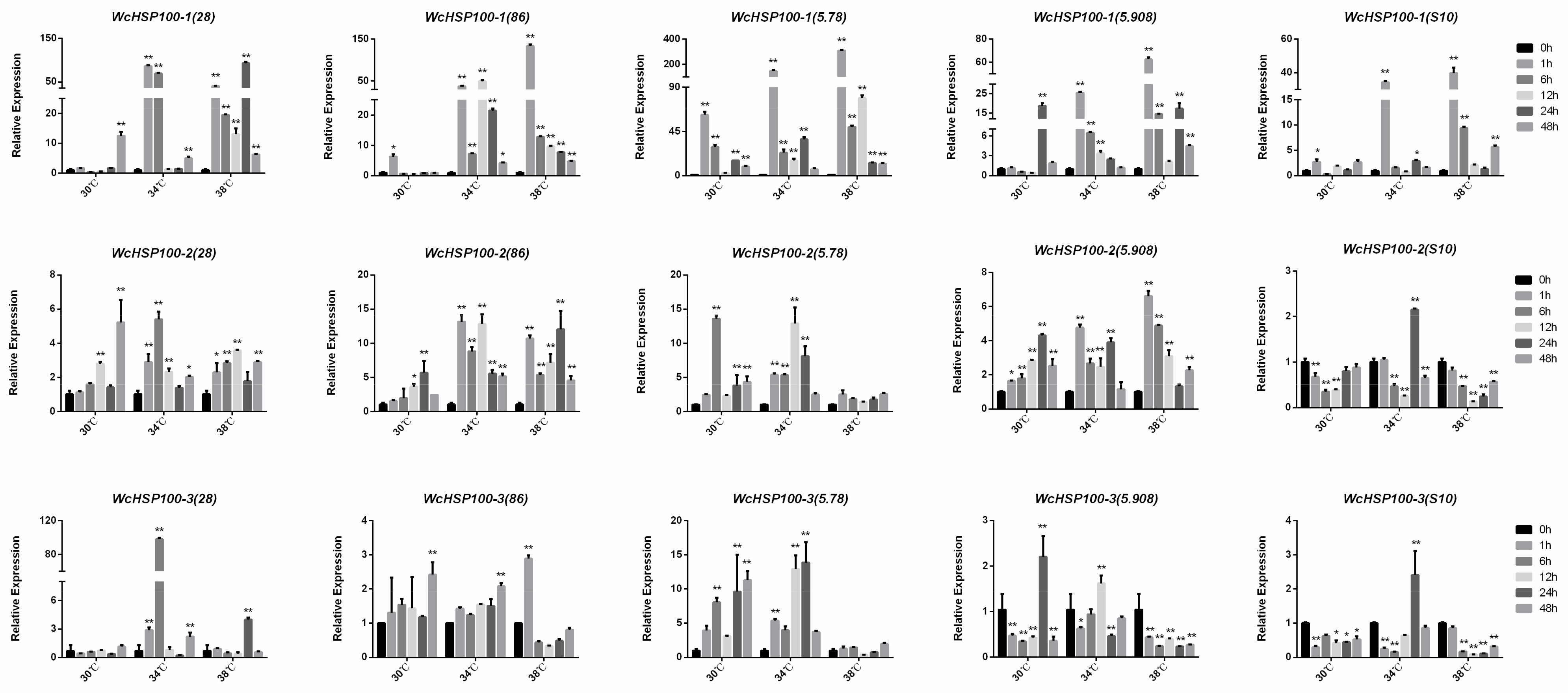
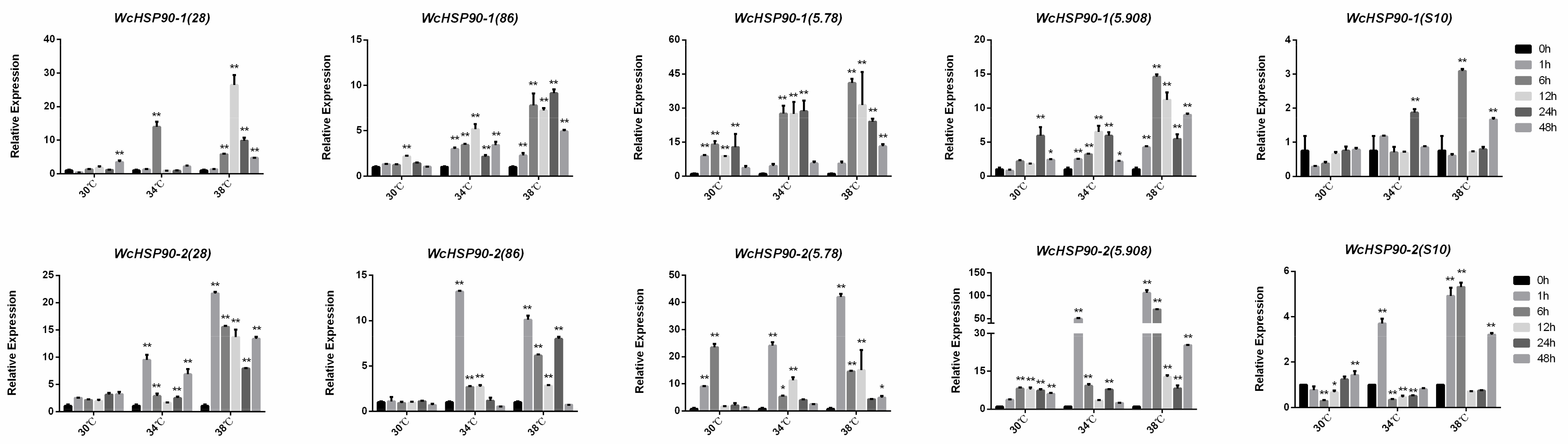
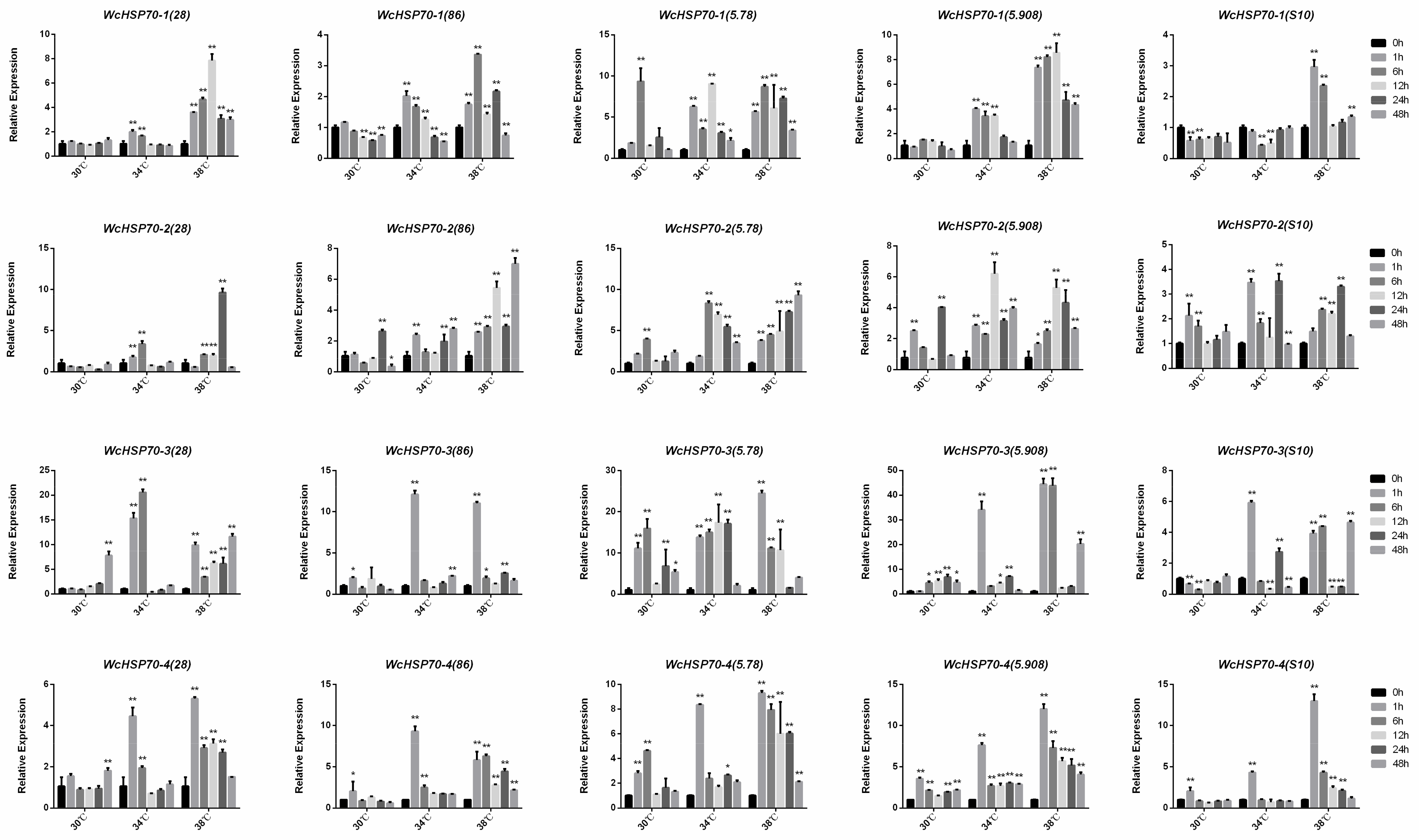
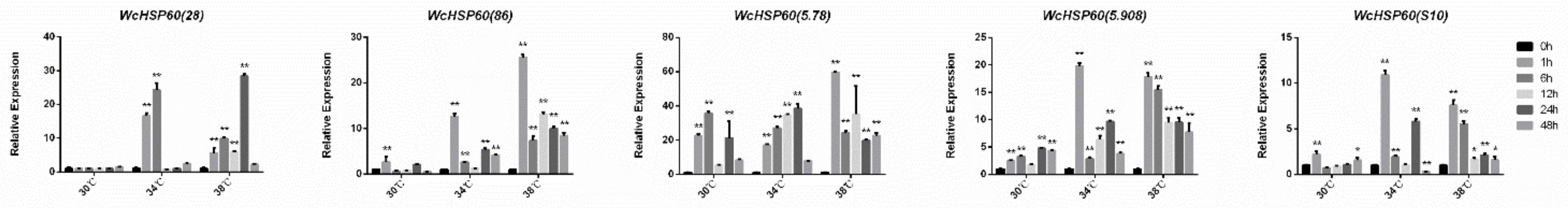
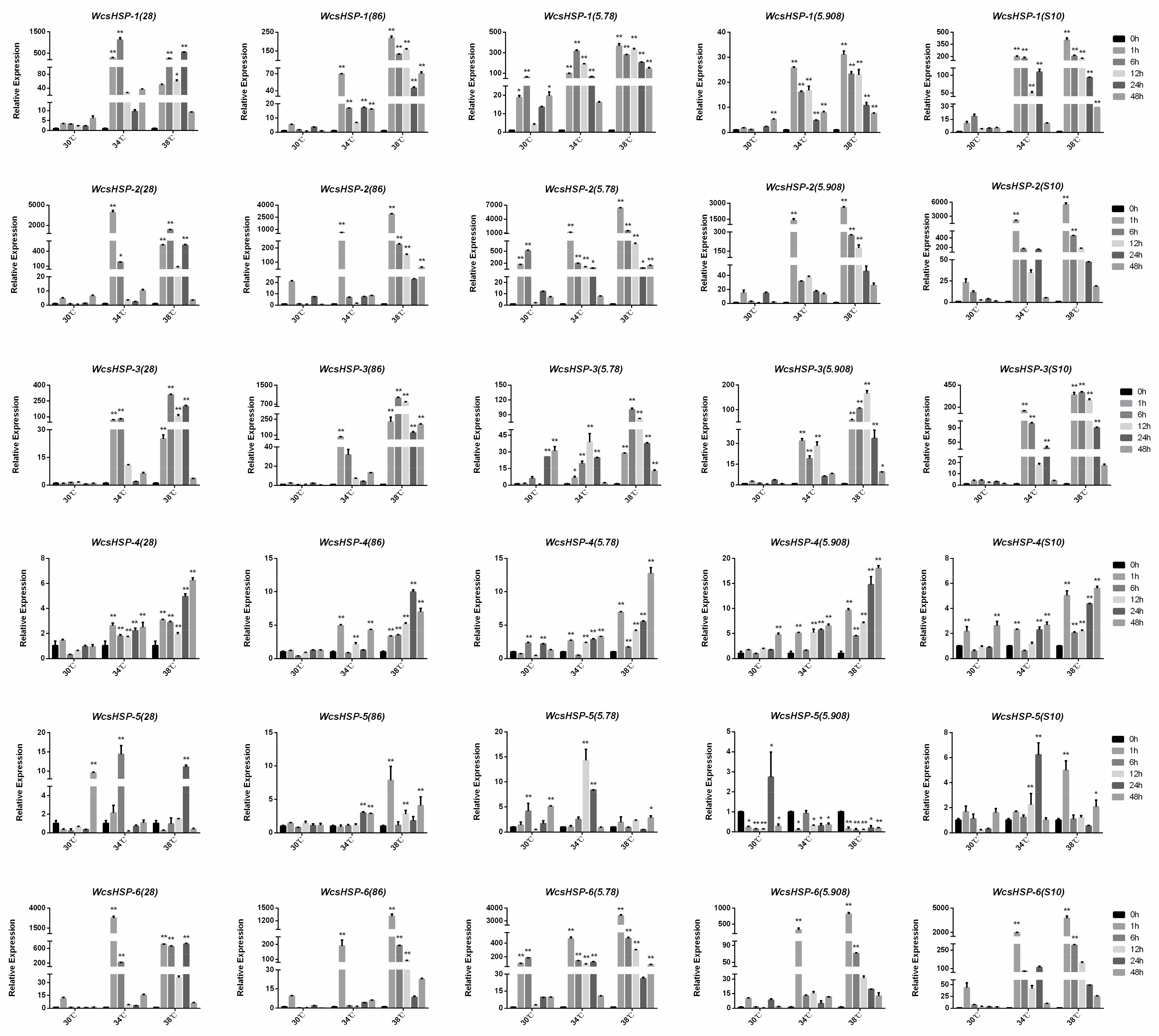
3.5. Expression Patterns of Heat Shock Proteins at Different Temperatures in W. cocos
3.6. Prokaryotic Expression and Thermal Function Analysis of Heat Shock Protein of W. cocos
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zininga, T.; Ramatsui, L.; Shonhai, A. Heat Shock Proteins as Immunomodulants. Molecules 2018, 23, 2846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Roh, S.H.; Lee, J.; Sung, N.; Liu, J.; Tsai, F.T.F. Cryo-EM Structures of the Hsp104 Protein Disaggregase Captured in the ATP Conformation. Cell Rep. 2019, 26, 29–36.e3. [Google Scholar] [CrossRef] [Green Version]
- Bösl, B.; Grimminger, V.; Walter, S. The Molecular Chaperone Hsp104—A Molecular Machine for Protein Disaggregation. J. Struct. Biol. 2006, 156, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.E.; Queitsch, C.; Toft, D. Hsp90: From Structure to Phenotype. Nat. Struct. Mol. Biol. 2004, 11, 1152–1155. [Google Scholar] [CrossRef] [PubMed]
- Żwirowski, S.; Kłosowska, A.; Obuchowski, I.; Nillegoda, N.B.; Piróg, A.; Ziętkiewicz, S.; Bukau, B.; Mogk, A.; Liberek, K. Hsp70 Displaces Small Heat Shock Proteins from Aggregates to Initiate Protein Refolding. EMBO J. 2017, 36, 783–796. [Google Scholar] [CrossRef] [Green Version]
- Jones, G.W.; Masison, D.C. Saccharomyces Cerevisiae Hsp70 Mutations Affect [PSI +] Prion Propagation and Cell Growth Differently and Implicate Hsp40 and Tetratricopeptide Repeat Cochaperones in Impairment of [PSI +]. Genetics 2003, 163, 495–506. [Google Scholar] [CrossRef]
- Ranson, N.A.; White, H.E.; Saibil, H.R. Chaperonins. Biochem. J. 1998, 333, 233–242. [Google Scholar] [CrossRef] [Green Version]
- Kaufman, B.A.; Kolesar, J.E.; Perlman, P.S.; Butow, R.A. A Function for the Mitochondrial Chaperonin Hsp60 in the Structure and Transmission of Mitochondrial DNA Nucleoids in Saccharomyces Cerevisiae. J. Cell Biol. 2003, 163, 457–461. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, H.; Fukui, N.; Adachi, M.; Saiki, E.; Yamasaki, A.; Matsumura, R.; Kuroyanagi, D.; Hongo, K.; Mizobata, T.; Kawata, Y. Human Molecular Chaperone Hsp60 and Its Apical Domain Suppress Amyloid Fibril Formation of α-Synuclein. Int. J. Mol. Sci. 2019, 21, 47. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, T.; Yamamoto, H.; Kudo, I.; Matsumoto, K.; Odaka, M.; Grave, E.; Itoh, H. HSP60 Possesses a GTPase Activity and Mediates Protein Folding with HSP10. Sci. Rep. 2017, 7, 16931. [Google Scholar] [CrossRef] [Green Version]
- Craig, E.A.; Gambill, B.D.; Nelson, R.J. Heat Shock Proteins: Molecular Chaperones of Protein Biogenesis. Microbiol. Rev. 1993, 57, 402–414. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of Plant Heat-Shock Proteins and Molecular Chaperones in the Abiotic Stress Response. Trends Plant Sci. 2004, 9, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Moore, M.L.; Six, D.L. Effects of Temperature on Growth, Sporulation, and Competition of Mountain Pine Beetle Fungal Symbionts. Microb. Ecol. 2015, 70, 336–347. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Huang, W.; Xiong, C.; Zhao, J. Transcriptome Analysis Reveals the Role of Nitric Oxide in Pleurotus Eryngii Responses to Cd2+ Stress. Chemosphere 2018, 201, 294–302. [Google Scholar] [CrossRef]
- Zhang, X.; Ren, A.; Li, M.-J.; Cao, P.-F.; Chen, T.-X.; Zhang, G.; Shi, L.; Jiang, A.-L.; Zhao, M.-W. Heat Stress Modulates Mycelium Growth, Heat Shock Protein Expression, Ganoderic Acid Biosynthesis, and Hyphal Branching of Ganoderma Lucidum via Cytosolic Ca2+. Appl. Environ. Microbiol. 2016, 82, 4112–4125. [Google Scholar] [CrossRef] [Green Version]
- Rayner, A.D.M. The Challenge of the Individualistic Mycelium. Mycologia 1991, 83, 48–71. [Google Scholar] [CrossRef]
- Solheim, H. Early Stages of Blue-Stain Fungus Invasion of Lodgepole Pine Sapwood Following Mountain Pine Beetle Attack. Can. J. Bot. 1995, 73, 70–74. [Google Scholar] [CrossRef]
- Six, D.L. Ecological and Evolutionary Determinants of Bark Beetle—Fungus Symbioses. Insects 2012, 3, 339–366. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; St. Leger, R.J.; Fang, W. Pyruvate Accumulation Is the First Line of Cell Defense against Heat Stress in a Fungus. mBio 2017, 8, e01284-17. [Google Scholar] [CrossRef] [Green Version]
- Weber, G.F. The Occurrence of Tuckahoes and Poria Cocos in Florida. Mycologia 1929, 21, 113–130. [Google Scholar] [CrossRef]
- Nie, A.; Chao, Y.; Zhang, X.; Jia, W.; Zhou, Z.; Zhu, C. Phytochemistry and Pharmacological Activities of Wolfiporia Cocos (F.A. Wolf) Ryvarden & Gilb. Front. Pharmacol. 2020, 11, 505249. [Google Scholar] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing Real-Time PCR Data by the Comparative C(T) Method. Protoc. Exch. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Richter, K.; Haslbeck, M.; Buchner, J. The Heat Shock Response: Life on the Verge of Death. Mol. Cell 2010, 40, 253–266. [Google Scholar] [CrossRef] [PubMed]
- McAlister, L.; Finkelstein, D.B. Heat Shock Proteins and Thermal Resistance in Yeast. Biochem. Biophys. Res. Commun. 1980, 93, 819–824. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.J.; Tang, S.F.; Jin, L.; Shao, C.X.; Yang, Y.; He, Y.L. Quality Evaluation and Genetic Relation Identification for Different Poria cocos Strains. J. South. Agric. 2018, 49, 8–13. [Google Scholar]
- Xiong, J.; Lin, F.C.; Wang, K.Q.; Su, W.; Fu, J. Studies on Basic Biological Characters of Wolfiporia cocos. Mycosystema 2006, 25, 446–453. [Google Scholar]
- Scharf, K.-D.; Berberich, T.; Ebersberger, I.; Nover, L. The Plant Heat Stress Transcription Factor (Hsf) Family: Structure, Function and Evolution. Biochim. Biophys. Acta BBA Gene Regul. Mech. 2012, 1819, 104–119. [Google Scholar] [CrossRef]
- Mager, W.H.; De Kruijff, A.J. Stress-Induced Transcriptional Activation. Microbiol. Rev. 1995, 59, 506–531. [Google Scholar] [CrossRef]
- Wang, G.-Z.; Ma, C.-J.; Luo, Y.; Zhou, S.-S.; Zhou, Y.; Ma, X.-L.; Cai, Y.-L.; Yu, J.-J.; Bian, Y.-B.; Gong, Y.-H. Proteome and Transcriptome Reveal Involvement of Heat Shock Proteins and Indoleacetic Acid Metabolism Process in Lentinula Edodes Thermotolerance. Cell. Physiol. Biochem. 2018, 50, 1617–1637. [Google Scholar] [CrossRef]
- O’Connor, E.; Owens, R.A.; Doyle, S.; Amini, A.; Grogan, H.; Fitzpatrick, D.A. Proteomic Investigation of Interhyphal Interactions between Strains of Agaricus Bisporus. Fungal Biol. 2020, 124, 579–591. [Google Scholar] [CrossRef]
- Wu, M.-X.; Zou, Y.; Yu, Y.-H.; Chen, B.-X.; Zheng, Q.-W.; Ye, Z.-W.; Wei, T.; Ye, S.-Q.; Guo, L.-Q.; Lin, J.-F. Comparative Transcriptome and Proteome Provide New Insights into the Regulatory Mechanisms of the Postharvest Deterioration of Pleurotus Tuoliensis Fruitbodies during Storage. Food Res. Int. 2021, 147, 110540. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Sun, J.; Ning, H.; Qin, Z.; Miao, Y.; Sun, T.; Zhang, X. De Novo Transcriptome Sequencing and Comprehensive Analysis of the Heat Stress Response Genes in the Basidiomycetes Fungus Ganoderma Lucidum. Gene 2018, 661, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Kurahashi, A.; Sato, M.; Nishibori, K.; Fujimori, F. Heat Shock Protein 9 MRNA Expression Increases during Fruiting Body Differentiation in Grifola Frondosa and Other Edible Mushrooms. Mycoscience 2014, 55, 98–102. [Google Scholar] [CrossRef]
- Kiang, J. Heat Shock Protein 70 KDa Molecular Biology, Biochemistry, and Physiology. Pharmacol. Ther. 1998, 80, 183–201. [Google Scholar] [CrossRef]
- Zou, Y.; Zhang, M.; Qu, J.; Zhang, J. ITRAQ-Based Quantitative Proteomic Analysis Reveals Proteomic Changes in Mycelium of Pleurotus Ostreatus in Response to Heat Stress and Subsequent Recovery. Front. Microbiol. 2018, 9, 2368. [Google Scholar] [CrossRef]
- Chen, C.; Li, Q.; Wang, Q.; Lu, D.; Zhang, H.; Wang, J.; Fu, R. Transcriptional Profiling Provides New Insights into the Role of Nitric Oxide in Enhancing Ganoderma Oregonense Resistance to Heat Stress. Sci. Rep. 2017, 7, 15694. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Wang, Z.; Tang, R.; Wang, L.; Chen, C.; Ren, Z. Genome-Wide Identification and Expression Analysis of Hsf and Hsp Gene Families in Cucumber (Cucumis Sativus L.). Plant Growth Regul. 2021, 95, 223–239. [Google Scholar] [CrossRef]
- Pirkkala, L.; Nykänen, P.; Sistonen, L. Roles of the Heat Shock Transcription Factors in Regulation of the Heat Shock Response and Beyond. FASEB J. 2001, 15, 1118–1131. [Google Scholar] [CrossRef] [Green Version]
- Leach, M.D.; Farrer, R.A.; Tan, K.; Miao, Z.; Walker, L.A.; Cuomo, C.A.; Wheeler, R.T.; Brown, A.J.P.; Wong, K.H.; Cowen, L.E. Hsf1 and Hsp90 Orchestrate Temperature-Dependent Global Transcriptional Remodelling and Chromatin Architecture in Candida Albicans. Nat. Commun. 2016, 7, 11704. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Luo, Y.; Chen, W.; Gong, Y.; Zhou, Y.; Bian, Y. The DnaJ Gene Family in Shiitake Culinary-Medicinal Mushroom, Lentinus Edodes (Agaricomycetes): Comprehensive Identification, Characterization, and Expression Profiles under Different Conditions. Int. J. Med. Mushrooms 2019, 21, 909–919. [Google Scholar] [CrossRef]
- Xu, L.; Guo, L.; Yu, H. Label-Free Comparative Proteomics Analysis Revealed Heat Stress Responsive Mechanism in Hypsizygus Marmoreus. Front. Microbiol. 2021, 11, 541967. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Gao, J.; Guo, L.; Yu, H. Heat Shock Protein 70 (HmHsp70) from Hypsizygus Marmoreus Confers Thermotolerance to Tobacco. AMB Express 2020, 10, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.-O.; Jeong, M.-J.; Kwon, T.-R.; Lee, S.-K.; Byun, M.-O.; Chung, I.-M.; Park, S.-C. Pleurotus Sajor-Caju HSP100 Complements a Thermotolerance Defect Inhsp104 MutantSaccharomyces Cerevisiae. J. Biosci. 2006, 31, 223–233. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Amino Acid Residue Base | Molecular Formula | Molecular Weight (kDa) | Isoelectric Point | Fat Soluble Index | Subcellular Localization |
|---|---|---|---|---|---|---|
| WcHSP100-1 | 904 | C4385H7195N1267O1354S23 | 100.07 | 5.68 | 100.5 | cytoplasm |
| WcHSP100-2 | 956 | C4688H7615N1383O1429S32 | 107.24 | 6.53 | 91.59 | nucleus |
| WcHSP100-3 | 1043 | C5144H8237N1447O1601S28 | 116.87 | 5.31 | 95.68 | nucleus |
| WcHSP90-1 | 800 | C4019H6315N1073O1273S17 | 90.58 | 4.75 | 84.53 | endoplasmic reticulum |
| WcHSP90-2 | 695 | C3512H5586N906O1116S18 | 78.935 | 4.91 | 86.85 | cytoplasm |
| WcHSP70-1 | 675 | C3252H5227N903O1042S13 | 74.06 | 5.15 | 84.8 | endoplasmic reticulum |
| WcHSP70-2 | 612 | C2940H4765N829O933S13 | 67.07 | 5.43 | 87.53 | cytoplasm |
| WcHSP70-3 | 780 | C3799H6073N1057O1187S17 | 86.09 | 5.23 | 85.68 | cytoplasm |
| WcHSP70-4 | 619 | C2945H4816N826O946S13 | 67.35 | 5.42 | 89.18 | cytoplasm |
| WcHSP60 | 456 | C2091H3444N578O674S14 | 47.91 | 4.99 | 96.32 | cytoplasm |
| WcsHSP-1 | 158 | C775H1214N226O246S3 | 17.73 | 5.93 | 64.81 | nucleus |
| WcsHSP-2 | 173 | C842H1321N247O267S7 | 19.42 | 5.74 | 70.69 | nucleus |
| WcsHSP-3 | 137 | C684H1079N203O210S4 | 15.63 | 9.03 | 72.55 | nucleus |
| WcsHSP-4 | 266 | C1398H2078N332O356S9 | 29.52 | 6.2 | 110 | cytoplasmic membrane |
| WcsHSP-5 | 287 | C1464H2201N357O385S11 | 31.32 | 5.81 | 109.2 | cytoplasmic membrane |
| WcsHSP-6 | 128 | C627H984N180O204S2 | 14.37 | 5.38 | 75.39 | nucleus |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, X.; Tang, X.; Zhou, Y.; ahmad, B.; Zhang, D.; Zeng, Y.; Wei, J.; Deng, L.; Chen, S.; Pan, Y. Bioinformatics Analysis, Expression Profiling, and Functional Characterization of Heat Shock Proteins in Wolfi-poria cocos. Bioengineering 2023, 10, 390. https://doi.org/10.3390/bioengineering10030390
Hu X, Tang X, Zhou Y, ahmad B, Zhang D, Zeng Y, Wei J, Deng L, Chen S, Pan Y. Bioinformatics Analysis, Expression Profiling, and Functional Characterization of Heat Shock Proteins in Wolfi-poria cocos. Bioengineering. 2023; 10(3):390. https://doi.org/10.3390/bioengineering10030390
Chicago/Turabian StyleHu, Xin, Xue Tang, Yumei Zhou, Bilal ahmad, Deli Zhang, Yue Zeng, Jingyi Wei, Liling Deng, Shijiang Chen, and Yu Pan. 2023. "Bioinformatics Analysis, Expression Profiling, and Functional Characterization of Heat Shock Proteins in Wolfi-poria cocos" Bioengineering 10, no. 3: 390. https://doi.org/10.3390/bioengineering10030390
APA StyleHu, X., Tang, X., Zhou, Y., ahmad, B., Zhang, D., Zeng, Y., Wei, J., Deng, L., Chen, S., & Pan, Y. (2023). Bioinformatics Analysis, Expression Profiling, and Functional Characterization of Heat Shock Proteins in Wolfi-poria cocos. Bioengineering, 10(3), 390. https://doi.org/10.3390/bioengineering10030390