



Optimizing Bioink Composition for Human Chondrocyte Expression of Lubricin

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Engineering of Lubricin Promoter-Driven Reporter Human Chondrocytes (HuPRG4gLuc)
2.2. HuPRG4gLuc Reporter Cell Aggregate Culture and TGFβ1 Dose Response
2.3. Luciferase Assessment
2.4. Histology
2.5. Biochemical Assays
2.6. HuPRG4gLuc Reporter Cells qPCR Assessment
2.7. Design of Experiment (DoE) Screen Design
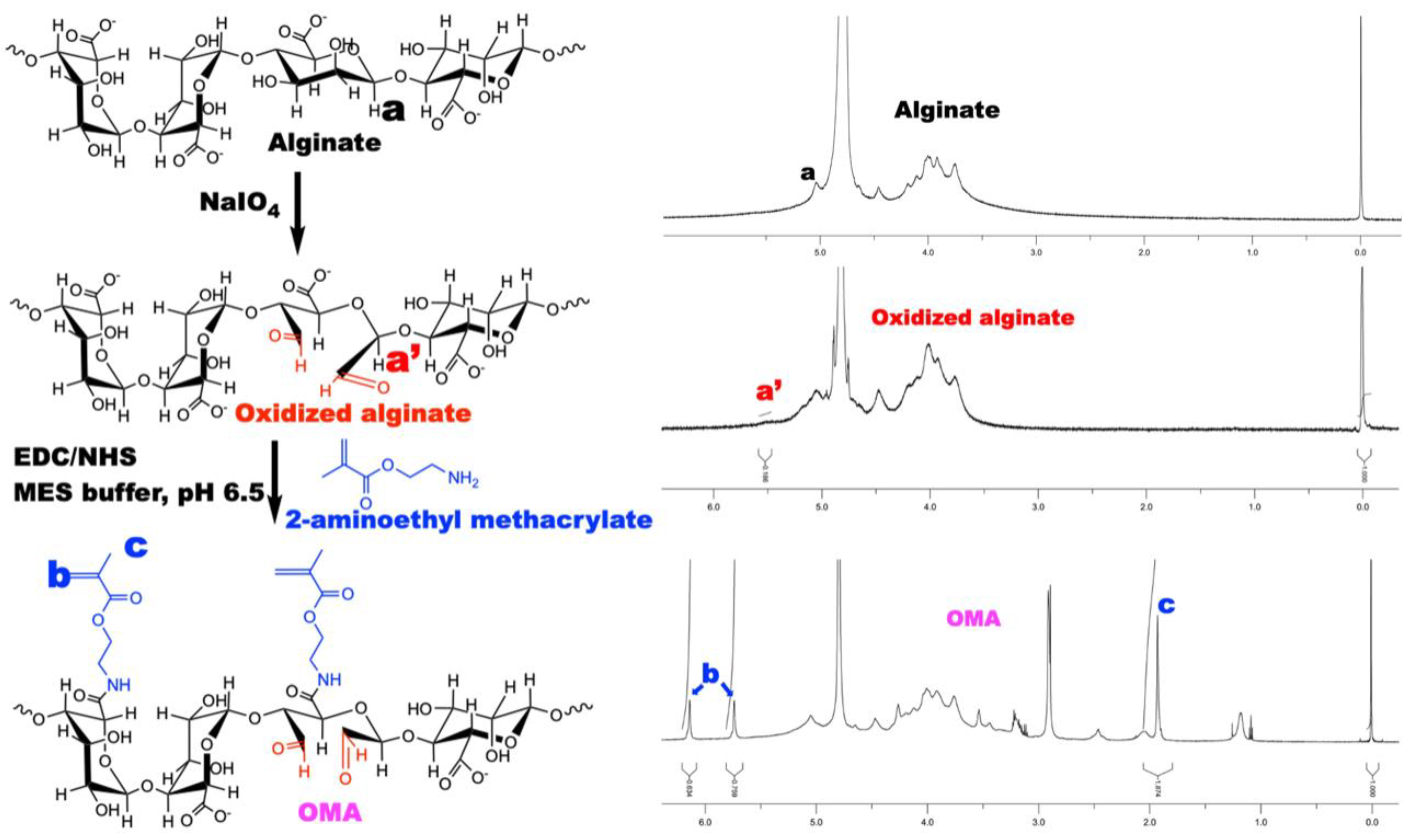
2.8. Oxidized Methacrylated Alginate (OMA) Synthesis and Characterization
2.9. Preparation of GelMA and OMA Stocks and Combinations
2.10. 3D Bioprinting
2.11. Cell Viability
2.12. Mechanical Characterization
2.12.1. Dynamic Mechanical Analysis (DMA)
2.12.2. Lap-Shear
2.13. Swelling and Degradation
2.14. Lubricin (PRG4) ELISA
2.15. Statistical Analysis
3. Results
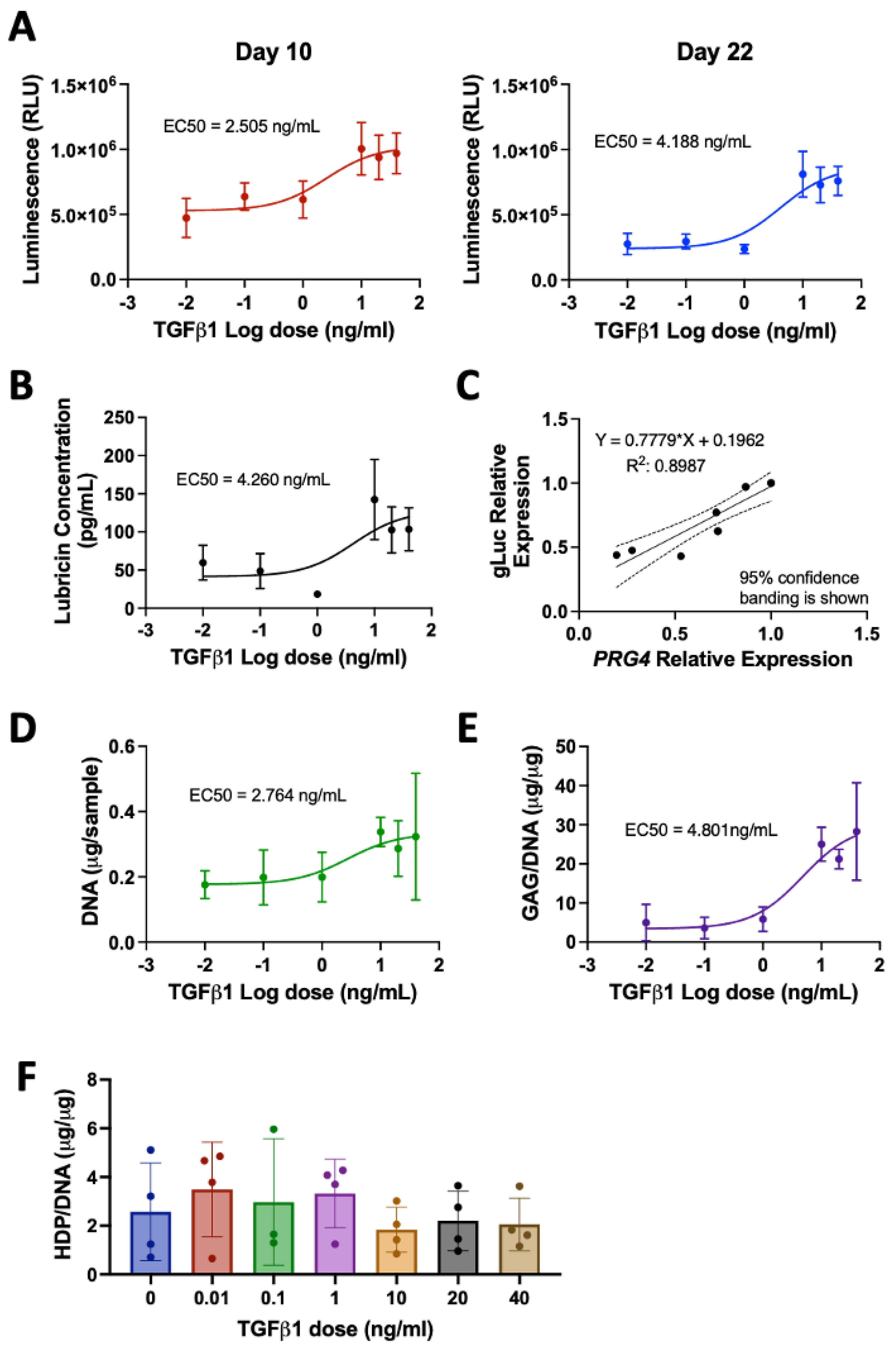
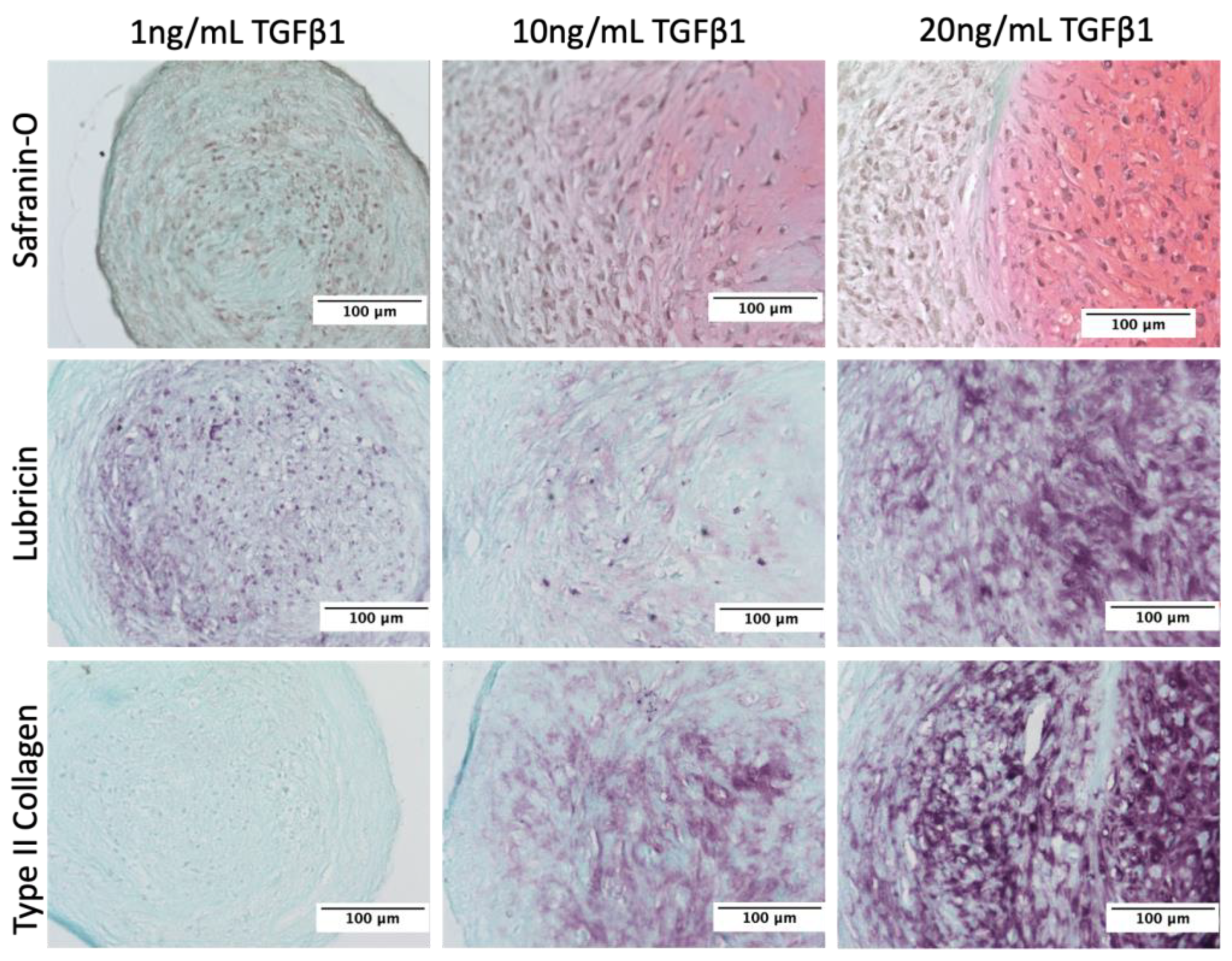
3.1. Stimulation of Lubricin by TGFβ1 in Human Primary Articular Chondrocytes
3.2. Characterization of OMA
3.3. Design of Experiment (DoE) Screen of GelMA and OMA Combinations for Lubricin Expression
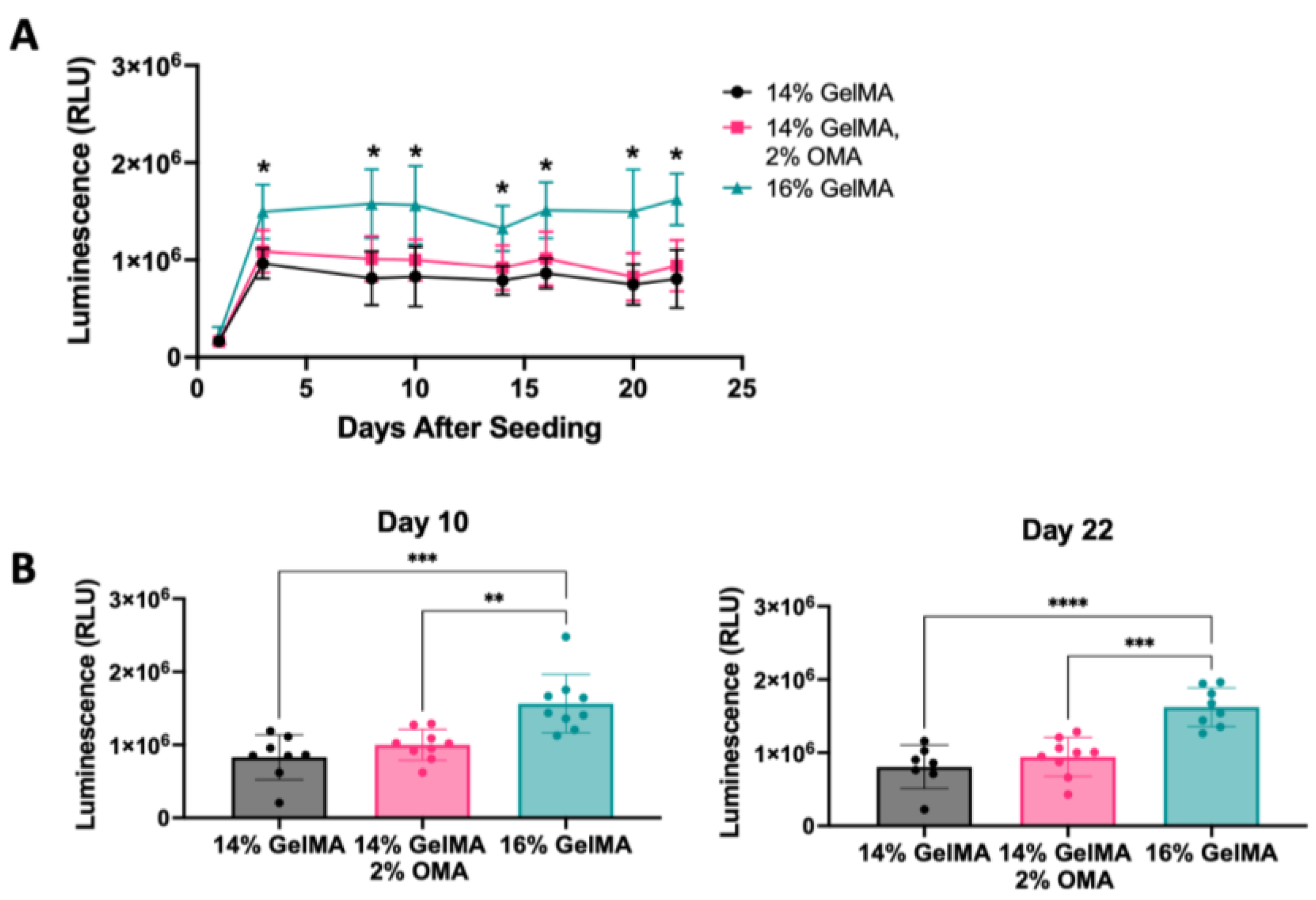
3.4. Validation of the DoE Screen
3.5. Cell Viability
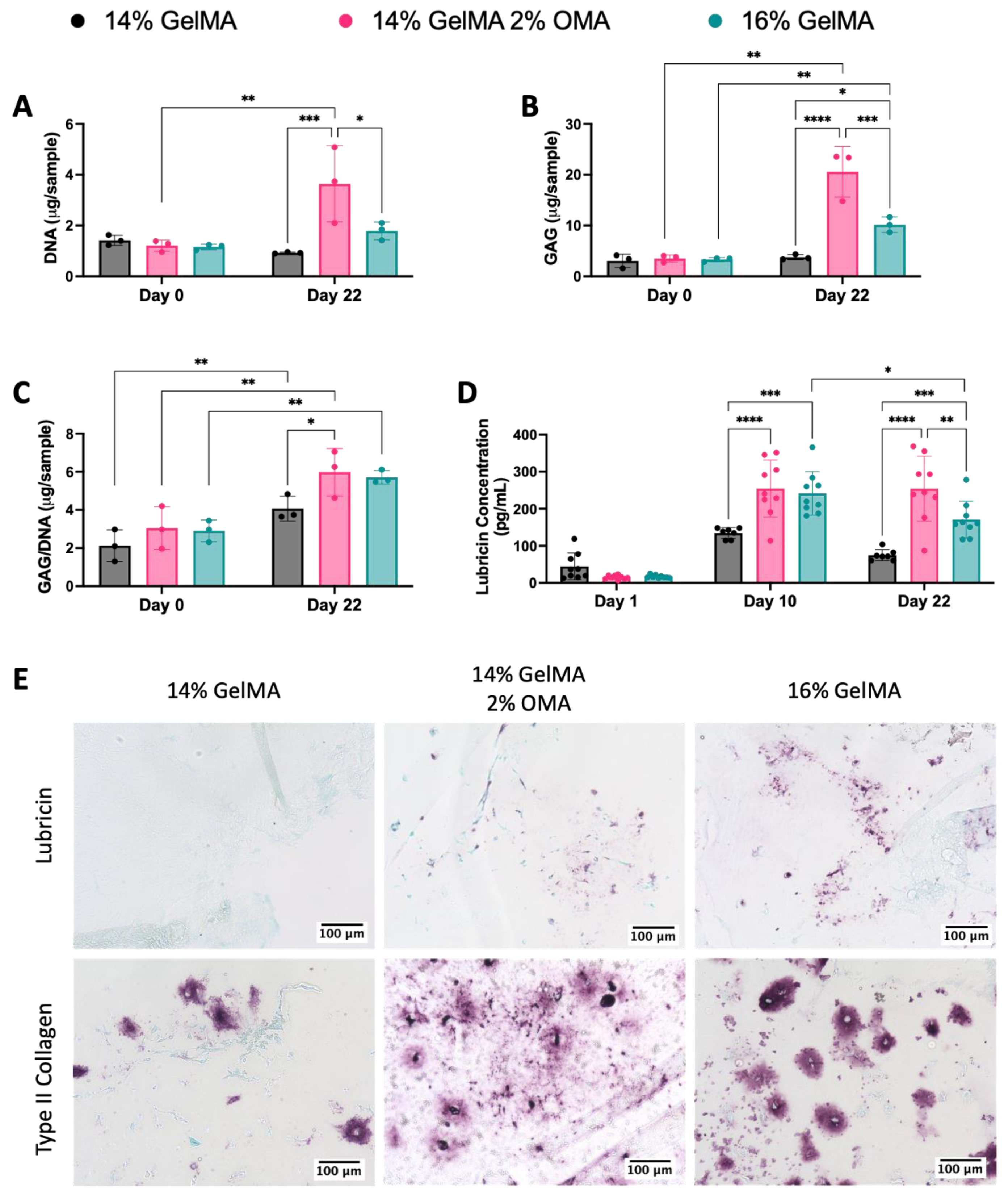
3.6. 3D Bioprinted Disc Construct Containing HuPRG4gLuc Cells
3.7. Mechanical Characterization of Disc Constructs
3.8. Shape Fidelity of the Bioprinted Constructs and Degradation of Bioinks
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hunziker, E.B.; Lippuner, K.; Keel, M.J.B.; Shintani, N. An Educational Review of Cartilage Repair: Precepts & Practice--Myths & Misconceptions--Progress & Prospects. Osteoarthr. Cartil. 2015, 23, 334–350. [Google Scholar] [CrossRef]
- Klein, T.J.; Rizzi, S.C.; Reichert, J.C.; Georgi, N.; Malda, J.; Schuurman, W.; Crawford, R.W.; Hutmacher, D.W. Strategies for Zonal Cartilage Repair Using Hydrogels. Macromol. Biosci. 2009, 9, 1049–1058. [Google Scholar] [CrossRef] [PubMed]
- Niikura, T.; Reddi, A.H. Differential Regulation of Lubricin/Superficial Zone Protein by Transforming Growth Factor Beta/Bone Morphogenetic Protein Superfamily Members in Articular Chondrocytes and Synoviocytes. Arthritis Rheum. 2007, 56, 2312–2321. [Google Scholar] [CrossRef]
- Hollander, A.P.; Pidoux, I.; Reiner, A.; Rorabeck, C.; Bourne, R.; Poole, A.R. Damage to Type II Collagen in Aging and Osteoarthritis Starts at the Articular Surface, Originates around Chondrocytes, and Extends into the Cartilage with Progressive Degeneration. J. Clin. Investig. 1995, 96, 2859–2869. [Google Scholar] [CrossRef] [PubMed]
- Hauselmann, H.J.; Flechtenmacher, J.; Michal, L.; Thonar, E.J.; Shinmei, M.; Kuettner, K.E.; Aydelotte, M.B. The Superficial Layer of Human Articular Cartilage Is More Susceptible to Interleukin-1-Induced Damage than the Deeper Layers. Arthritis Rheum. 1996, 39, 478–488. [Google Scholar] [CrossRef]
- Sophia Fox, A.J.; Bedi, A.; Rodeo, S.A. The Basic Science of Articular Cartilage: Structure, Composition, and Function. Sports Health 2009, 1, 461–468. [Google Scholar] [CrossRef]
- Thielen, N.G.M.; van der Kraan, P.M.; van Caam, A.P.M. TGFβ/BMP Signaling Pathway in Cartilage Homeostasis. Cells 2019, 8, 969. [Google Scholar] [CrossRef]
- Lee, Y.; Choi, J.; Hwang, N.S. Regulation of Lubricin for Functional Cartilage Tissue Regeneration: A Review. Biomater. Res. 2018, 22, 9. [Google Scholar] [CrossRef]
- Chang, D.P.; Abu-Lail, N.I.; Coles, J.M.; Guilak, F.; Jay, G.D.; Zauscher, S. Friction Force Microscopy of Lubricin and Hyaluronic Acid between Hydrophobic and Hydrophilic Surfaces. Soft Matter 2009, 5, 3438–3445. [Google Scholar] [CrossRef]
- Rhee, D.K.; Marcelino, J.; Baker, M.; Gong, Y.; Smits, P.; Lefebvre, V.; Jay, G.D.; Stewart, M.; Wang, H.; Warman, M.L.; et al. The Secreted Glycoprotein Lubricin Protects Cartilage Surfaces and Inhibits Synovial Cell Overgrowth. J. Clin. Investig. 2005, 115, 622–631. [Google Scholar] [CrossRef]
- Watkins, A.R.; Reesink, H.L. Lubricin in Experimental and Naturally Occurring Osteoarthritis: A Systematic Review. Osteoarthr. Cartil. 2020, 28, 1303–1315. [Google Scholar] [CrossRef] [PubMed]
- Nemirov, D.; Nakagawa, Y.; Sun, Z.; Lebaschi, A.; Wada, S.; Carballo, C.; Deng, X.-H.; Putnam, D.; Bonassar, L.J.; Rodeo, S.A. Effect of Lubricin Mimetics on the Inhibition of Osteoarthritis in a Rat Anterior Cruciate Ligament Transection Model. Am. J. Sports Med. 2020, 48, 624–634. [Google Scholar] [CrossRef] [PubMed]
- Flannery, C.R.; Zollner, R.; Corcoran, C.; Jones, A.R.; Root, A.; Rivera-Bermúdez, M.A.; Blanchet, T.; Gleghorn, J.P.; Bonassar, L.J.; Bendele, A.M.; et al. Prevention of Cartilage Degeneration in a Rat Model of Osteoarthritis by Intraarticular Treatment with Recombinant Lubricin. Arthritis Rheum. 2009, 60, 840–847. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Xu, C.; Li, X.; Song, J.; Yu, B. Treatment with Recombinant Lubricin Attenuates Osteoarthritis by Positive Feedback Loop between Articular Cartilage and Subchondral Bone in Ovariectomized Rats. Bone 2015, 74, 37–47. [Google Scholar] [CrossRef]
- Murray, I.R.; Benke, M.T.; Mandelbaum, B.R. Management of Knee Articular Cartilage Injuries in Athletes: Chondroprotection, Chondrofacilitation, and Resurfacing. Knee Surg. Sports Traumatol. Arthrosc. 2016, 24, 1617–1626. [Google Scholar] [CrossRef]
- Zhang, L.; Yang, G.; Johnson, B.N.; Jia, X. Three-Dimensional (3D) Printed Scaffold and Material Selection for Bone Repair. Acta Biomater. 2019, 84, 16–33. [Google Scholar] [CrossRef]
- Kean, T.J.; Thanou, M. Utility of Chitosan for 3D Printing and Bioprinting. In Sustainable Agriculture Reviews 35: Chitin and Chitosan: History, Fundamentals and Innovations; Crini, G., Lichtfouse, E., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 271–292. ISBN 978-3-030-16538-3. [Google Scholar]
- Chan, B.P.; Leong, K.W. Scaffolding in Tissue Engineering: General Approaches and Tissue-Specific Considerations. Eur. Spine J. 2008, 17 (Suppl. S4), 467–479. [Google Scholar] [CrossRef]
- You, F.; Eames, B.F.; Chen, X. Application of Extrusion-Based Hydrogel Bioprinting for Cartilage Tissue Engineering. Int. J. Mol. Sci. 2017, 18, 1597. [Google Scholar] [CrossRef]
- Fan, Y.; Yue, Z.; Lucarelli, E.; Wallace, G.G. Hybrid Printing Using Cellulose Nanocrystals Reinforced GelMA/HAMA Hydrogels for Improved Structural Integration. Adv. Healthc. Mater. 2020, 9, 2001410. [Google Scholar] [CrossRef]
- Costantini, M.; Idaszek, J.; Szöke, K.; Jaroszewicz, J.; Dentini, M.; Barbetta, A.; Brinchmann, J.E.; Święszkowski, W. 3D Bioprinting of BM-MSCs-Loaded ECM Biomimetic Hydrogels for in Vitro Neocartilage Formation. Biofabrication 2016, 8, 035002. [Google Scholar] [CrossRef]
- O’Connell, C.D.; Onofrillo, C.; Duchi, S.; Li, X.; Zhang, Y.; Tian, P.; Lu, L.; Trengove, A.; Quigley, A.; Gambhir, S.; et al. Evaluation of Sterilisation Methods for Bio-Ink Components: Gelatin, Gelatin Methacryloyl, Hyaluronic Acid and Hyaluronic Acid Methacryloyl. Biofabrication 2019, 11, 035003. [Google Scholar] [CrossRef] [PubMed]
- Lam, T.; Dehne, T.; Krüger, J.P.; Hondke, S.; Endres, M.; Thomas, A.; Lauster, R.; Sittinger, M.; Kloke, L. Photopolymerizable Gelatin and Hyaluronic Acid for Stereolithographic 3D Bioprinting of Tissue-engineered Cartilage. J. Biomed. Mater. Res. B Appl. Biomater. 2019, 107, 2649–2657. [Google Scholar] [CrossRef] [PubMed]
- Kessel, B.; Lee, M.; Bonato, A.; Tinguely, Y.; Tosoratti, E.; Zenobi-Wong, M. 3D Bioprinting of Macroporous Materials Based on Entangled Hydrogel Microstrands. Adv. Sci. 2020, 7, 2001419. [Google Scholar] [CrossRef] [PubMed]
- Martyniak, K.; Lokshina, A.; Cruz, M.A.; Karimzadeh, M.; Kemp, R.; Kean, T.J. Biomaterial Composition and Stiffness as Decisive Properties of 3D Bioprinted Constructs for Type II Collagen Stimulation. Acta Biomater. 2022. [Google Scholar] [CrossRef]
- Wang, B.; Díaz-Payno, P.J.; Browe, D.C.; Freeman, F.E.; Nulty, J.; Burdis, R.; Kelly, D.J. Affinity-Bound Growth Factor within Sulfated Interpenetrating Network Bioinks for Bioprinting Cartilaginous Tissues. Acta Biomater. 2021, 128, 130–142. [Google Scholar] [CrossRef]
- Levato, R.; Webb, W.R.; Otto, I.A.; Mensinga, A.; Zhang, Y.; van Rijen, M.; van Weeren, R.; Khan, I.M.; Malda, J. The Bio in the Ink: Cartilage Regeneration with Bioprintable Hydrogels and Articular Cartilage-Derived Progenitor Cells. Acta Biomater. 2017, 61, 41–53. [Google Scholar] [CrossRef]
- Daly, A.C.; Cunniffe, G.M.; Sathy, B.N.; Jeon, O.; Alsberg, E.; Kelly, D.J. 3D Bioprinting of Developmentally Inspired Templates for Whole Bone Organ Engineering. Adv. Healthc. Mater. 2016, 5, 2353–2362. [Google Scholar] [CrossRef]
- Schipani, R.; Scheurer, S.; Florentin, R.; Critchley, S.E.; Kelly, D.J. Reinforcing Interpenetrating Network Hydrogels with 3D Printed Polymer Networks to Engineer Cartilage Mimetic Composites. Biofabrication 2020, 12, 035011. [Google Scholar] [CrossRef]
- Kosik-Koziol, A.; Costantini, M.; Mróz, A.; Idaszek, J.; Heljak, M.; Jaroszewicz, J.; Kijeńska, E.; Szöke, K.; Frerker, N.; Barbetta, A.; et al. 3D Bioprinted Hydrogel Model Incorporating β-Tricalcium Phosphate for Calcified Cartilage Tissue Engineering. Biofabrication 2019, 11, 035016. [Google Scholar] [CrossRef]
- Lim, K.S.; Levato, R.; Costa, P.F.; Castilho, M.D.; Alcala-Orozco, C.R.; van Dorenmalen, K.M.A.; Melchels, F.P.W.; Gawlitta, D.; Hooper, G.J.; Malda, J.; et al. Bio-Resin for High Resolution Lithography-Based Biofabrication of Complex Cell-Laden Constructs. Biofabrication 2018, 10, 034101. [Google Scholar] [CrossRef]
- Daly, A.C.; Critchley, S.E.; Rencsok, E.M.; Kelly, D.J. A Comparison of Different Bioinks for 3D Bioprinting of Fibrocartilage and Hyaline Cartilage. Biofabrication 2016, 8, 045002. [Google Scholar] [CrossRef] [PubMed]
- Irmak, G.; Gümüşderelioğlu, M. Photo-Activated Platelet-Rich Plasma (PRP)-Based Patient-Specific Bio-Ink for Cartilage Tissue Engineering. Biomed. Mater. 2020, 15, 065010. [Google Scholar] [CrossRef] [PubMed]
- De Moor, L.; Fernandez, S.; Vercruysse, C.; Tytgat, L.; Asadian, M.; De Geyter, N.; Van Vlierberghe, S.; Dubruel, P.; Declercq, H. Hybrid Bioprinting of Chondrogenically Induced Human Mesenchymal Stem Cell Spheroids. Front. Bioeng. Biotechnol. 2020, 8, 484. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Zhang, L.; Du, X.; Fan, Z.; Wang, L.; Sun, W.; Cheng, Y.; Zhu, Y.; Chen, C. Reversible Physical Crosslinking Strategy with Optimal Temperature for 3D Bioprinting of Human Chondrocyte-Laden Gelatin Methacryloyl Bioink. J. Biomater. Appl. 2018, 33, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Cantu, L.; Gleadall, A.; Faris, C.; Segal, J.; Shakesheff, K.; Yang, J. Multi-Material 3D Bioprinting of Porous Constructs for Cartilage Regeneration. Mater. Sci. Eng. C 2020, 109, 110578. [Google Scholar] [CrossRef]
- Zhu, W.; Cui, H.; Boualam, B.; Masood, F.; Flynn, E.; Rao, R.D.; Zhang, Z.-Y.; Zhang, L.G. 3D Bioprinting Mesenchymal Stem Cell-Laden Construct with Core–Shell Nanospheres for Cartilage Tissue Engineering. Nanotechnology 2018, 29, 185101. [Google Scholar] [CrossRef]
- Luo, C.; Xie, R.; Zhang, J.; Liu, Y.; Li, Z.; Zhang, Y.; Zhang, X.; Yuan, T.; Chen, Y.; Fan, W. Low-Temperature Three-Dimensional Printing of Tissue Cartilage Engineered with Gelatin Methacrylamide. Tissue Eng. Part. C: Methods 2020, 26, 306–316. [Google Scholar] [CrossRef]
- Jeon, O.; Bouhadir, K.H.; Mansour, J.M.; Alsberg, E. Photocrosslinked Alginate Hydrogels with Tunable Biodegradation Rates and Mechanical Properties. Biomaterials 2009, 30, 2724–2734. [Google Scholar] [CrossRef]
- Jeon, O.; Alt, D.S.; Ahmed, S.M.; Alsberg, E. The Effect of Oxidation on the Degradation of Photocrosslinkable Alginate Hydrogels. Biomaterials 2012, 33, 3503–3514. [Google Scholar] [CrossRef]
- Jeon, O.; Lee, Y.B.; Lee, S.J.; Guliyeva, N.; Lee, J.; Alsberg, E. Stem Cell-Laden Hydrogel Bioink for Generation of High Resolution and Fidelity Engineered Tissues with Complex Geometries. Bioact. Mater. 2022, 15, 185–193. [Google Scholar] [CrossRef]
- Xu, H.; Casillas, J.; Krishnamoorthy, S.; Xu, C. Effects of Irgacure 2959 and Lithium Phenyl-2,4,6-Trimethylbenzoylphosphinate on Cell Viability, Physical Properties, and Microstructure in 3D Bioprinting of Vascular-like Constructs. Biomed. Mater. 2020, 15, 055021. [Google Scholar] [CrossRef] [PubMed]
- Dennis, J.E.; Splawn, T.; Kean, T.J. High-Throughput, Temporal and Dose Dependent, Effect of Vitamins and Minerals on Chondrogenesis. Front. Cell Dev. Biol. 2020, 8, 92. [Google Scholar] [CrossRef] [PubMed]
- Cruz, M.A.; Gonzalez, Y.; Vélez Toro, J.A.; Karimzadeh, M.; Rubbo, A.; Morris, L.; Medam, R.; Splawn, T.; Archer, M.; Fernandes, R.J.; et al. Micronutrient Optimization for Tissue Engineered Articular Cartilage Production of Type II Collagen. Front. Bioeng. Biotechnol. 2023, 11, 1179332. [Google Scholar] [CrossRef] [PubMed]
- Ly, O.; Monchau, F.; Rémond, S.; Lors, C.; Jouanneaux, A.; Debarre, É.; Damidot, D. Optimization of the Formulation of an Original Hydrogel-Based Bone Cement Using a Mixture Design. J. Mech. Behav. Biomed. Mater. 2020, 110, 103886. [Google Scholar] [CrossRef]
- Eriksson, L.; Johansson, E.; Wikström, C. Mixture Design—Design Generation, PLS Analysis, and Model Usage. Chemom. Intell. Lab. Syst. 1998, 43, 1–24. [Google Scholar] [CrossRef]
- Kean, T.J.; Dennis, J.E. Synoviocyte Derived-Extracellular Matrix Enhances Human Articular Chondrocyte Proliferation and Maintains Re-Differentiation Capacity at Both Low and Atmospheric Oxygen Tensions. PLoS ONE 2015, 10, e0129961. [Google Scholar] [CrossRef]
- Truong, R.D.; Bernier, M.A.; Dennis, J.E.; Kean, T.J. Synoviocyte-Derived Extracellular Matrix and BFGF Speed Human Chondrocyte Proliferation While Maintaining Differentiation Potential. Front. Bioeng. Biotechnol. 2022, 10, 825005. [Google Scholar] [CrossRef]
- Lennon, D.P.; Haynesworth, S.E.; Bruder, S.P.; Jaiswal, N.; Caplan, A.I. Human and Animal Mesenchymal Progenitor Cells from Bone Marrow: Identification of Serum for Optimal Selection and Proliferation. Vitr. Cell.Dev.Biol.-Anim. 1996, 32, 602–611. [Google Scholar] [CrossRef]
- Dennis, J.E.; Whitney, G.A.; Rai, J.; Fernandes, R.J.; Kean, T.J. Physioxia Stimulates Extracellular Matrix Deposition and Increases Mechanical Properties of Human Chondrocyte-Derived Tissue-Engineered Cartilage. Front. Bioeng. Biotechnol. 2020, 8, 590743. [Google Scholar] [CrossRef]
- Kean, T.J.; Mera, H.; Whitney, G.A.; MacKay, D.L.; Awadallah, A.; Fernandes, R.J.; Dennis, J.E. Disparate Response of Articular- and Auricular-Derived Chondrocytes to Oxygen Tension. Connect. Tissue Res. 2016, 57, 319–333. [Google Scholar] [CrossRef]
- Schmidt, U.; Weigert, M.; Broaddus, C.; Myers, G. Cell Detection with Star-Convex Polygons. In Medical Image Computing and Computer Assisted Intervention–MICCAI 2018: 21st International Conference, Granada, Spain, 16–20 September 2018, Proceedings, Part II 11; Springer: Berlin/Heidelberg, Germany, 2018; Volume 11071, pp. 265–273. [Google Scholar]
- Schmidt, T.A.; Gastelum, N.S.; Han, E.H.; Nugent-Derfus, G.E.; Schumacher, B.L.; Sah, R.L. Differential Regulation of Proteoglycan 4 Metabolism in Cartilage by IL-1alpha, IGF-I, and TGF-Beta1. Osteoarthr. Cartil. 2008, 16, 90–97. [Google Scholar] [CrossRef]
- Pan, R.L.; Martyniak, K.; Karimzadeh, M.; Gelikman, D.G.; DeVries, J.; Sutter, K.; Coathup, M.; Razavi, M.; Sawh-Martinez, R.; Kean, T.J. Systematic Review on the Application of 3D-Bioprinting Technology in Orthoregeneration: Current Achievements and Open Challenges. J. Exp. Orthop. 2022, 9, 95. [Google Scholar] [CrossRef] [PubMed]
- Jadin, K.D.; Wong, B.L.; Bae, W.C.; Li, K.W.; Williamson, A.K.; Schumacher, B.L.; Price, J.H.; Sah, R.L. Depth-Varying Density and Organization of Chondrocytes in Immature and Mature Bovine Articular Cartilage Assessed by 3d Imaging and Analysis. J. Histochem. Cytochem. 2005, 53, 1109–1119. [Google Scholar] [CrossRef] [PubMed]
- Ding, A.; Jeon, O.; Tang, R.; Lee, Y.B.; Lee, S.J.; Alsberg, E. Cell-Laden Multiple-Step and Reversible 4D Hydrogel Actuators to Mimic Dynamic Tissue Morphogenesis. Adv. Sci. 2021, 8, 2004616. [Google Scholar] [CrossRef] [PubMed]
- Yue, K.; Santiago, G.T.; Alvarez, M.M.; Tamayol, A.; Annabi, N.; Khademhosseini, A. Synthesis, Properties, and Biomedical Applications of Gelatin Methacryloyl (GelMA) Hydrogels. Biomaterials 2015, 73, 254–271. [Google Scholar] [CrossRef] [PubMed]
- Heltmann-Meyer, S.; Steiner, D.; Müller, C.; Schneidereit, D.; Friedrich, O.; Salehi, S.; Engel, F.B.; Arkudas, A.; Horch, R.E. Gelatin Methacryloyl Is a Slow Degrading Material Allowing Vascularization and Long-Term Use In Vivo. Biomed. Mater. 2021, 16, 065004. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bioink | Pressure (psi) | Acceleration (mm/s2) | Speed (mm/s) | Ink Temp (°C) |
|---|---|---|---|---|
| 14% GelMA | 16–14 | 100 | 9 | 24 |
| 14% GelMA, 2% OMA | 33–18 | 100 | 5 | 25 |
| 16% GelMA | 15–12 | 100 | 7 | 25 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martyniak, K.; Kennedy, S.; Karimzadeh, M.; Cruz, M.A.; Jeon, O.; Alsberg, E.; Kean, T.J. Optimizing Bioink Composition for Human Chondrocyte Expression of Lubricin. Bioengineering 2023, 10, 997. https://doi.org/10.3390/bioengineering10090997
Martyniak K, Kennedy S, Karimzadeh M, Cruz MA, Jeon O, Alsberg E, Kean TJ. Optimizing Bioink Composition for Human Chondrocyte Expression of Lubricin. Bioengineering. 2023; 10(9):997. https://doi.org/10.3390/bioengineering10090997
Chicago/Turabian StyleMartyniak, Kari, Sean Kennedy, Makan Karimzadeh, Maria A. Cruz, Oju Jeon, Eben Alsberg, and Thomas J. Kean. 2023. "Optimizing Bioink Composition for Human Chondrocyte Expression of Lubricin" Bioengineering 10, no. 9: 997. https://doi.org/10.3390/bioengineering10090997
APA StyleMartyniak, K., Kennedy, S., Karimzadeh, M., Cruz, M. A., Jeon, O., Alsberg, E., & Kean, T. J. (2023). Optimizing Bioink Composition for Human Chondrocyte Expression of Lubricin. Bioengineering, 10(9), 997. https://doi.org/10.3390/bioengineering10090997